Fucoidan–Fucoxanthin Ameliorated Cardiac Function via IRS1/GRB2/ SOS1, GSK3β/CREB Pathways and Metabolic Pathways in Senescent Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and LMWF/HSFUCO Administration
2.2. Grip Strength Test
2.3. Exhaustive Swimming Time Test
2.4. Reactive Oxygen Species and Aging
2.5. Measurements from Electrocardiogram (ECG)
2.6. Isolation of Cardiomyocytes
2.7. Urine Sample Preparation
2.8. Metabolomics Analysis
2.9. Action Potential Measurement in C57BL/6 Mice Myocytes by Patch-Clamp Technique
2.10. Histology (Hematoxylin and Eosin (H&E) Staining)
2.11. Masson Trichrome Staining
2.12. Western Blot Assay
2.13. Statistical Analysis
2.14. Bioinformatics Analysis
3. Results
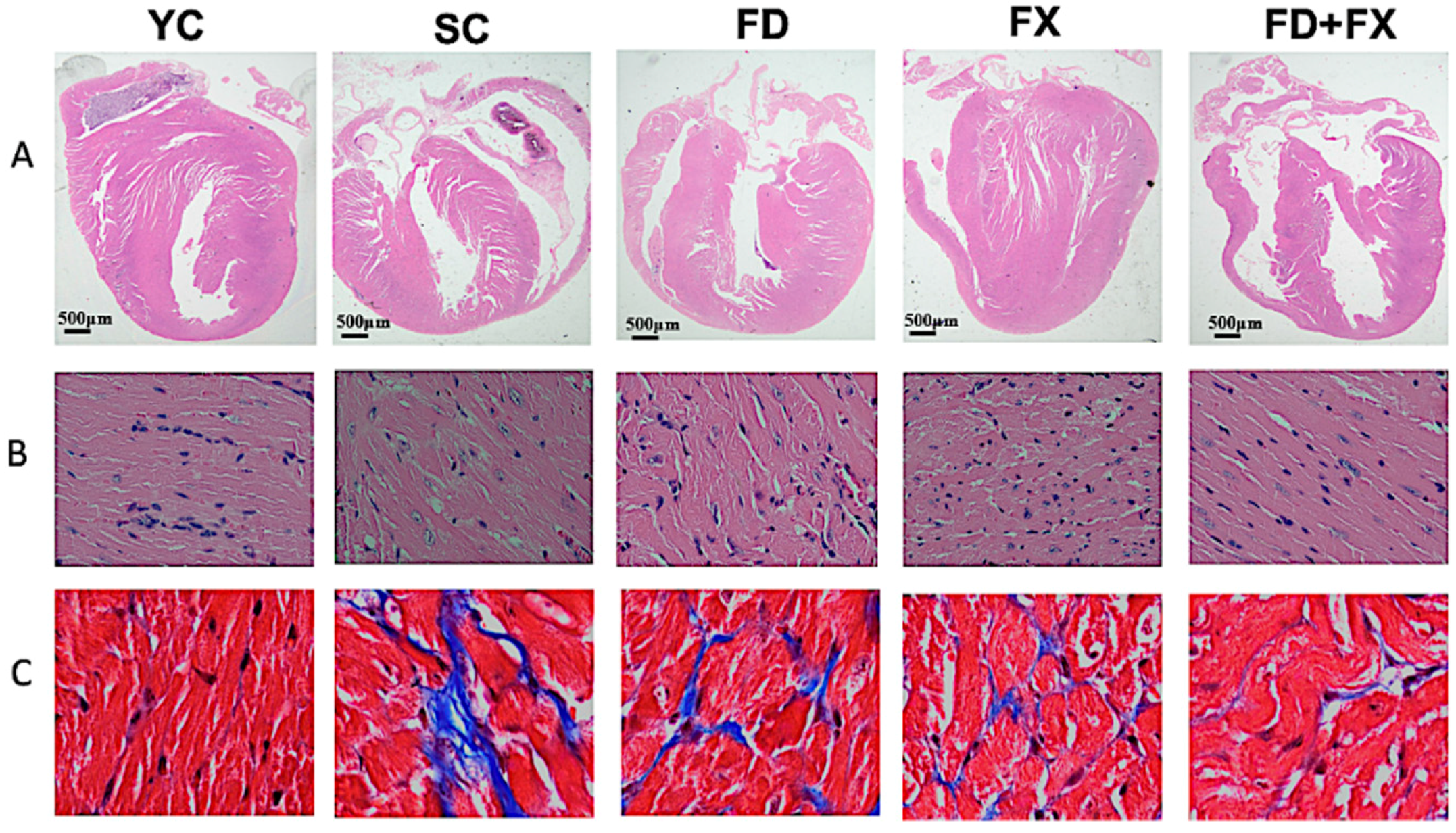
3.1. H&E Staining and Masson’s Trichrome Staining
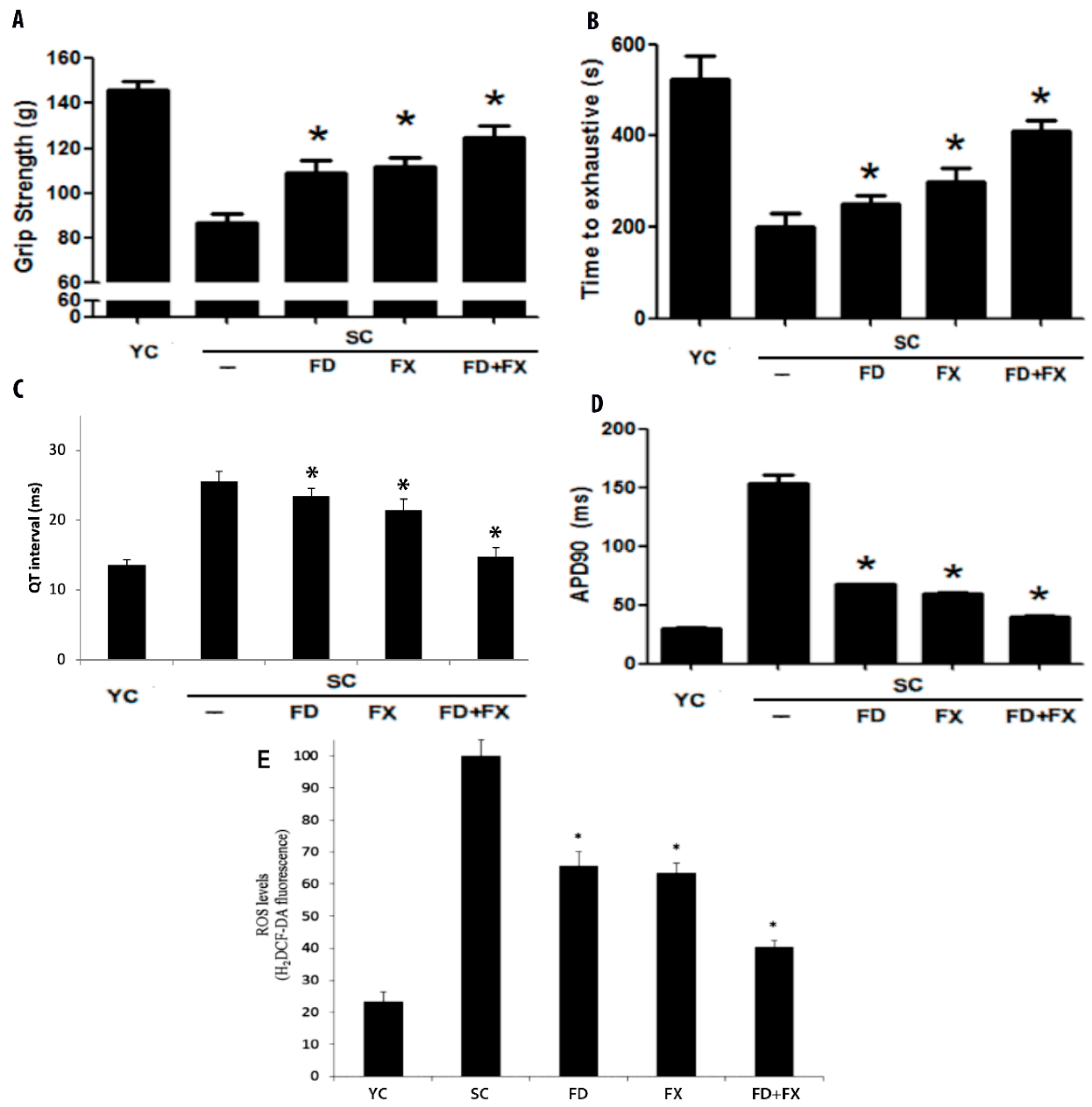
3.2. Grip Strength and Swimming Exhaustion Test Showed Significant Differences between Aging Mice and Young Controls
3.3. Effect of LMF and HSFUCO on QT Interval and Action Potential in Aging Mice Model
3.4. Reactive Oxygen Species and Aging
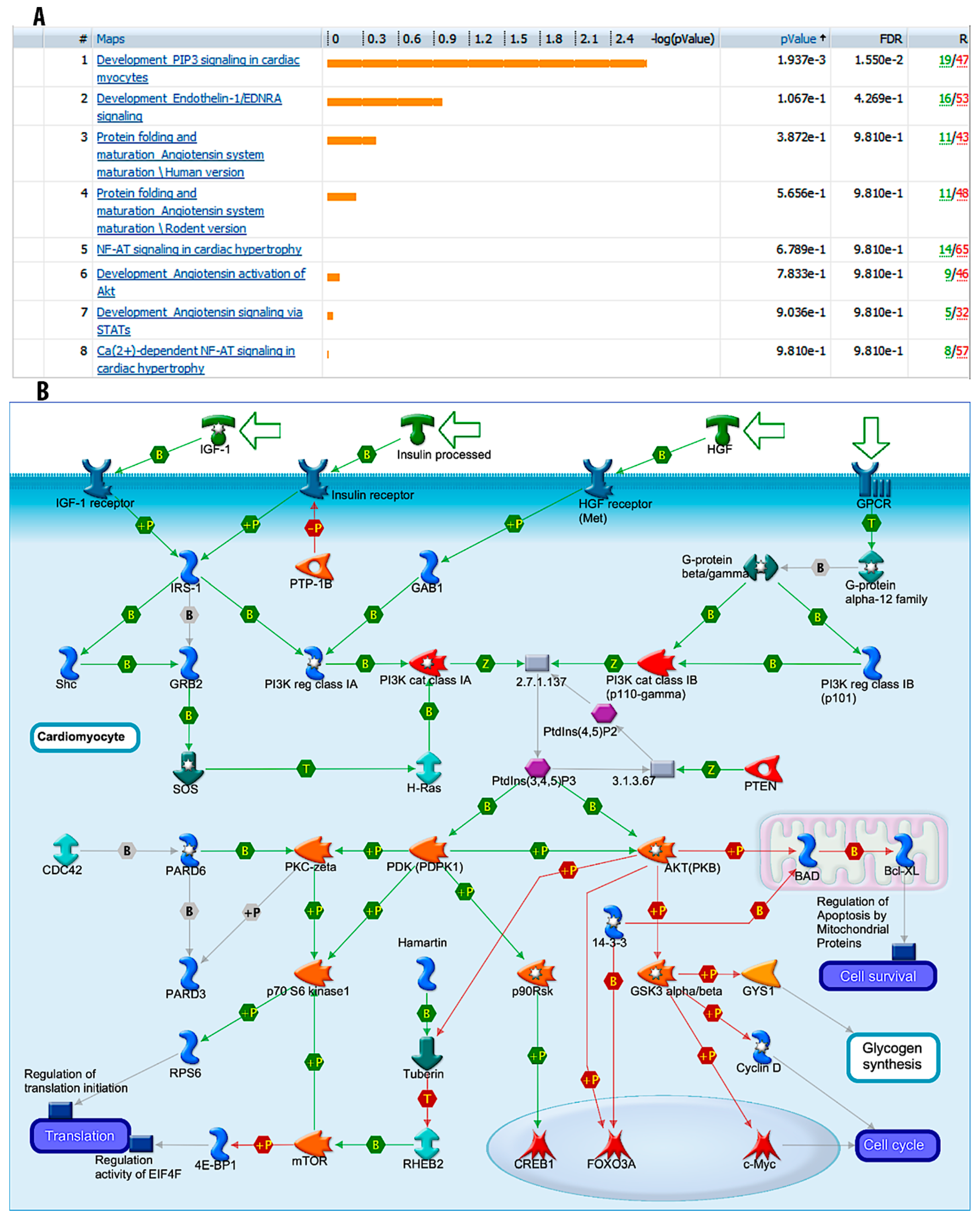
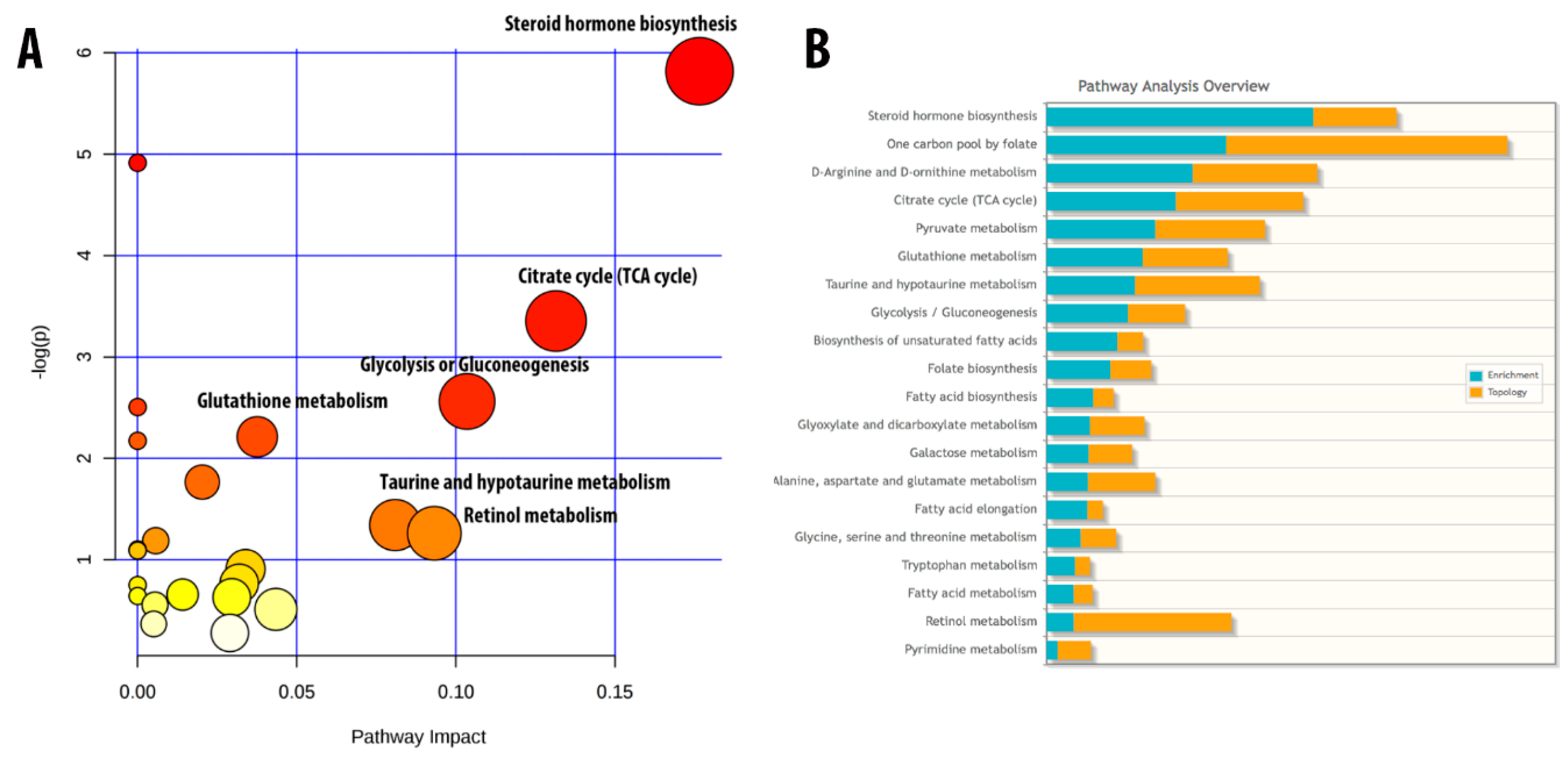
3.5. “Development_PIP3 Signaling in Cardiac Myocytes” Pathways are Significantly Modulated during Fucoidan Treatment
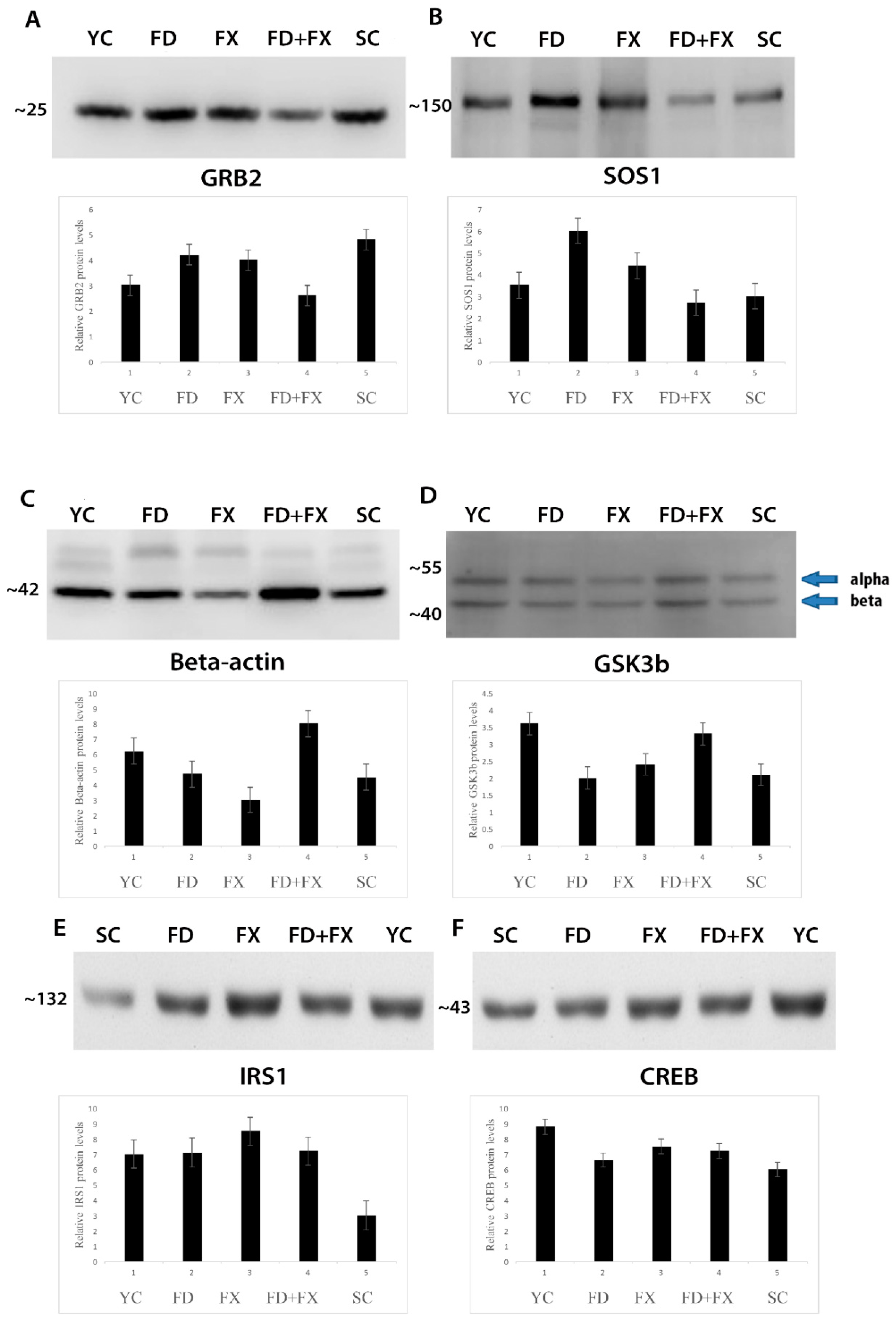
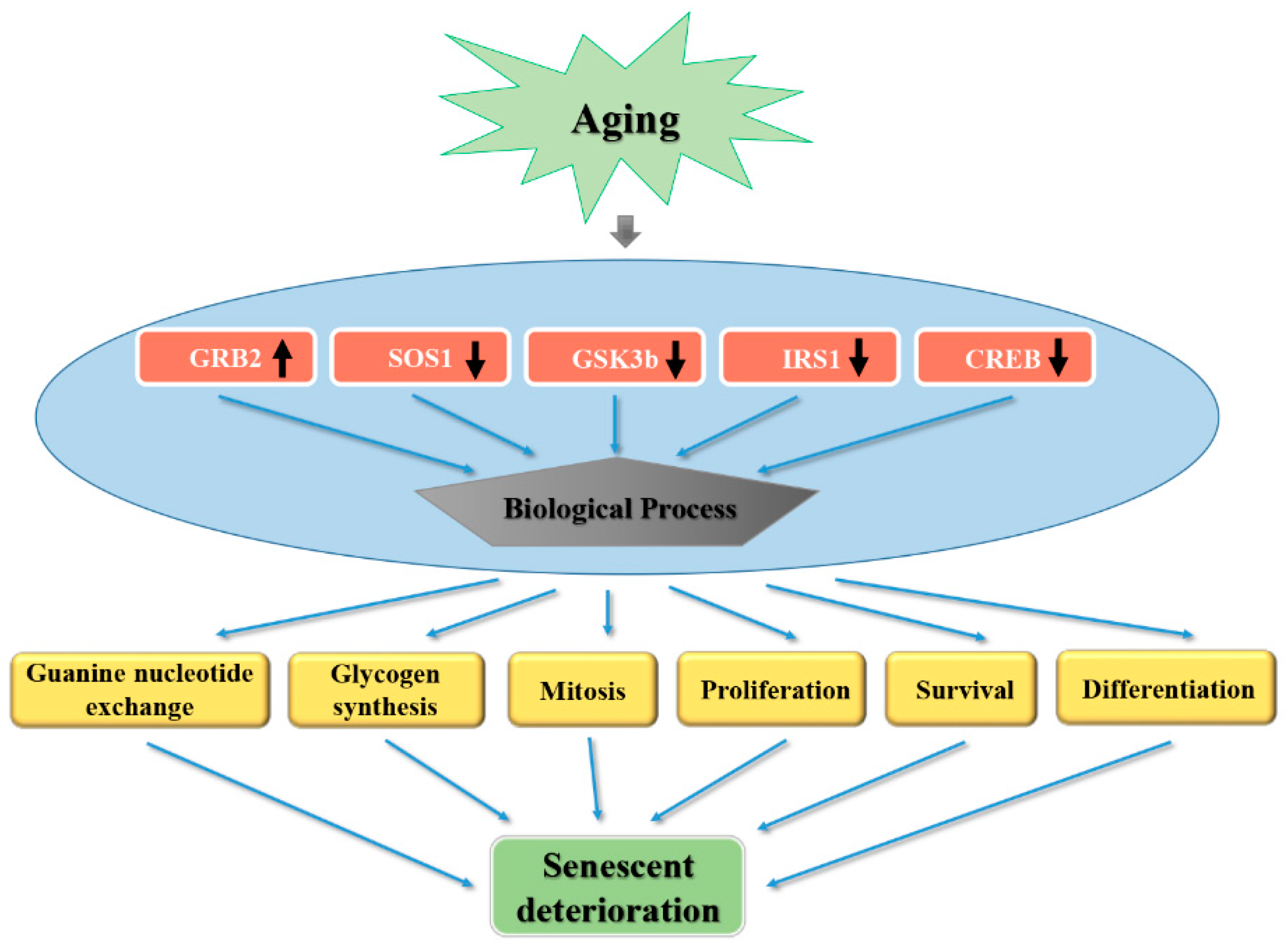
3.6. SOS1, GSK3β, GRB2, CREB, and IRS1 Protein Expression Level in Aging were Ameliorated with Fucoidan and Fucoxanthin
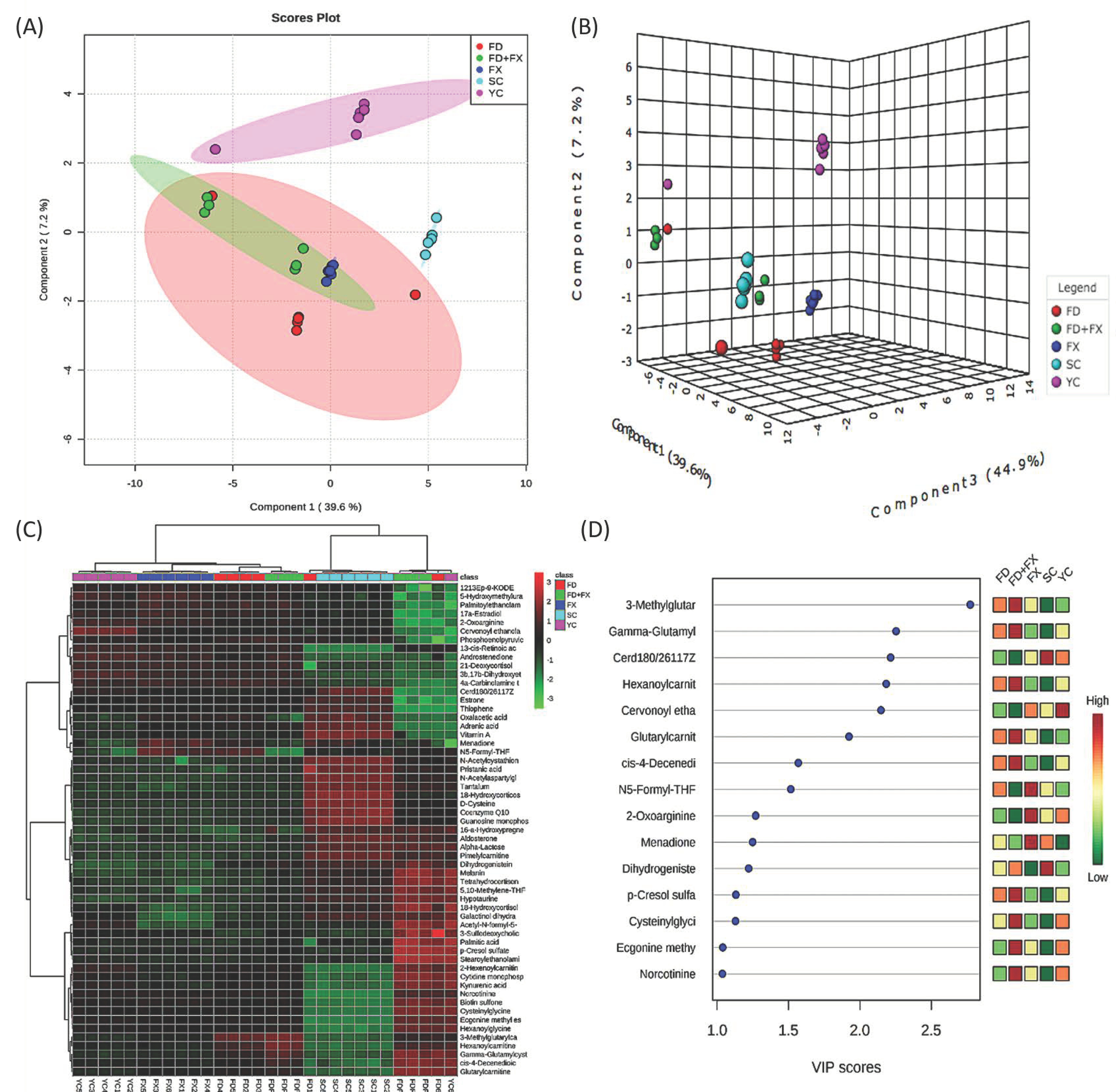
3.7. Chemometric Analysis and PLS-DA Showed Positive Effects for Fucoidan and Fucoxanthin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ames, B.N.; Shigenaga, M.K.; Hagen, T.M. Oxidants, antioxidants, and the degenerative diseases of aging. Proc. Natl. Acad. Sci. USA 1993, 90, 7915–7922. [Google Scholar] [CrossRef] [PubMed]
- Jo, H.E.; Randhawa, S.; Corte, T.J.; Moodley, Y. Idiopathic Pulmonary Fibrosis and the Elderly: Diagnosis and Management Considerations. Drugs Aging 2016, 33, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Borlaug, B.A. Cardiac aging and the fountain of youth. Eur. J. Heart Fail. 2016, 18, 