Anti-Inflammatory Activity of a Peptide from Skipjack (Katsuwonus pelamis)

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Evaluation of the Anti-Inflammatory Activity of SEP in Transgenic Zebrafish
2.1.1. SEP Minimal Toxic Concentration (MTC)
2.1.2. SEP Anti-Inflammatory Activity Evaluation Results
2.2. Effects of SEP on Body Weight and Disease Activity Index (DAI) Score of Mice with UC
2.3. Effect of SEP on Colonic Tissue Structure in UC Mice
2.4. Effect of SEP on Serum IL-10, IL-6 and TNF-α Levels in Mice with UC
2.5. Analysis of the Intestinal Flora Structure in Mice
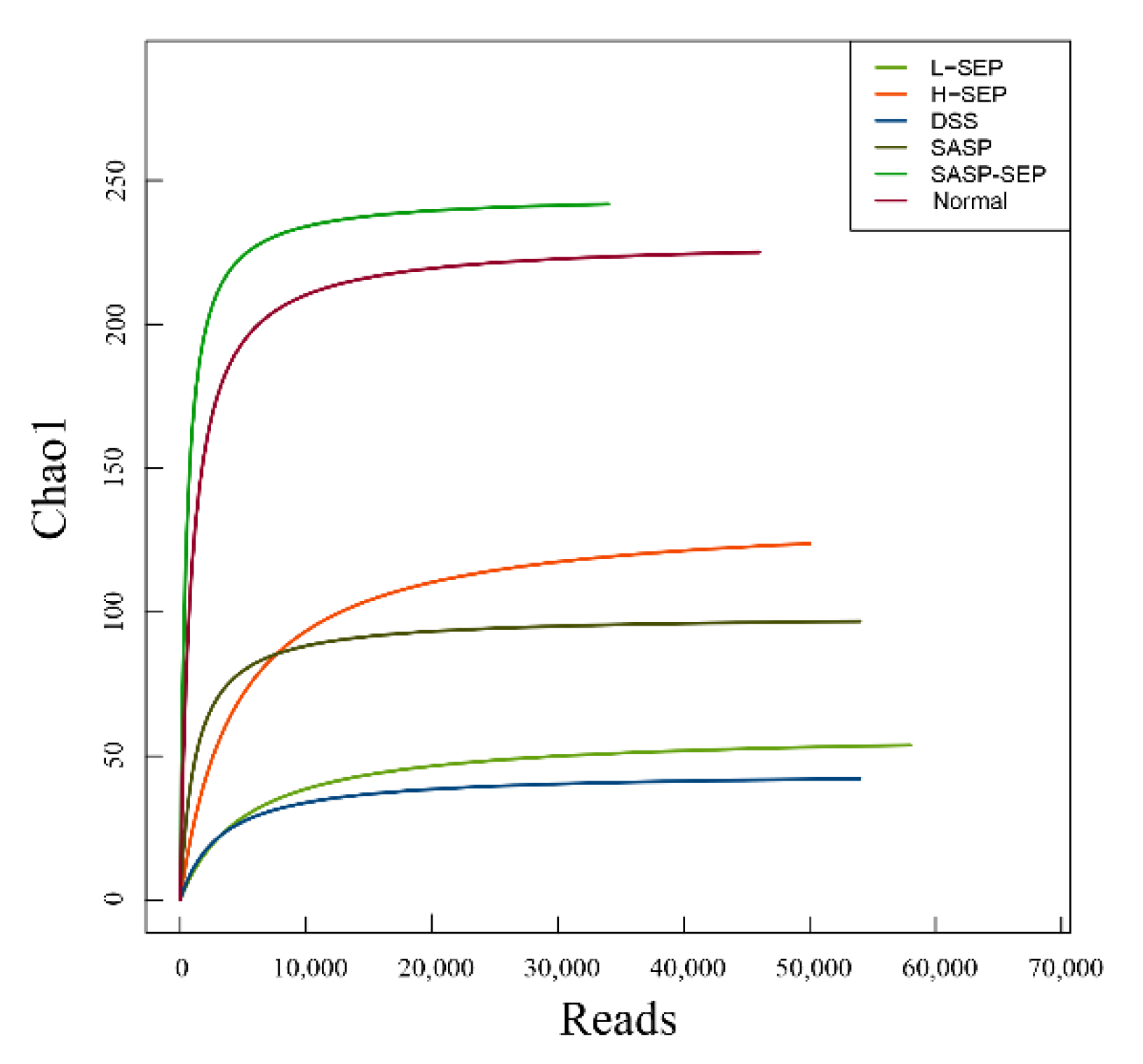
2.5.1. Diversity of the Intestinal Flora in Mice
2.5.2. Analysis of Species Abundance
3. Materials and Methods
3.1. Materials
3.1.1. Chemical
3.1.2. Experimental Animals
3.2. Equipment
3.3. Methods
3.3.1. Evaluation of the Anti-Inflammatory Activity of SEP in Transgenic Zebrafish
SEP Minimal Toxic Concentration (MTC)
Evaluation of the Anti-Inflammatory Activity of SEP
3.3.2. Evaluation of the Anti-UC Activity in Mice
3.3.3. Extraction and Analysis of DNA from the Gut Flora of Mice
3.4. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Osamu, T.; Shizuo, A. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar]
- Ala, A.; Dhillon, A.P.; Hodgson, H.J. Role of cell adhesion molecules in leukocyte recruitment in the liver and gut. Int. J. Exp. Pathol. 2010, 84, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Elzbieta, K.; Paul, K. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar]
- Feehan, K.T.; Gilroy, D.W. Is Resolution the End of Inflammation? Trends Mol. Med. 2019, 25, 198–214. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Zhu, L. Pay attention to the treatment of ulcerative colitis with Chinese and western medicine. Chin. J. Integr. Tradit. West. Med. Dig. 2016, 24, 571–574. [Google Scholar]
- Watanabe, T.; Ajioka, Y.; Mitsuyama, K.; Watanabe, K.; Hanai, H.; Nakase, H.; Kunisaki, R.; Matsuda, K.; Iwakiri, R.; Hida, N. Comparison of Targeted vs. Random Biopsies for Surveillance ofUlcerative Colitis-Associated Colorectal Cancer. Gastroenterology 2016, 151, 1122–1130. [Google Scholar] [CrossRef]
- Goyal, N.; Rana, A.; Ahlawat, A.; Bijjem, K.R.V.; Kumar, P. Animal models of inflammatory bowel disease: A review. Inflammopharmacology 2014, 22, 219–233. [Google Scholar] [CrossRef]
- Fang, S.; Evans, R.M. Wealth management in the gut. Nature 2013, 500, 538. [Google Scholar] [CrossRef]
- Wang, Q.; Tian, H.; Zhang, L. Animal Models of Ulcerative Colitis: A Review. Acta Neuropharmacol. 2013, 23–28. [Google Scholar]