611–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santulli, G.; Iaccarino, G. Adrenergic signaling in heart failure and cardiovascular aging. Maturitas 2016, 93, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.J.; Zhu, J.Z.; Bao, X.Y.; Zheng, Q.; Zheng, G.Q.; Wang, Y. Shexiang Baoxin Pills for Coronary Heart Disease in Animal Models: Preclinical Evidence and Promoting Angiogenesis Mechanism. Front. Pharmacol. 2017, 8, 404. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Liu, Y.; Shi, D.Z. Cardiovascular Actions and Therapeutic Potential of Tetramethylpyrazine (Active Component Isolated from Rhizoma Chuanxiong): Roles and Mechanisms. Biomed. Res. Int. 2016, 2016, 2430329. [Google Scholar] [CrossRef] [PubMed]
- Brenyo, A.; Aktas, M.K. Review of Complementary and Alternative Medical Treatment of Arrhythmias. Am. J. Cardiol. 2014, 113, 897–903. [Google Scholar] [CrossRef]
- Fitton, J.H. Therapies from fucoidan; multifunctional marine polymers. Mar. Drugs 2011, 9, 1731–1760. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.-Y.; Lin, T.-Y.; Hwang, P.-A.; Chen, R.-H.; Tsao, S.-M.; Hsu, J. Fucoidan induces changes in the epithelial-mesenchymal transition and decreases metastasis by enhancing ubiquitin-dependent TGFβ receptor degradation in breast cancer. Carcinogenesis 2012, 34, 874–884. [Google Scholar] [CrossRef] [PubMed]
- Tengdelius, M.E.; Lee, C.-J.; Grenegård, M.; Griffith, M.; Påhlsson, P.; Konradsson, P. Synthesis and Biological Evaluation of Fucoidan-Mimetic Glycopolymers through Cyanoxyl-Mediated Free-Radical Polymerization. Biomacromolecules 2014, 15, 2359–2368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, P.-A.; Chien, S.-Y.; Chan, Y.-L.; Lu, M.-K.; Wu, C.-H.; Kong, Z.-L.; Wu, C.-J. Inhibition of lipopolysaccharide (LPS)-induced inflammatory responses by Sargassum hemiphyllum sulfated polysaccharide extract in RAW 264.7 macrophage cells. J. Agric. Food Chem. 2011, 59, 2062–2068. [Google Scholar] [CrossRef]
- Irhimeh, M.R.; Fitton, J.H.; Lowenthal, R.M. Pilot clinical study to evaluate the anticoagulant activity of fucoidan. Blood Coagul. Fibrinolysis 2009, 20, 607–610. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Chung, D.; Shin, I.-S.; Lee, H.; Kim, J.; Lee, Y.; You, S. Effects of molecular weight and hydrolysis conditions on anticancer activity of fucoidans from sporophyll of Undaria pinnatifida. Int. J. Biol. Macromol. 2008, 43, 433–437. [Google Scholar] [CrossRef]
- Soeda, S.; Sakaguchi, S.; Shimeno, H.; Nagamatsu, A. Fibrinolytic and anticoagulant activities of highly sulfated fucoidan. Biochem. Pharm. 1992, 43, 1853–1858. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Q.; Zhang, Z.; Song, H.; Li, P. Potential antioxidant and anticoagulant capacity of low molecular weight fucoidan fractions extracted from Laminaria japonica. Int. J. Biol. Macromol. 2010, 46, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Q.; Zhang, Z.; Li, Z. Antioxidant activity of sulfated polysaccharide fractions extracted from Laminaria japonica. Int. J. Biol. Macromol. 2008, 42, 127–132. [Google Scholar] [CrossRef] [PubMed]
- De Souza, M.C.R.; Marques, C.T.; Dore, C.M.G.; da Silva, F.R.F.; Rocha, H.A.O.; Leite, E.L. Antioxidant activities of sulfated polysaccharides from brown and red seaweeds. J. Appl. Phycol. 2007, 19, 153–160. [Google Scholar] [CrossRef]
- Park, H.Y.; Han, M.H.; Park, C.; Jin, C.-Y.; Kim, G.-Y.; Choi, I.-W.; Kim, N.D.; Nam, T.-J.; Kwon, T.K.; Choi, Y.H. Anti-inflammatory effects of fucoidan through inhibition of NF-κB, MAPK and Akt activation in lipopolysaccharide-induced BV2 microglia cells. Food Chem. Toxicol. 2011, 49, 1745–1752. [Google Scholar] [CrossRef]
- Medeiros, V.; Queiroz, K.; Cardoso, M.; Monteiro, G.; Oliveira, F.; Chavante, S.; Guimaraes, L.; Rocha, H.; Leite, E. Sulfated galactofucan from Lobophora variegata: Anticoagulant and anti-inflammatory properties. Biochemistry (Moscow) 2008, 73, 1018–1024. [Google Scholar] [CrossRef]
- Colliec, S.; Boisson-vidal, C.; Jozefonvicz, J. A low molecular weight fucoidan fraction from the brown seaweed Pelvetia canaliculata. Phytochemistry 1994, 35, 697–700. [Google Scholar] [CrossRef]