- Yin, B.; Yu, M.; Wang, J.; Jiang, X.; Li, Z. Therapeutic effects of TNF-α binding peptide and TNFR blocking peptide on TNBS-induced colitis in rats. Immunol. J. 2007, 23, 663–667. [Google Scholar]
- Jia, Y.; Chen, Q. Effect of Bovine Casein Glycomacropeptide on MUC2 Expression in Mice with Ulcerative Colitis. Food Sci. 2010, 31, 365–368. [Google Scholar]
- Zhang, M.; Zhao, Y.; Yao, Y.; Xu, M.; Du, H.; Wu, N.; Tu, Y. Isolation and identification of peptides from simulated gastrointestinal digestion of preserved egg white and their anti-inflammatory activity in TNF-α-induced Caco-2 cells. J. Nutr. Biochem. 2019, 63, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Popov, C.S.F.C.; Magalhães, B.S.; Goodfellow, B.J.; Bocca, A.L.; Pereira, D.M.; Andrade, P.B.; Valentão, P.; Pereira, P.J.B.; Rodrigues, J.E.; de Holanda Veloso, P.H.; et al. Host-defense peptides AC12, DK16 and RC11 with immunomodulatory activity isolated from Hypsiboas raniceps skin secretion. Peptides 2019, 113, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Joshi, I.; Sudhakar, S.; Nazeer, R.A. Anti-inflammatory Properties of Bioactive Peptide Derived from Gastropod Influenced by Enzymatic Hydrolysis. Appl. Biochem. Biotechnol. 2016, 180, 1128–1140. [Google Scholar] [CrossRef]
- Stephani, J.; Radulovic, K.; Niess, J.H. Gut Microbiota, Probiotics and Inflammatory Bowel Disease. Arch. Immunol. Ther. Exp. 2011, 59, 161–177. [Google Scholar] [CrossRef]
- Yu, L.C.-H. Microbiota dysbiosis and barrier dysfunction in inflammatory bowel disease and colorectal cancers: Exploring a common ground hypothesis. J. Biomed. Sci. 2018, 25, 79. [Google Scholar] [CrossRef]
- Alexander, S.; Jutta, W.; Vera, L.B.; Hale, L.P.; Herbert, L. Spatial organization and composition of the mucosal flora in patients with inflammatory bowel disease. Z. Gastroenterol. 2005, 43, 3380–3389. [Google Scholar]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Ivanov, I.I.; Frutos, R.L.; Manel, N.; Yoshinaga, K.; Rifkin, D.B.; Sartor, R.B.; Finlay, B.B.; Littman, D.R. Specific microbiota direct the differentiation of IL-17-producing T-helper cells in the mucosa of the small intestine. Cell Host Microbe 2008, 4, 337–349. [Google Scholar] [CrossRef]