- Luyt, C.-E.; Meddahi-Pellé, A.; Ho-Tin-Noe, B.; Colliec-Jouault, S.; Guezennec, J.; Louedec, L.; Prats, H.; Jacob, M.-P.; Osborne-Pellegrin, M.; Letourneur, D. Low-molecular-weight fucoidan promotes therapeutic revascularization in a rat model of critical hindlimb ischemia. J. Pharm. Exp. Ther. 2003, 305, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Deux, J.-F.; Meddahi-Pellé, A.; Le Blanche, A.F.; Feldman, L.J.; Colliec-Jouault, S.; Brée, F.; Boudghène, F.; Michel, J.-B.; Letourneur, D. Low molecular weight fucoidan prevents neointimal hyperplasia in rabbit iliac artery in-stent restenosis model. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1604–1609. [Google Scholar] [CrossRef] [PubMed]
- Lake, A.C.; Vassy, R.; Di Benedetto, M.; Lavigne, D.; Le Visage, C.; Perret, G.Y.; Letourneur, D. Low molecular weight fucoidan increases VEGF165-induced endothelial cell migration by enhancing VEGF165 binding to VEGFR-2 and NRP1. J. Biol. Chem. 2006, 281, 37844–37852. [Google Scholar] [CrossRef] [PubMed]
- Thomes, P.; Rajendran, M.; Pasanban, B.; Rengasamy, R. Cardioprotective activity of Cladosiphon okamuranus fucoidan against isoproterenol induced myocardial infarction in rats. Phytomedicine 2010, 18, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Gao, Y.; Xing, Y.; Zhu, H.; Shen, J.; Tian, J. Fucoidan, a sulfated polysaccharide from brown algae, against myocardial ischemia–reperfusion injury in rats via regulating the inflammation response. Food Chem. Toxicol. 2011, 49, 2090–2095. [Google Scholar] [CrossRef] [PubMed]
- Park, S.B.; Chun, K.R.; Kim, J.K.; Suk, K.; Jung, Y.M.; Lee, W.H. The differential effect of high and low molecular weight fucoidans on the severity of collagen-induced arthritis in mice. Phytother. Res. 2010, 24, 1384–1391. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.J.; Yoon, W.J.; Kim, K.N.; Ahn, G.N.; Kang, S.M.; Kang, D.H.; Affan, A.; Oh, C.; Jung, W.K.; Jeon, Y.J. Evaluation of anti-inflammatory effect of fucoxanthin isolated from brown algae in lipopolysaccharide-stimulated RAW 264.7 macrophages. Food Chem. Toxicol. 2010, 48, 2045–2451. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.P.; Hou, Y.H. First evidence for the anti-inflammatory activity of fucoxanthin in high-fat-diet-induced obesity in mice and the antioxidant functions in PC12 cells. Inflammation 2014, 37, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Ha, S. Methods of Treatment of Cardiovascular and Cerebrovascular Diseases with Fucoidan. U.S. Patent Application No. 12/397,443, 2 July 2009. [Google Scholar]
- Ghosh, J.; Miller, R.A. Rapid tyrosine phosphorylation of Grb2 and Shc in T cells exposed to anti-CD3, anti-CD4, and anti-CD45 stimuli: Differential effects of aging. Mech. Ageing Dev. 1995, 80, 171–187. [Google Scholar] [CrossRef]
- Chien, T.-Y.; Yang, C.-M. Human Cardiac Hypertrophy Induced by Thrombin/COX-2 is Mediated Through a c-Src/Pyk2/EGFR/CREB-dependent Pathway. FASEB J. 2015, 29 (Suppl. S1), 781. [Google Scholar]
- Chien, P.T.-Y.; Lin, C.-C.; Hsiao, L.-D.; Yang, C.-M. c-Src/Pyk2/EGFR/PI3K/Akt/CREB-activated pathway contributes to human cardiomyocyte hypertrophy: Role of COX-2 induction. Mol. Cell. Endocrinol. 2015, 409, 59–72. [Google Scholar] [CrossRef]
- Hutter, D.; Yo, Y.; Chen, W.; Liu, P.; Holbrook, N.J.; Roth, G.S.; Liu, Y. Age-related decline in Ras/ERK mitogen-activated protein kinase cascade is linked to a reduced association between Shc and EGF receptor. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2000, 55, B125–B134. [Google Scholar] [CrossRef]
- Sadoshima, J.-I.; Izumo, S. The heterotrimeric G q protein-coupled angiotensin II receptor activates p21 ras via the tyrosine kinase-Shc-Grb2-Sos pathway in cardiac myocytes. EMBO J. 1996, 15, 775. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Tu, X.; Prisco, M.; Wu, A.; Casiburi, I.; Baserga, R. Insulin-like growth factor I receptor signaling and nuclear translocation of insulin receptor substrates 1 and 2. Mol. Endocrinol. 2003, 17, 472–486. [Google Scholar] [CrossRef] [PubMed]
- Selman, C.; Lingard, S.; Choudhury, A.I.; Batterham, R.L.; Claret, M.; Clements, M.; Ramadani, F.; Okkenhaug, K.; Schuster, E.; Blanc, E. Evidence for lifespan extension and delayed age-related biomarkers in insulin receptor substrate 1 null mice. FASEB J. 2008, 22, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.-E.; Huang, W.-C.; Liao, C.-C.; Chang, Y.-K.; Kan, N.-W.; Huang, C.-C. Resveratrol protects against physical fatigue and improves exercise performance in mice. Molecules 2013, 18, 4689–4702. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Huang, A.; Zhang, Q.; Lin, Y.-C.; Yu, H.-G. Novel mechanism for suppression of hyperpolarization-activated cyclic nucleotide-gated pacemaker channels by receptor-like tyrosine phosphatase-α. J. Biol. Chem. 2008, 283, 29912–29919. [Google Scholar] [CrossRef]