- Neff, C.P.; Rhodes, M.E.; Arnolds, K.L.; Collins, C.B.; Donnelly, J.; Nusbacher, N.; Jedlicka, P.; Schneider, J.M.; McCarter, M.D.; Shaffer, M.; et al. Diverse Intestinal Bacteria Contain Putative Zwitterionic Capsular Polysaccharides with Anti-inflammatory Properties. Cell Host Microbe 2016, 20, 535–547. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F. Safety Evaluation of Enterococci from Healthy Chinese Infants as Well as Their Cholesterol-Lowering Effect In Vitro and In Vivo D; Nanchang University: Nanchang, China, 2017. [Google Scholar]
- Yang, Z.; Duan, G.; Dai, F.; Chang, H. Research progress of enterococcus faecalis. Heilongjiang Anim. Sci. Vet. Med. 2017, 64–65. [Google Scholar]
- Chen, F.; Liu, S.; Wang, L.; Fan, X.; Sun, W. The high-throughput sequencing analysis of intestinal microbiota in rats administered antibiotics or ally. Chin. J. Microecol. 2015, 27, 1019–1021. [Google Scholar]
- La Reau, A.J.; Suen, G. The Ruminococci: Key symbionts of the gut ecosystem. J. Microbiol. 2018, 56, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Tuikhar, N.; Keisam, S.; Labala, R.K.; Ramakrishnan, P.; Arunkumar, M.C.; Ahmed, G.; Biagi, E.; Jeyaram, K. Comparative analysis of the gut microbiota in centenarians and young adults shows a common signature across genotypically non-related populations. Mech. Ageing Dev. 2019, 179, 23–35. [Google Scholar] [CrossRef]
- Urbanek, K.; De Angelis, A.; Spaziano, G.; Piegari, E.; Matteis, M.; Cappetta, D.; Esposito, G.; Russo, R.; Tartaglione, G.; De Palma, R.; et al. Intratracheal Administration of Mesenchymal Stem Cells Modulates Tachykinin System, Suppresses Airway Remodeling and Reduces Airway Hyperresponsiveness in an Animal Model. PLoS ONE 2016, 11, e0158746. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Yi, Z.; Diao, J.; Shao, M.; Zhao, L.; Cai, H.; Fan, Q.; Yao, X.; Sun, X. ShaoYao decoction ameliorates colitis-associated colorectal cancer by downregulating proinflammatory cytokines and promoting epithelial-mesenchymal transition. J. Transl. Med. 2014, 12, 105. [Google Scholar] [CrossRef]
- Dariushnejad, H.; Farajnia, S.; Zargami, N.; Khosroshahi, S.A.; Rahbarnia, L. Dsbc chaperone mediated soluble expression of human TNF-α in E. coli. Minerva Biotecnol. 2018, 30, 14–21. [Google Scholar]
- Zhang, Y. Study on the Relationship Between Vitamin D Signaling, Gut Microbiota and Risk for Colorectal Cancer and Its Mechanisms D; Second Military Medical University: Shanghai, China, 2016. [Google Scholar]
- Bao, W.; Wang, K.; Yang, Y.; Gong, K.; Zhong, M. Study on intestinal flora in patients with colorectal cancer. J. Colorectal Anal. Surg. 2009, 15, 224–226. [Google Scholar]
- Ge, F. Anti-Inflammatory Effect of Ethanol Extracts of Fagopyrumcymosum (Trey.) on Ulcerative Colitis Models; Nanjing University of Chinese Medicine: Nanjing, China, 2016. [Google Scholar]