- Phan, N.N.; Wang, C.-Y.; Lin, Y.-C. The novel regulations of MEF2A, CAMKK2, CALM3, and TNNI3 in ventricular hypertrophy induced by arsenic exposure in rats. Toxicology 2014, 324, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Yokota, T.; Nomura, K.; Nagashima, M.; Kamimura, N. Fucoidan alleviates high-fat diet-induced dyslipidemia and atherosclerosis in ApoE shl mice deficient in apolipoprotein E expression. J. Nutr. Biochem. 2016, 32, 46–54. [Google Scholar] [CrossRef]
- Ito, T.; Asakura, K.; Tougou, K.; Fukuda, T.; Kubota, R.; Nonen, S.; Fujio, Y.; Azuma, J. Regulation of cytochrome P450 2E1 under hypertonic environment through TonEBP in human hepatocytes. Mol. Pharm. 2007, 72, 173–181. [Google Scholar] [CrossRef]
- Ohnishi, K.; Yoshida, H.; Shigeno, K.; Nakamura, S.; Fujisawa, S.; Naito, K.; Shinjo, K.; Fujita, Y.; Matsui, H.; Takeshita, A. Prolongation of the QT interval and ventricular tachycardia in patients treated with arsenic trioxide for acute promyelocytic leukemia. Ann. Intern. Med. 2000, 133, 881–885. [Google Scholar] [CrossRef] [PubMed]
- Lamm, S.H.; Engel, A.; Penn, C.A.; Chen, R.; Feinleib, M. Arsenic cancer risk confounder in southwest Taiwan data set. Environ. Health Perspect. 2006, 114, 1077–1082. [Google Scholar] [CrossRef] [PubMed]
- Roden, D.M. Keep the QT interval: It is a reliable predictor of ventricular arrhythmias. Heart Rhythm 2008, 5, 1213–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.J. Cardiac hypertrophy: A risk factor for QT-prolongation and cardiac sudden death. Toxicol. Pathol. 2006, 34, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Chuda, Y.; Suzuki, M.; Nagata, T. Fucoxanthin as the major antioxidant in Hijikia fusiformis, a common edible seaweed. Biosci. Biotechnol. Biochem. 1999, 63, 605–607. [Google Scholar] [CrossRef]
- Fung, A.; Hamid, N.; Lu, J. Fucoxanthin content and antioxidant properties of Undaria pinnatifida. Food Chem. 2013, 136, 1055–1062. [Google Scholar] [CrossRef]