- Zhen, Y. Application of HE staining technique in pathological diagnosis. Chin. Community Dr. 2016, 32, 109. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Experimental Concentration (μg/mL) | Death Number | Death Rate (%) | Toxicity Performance |
|---|---|---|---|---|
| Normal | - | 0 | 0 | - |
| Model | - | 0 | 0 | - |
| Solvent control (1% DMSO) | - | 0 | 0 | - |
| SEP | 31.25 | 0 | 0 | - |
| 62.5 | 0 | 0 | - | |
| 125 | 0 | 0 | - | |
| 250 | 0 | 0 | - | |
| 500 | 0 | 0 | - | |
| 1000 | - | - | precipitated | |
| 1500 | - | - | precipitated | |
| 2000 | - | - | precipitated |
| Group | Effective Sequence Number | Total Number of Bases (bp) | Average Length (bp) | OTUs |
|---|---|---|---|---|
| Low-dose SEP | 60,532 | 25,733,194 | 425 | 42 |
| High-dose SEP | 55,524 | 23,536,705 | 423 | 99 |
| DSS | 57,059 | 24,402,954 | 427 | 37 |
| SASP | 61,383 | 26,198,618 | 426 | 96 |
| SASP + SEP | 56,461 | 23,894,720 | 423 | 223 |
| Normal | 56,461 | 18,667,293 | 416 | 244 |
| Low-dose SEP | High-dose SEP | DSS | SASP | SASP + SEP | Normal | |
|---|---|---|---|---|---|---|
| Proteobacteria | 74.65 (%) | 70.52 (%) | 79.12 (%) | 64.89 (%) | 61.23 (%) | 7.74 (%) |
| Bacteroidetes | 18.31 (%) | 10.12 (%) | 0.3357 (%) | 13.96 (%) | 23.87 (%) | 63.88 (%) |
| Firmicutes | 6.28 (%) | 15.86 (%) | 20.54 (%) | 12.14 (%) | 11.78 (%) | 14.18 (%) |
| Groups | Number of Mice | Induction Drug | Gavage Drugs |
|---|---|---|---|
| Normal | 10 | Water | Saline |
| Model | 10 | 3% DSS | Saline |
| Low-dose SEP | 10 | 3% DSS | 1 g SEP /100 mL |
| High-dose SEP | 10 | 3% DSS | 3 g SEP /100 mL |
| SASP | 10 | 3% DSS | 2 g SASP /100 mL |
| SASP + SEP | 10 | 3% DSS | (2 g SASP + 3 g SEP)/100 mL |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.-g.; Ying, X.-g.; Gao, P.; Wang, C.-l.; Wang, Y.-f.; Yu, X.-w.; Chen, J.; Wang, B.; Luo, H.-y. Anti-Inflammatory Activity of a Peptide from Skipjack (Katsuwonus pelamis). Mar. Drugs 2019, 17, 582. https://doi.org/10.3390/md17100582
Wang Z-g, Ying X-g, Gao P, Wang C-l, Wang Y-f, Yu X-w, Chen J, Wang B, Luo H-y. Anti-Inflammatory Activity of a Peptide from Skipjack (Katsuwonus pelamis). Marine Drugs. 2019; 17(10):582. https://doi.org/10.3390/md17100582
Chicago/Turabian StyleWang, Zhi-gao, Xiao-guo Ying, Peng Gao, Chun-li Wang, Yi-fan Wang, Xin-wei Yu, Jing Chen, Bin Wang, and Hong-yu Luo. 2019. "Anti-Inflammatory Activity of a Peptide from Skipjack (Katsuwonus pelamis)" Marine Drugs 17, no. 10: 582. https://doi.org/10.3390/md17100582