- Nomura, T.; Kikuchi, M.; Kubodera, A.; Kawakami, Y. Proton-donative antioxidant activity of fucoxanthin with 1,1-diphenyl-2-picrylhydrazyl (DPPH). IUBMB Life 1997, 42, 361–370. [Google Scholar] [CrossRef]
- Maeda, H. Nutraceutical effects of fucoxanthin for obesity and diabetes therapy: A review. J. Oleo Sci. 2015, 64, 125–132. [Google Scholar] [CrossRef]
- Bejma, J.; Ji, L.L. Aging and acute exercise enhance free radical generation in rat skeletal muscle. J. Appl. Physiol. 1999, 87, 465–470. [Google Scholar] [CrossRef]
- Petropoulos, I.; Friguet, B. Protein maintenance in aging and replicative senescence: A role for the peptide methionine sulfoxide reductases. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2005, 1703, 261–266. [Google Scholar] [CrossRef]
- Stadtman, E.R.; Van Remmen, H.; Richardson, A.; Wehr, N.B.; Levine, R.L. Methionine oxidation and aging. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2005, 1703, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Voss, P.; Siems, W. Clinical oxidation parameters of aging. Free Radic. Res. 2006, 40, 1339–1349. [Google Scholar] [CrossRef] [PubMed]
- Kusaykin, M.; Bakunina, I.; Sova, V.; Ermakova, S.; Kuznetsova, T.; Besednova, N.; Zaporozhets, T.; Zvyagintseva, T. Structure, biological activity, and enzymatic transformation of fucoidans from the brown seaweeds. Biotechnol. J. 2008, 3, 904–915. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, N.; Kubota, T.; Kawano, S.; Monden, Y.; Feldman, A.M.; Tsutsui, H.; Takeshita, A.; Sunagawa, K. Blockade of NF-κB improves cardiac function and survival without affecting inflammation in TNF-α-induced cardiomyopathy. Cardiovasc. Res. 2005, 66, 520–529. [Google Scholar] [CrossRef] [Green Version]
- Kawano, S.; Kubota, T.; Monden, Y.; Tsutsumi, T.; Inoue, T.; Kawamura, N.; Tsutsui, H.; Sunagawa, K. Blockade of NF-κB improves cardiac function and survival after myocardial infarction. Am. J. Physiol.-Heart Circ. Physiol. 2006, 291, H1337–H1344. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Yoshiyama, M.; Izumi, Y.; Kawano, H.; Kimoto, M.; Zhan, Y.; Iwao, H. Effects of combination of ACE inhibitor and angiotensin receptor blocker on cardiac remodeling, cardiac function, and survival in rat heart failure. Circulation 2001, 103, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Fujita, A.; Takahashi-Yanaga, F.; Morimoto, S.; Yoshihara, T.; Arioka, M.; Igawa, K.; Tomooka, K.; Hoka, S.; Sasaguri, T. 2,5-Dimethylcelecoxib prevents pressure-induced left ventricular remodeling through GSK-3 activation. Hypertens. Res. 2017, 40, 130. [Google Scholar] [CrossRef]
- Zhang, S.; Weinheimer, C.; Courtois, M.; Kovacs, A.; Zhang, C.E.; Cheng, A.M.; Wang, Y.; Muslin, A.J. The role of the Grb2–p38 MAPK signaling pathway in cardiac hypertrophy and fibrosis. J. Clin. Investig. 2003, 111, 833–841. [Google Scholar] [CrossRef] [Green Version]
- Cai, W.; He, J.C.; Zhu, L.; Lu, C.; Vlassara, H. Advanced glycation end product (AGE) receptor 1 suppresses cell oxidant stress and activation signaling via EGF receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 13801–13806. [Google Scholar] [CrossRef] [Green Version]
- Akram, M. Citric acid cycle and role of its intermediates in metabolism. Cell Biochem. Biophys. 2014, 68, 475–478. [Google Scholar] [CrossRef]
- Gwynne, J.T.; Strauss, J.F., III. The role of lipoproteins in steroidogenesis and cholesterol metabolism in steroidogenic glands. Endocr. Rev. 1982, 3, 299–329. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.K.; Shen, W.-J.; Azhar, S. Impact of aging on steroid hormone biosynthesis and secretion. Open Longev. Sci 2012, 6, 1–30. [Google Scholar] [CrossRef]
- James, E.L.; Michalek, R.D.; Pitiyage, G.N.; de Castro, A.M.; Vignola, K.S.; Jones, J.; Mohney, R.P.; Karoly, E.D.; Prime, S.S.; Parkinson, E.K. Senescent human fibroblasts show increased glycolysis and redox homeostasis with extracellular metabolomes that overlap with those of irreparable DNA damage, aging, and disease. J. Proteome Res. 2015, 14, 1854–1871. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, P.-M.; Li, K.-L.; Lin, Y.-C. Fucoidan–Fucoxanthin Ameliorated Cardiac Function via IRS1/GRB2/ SOS1, GSK3β/CREB Pathways and Metabolic Pathways in Senescent Mice. Mar. Drugs 2019, 17, 69. https://doi.org/10.3390/md17010069
Chang P-M, Li K-L, Lin Y-C. Fucoidan–Fucoxanthin Ameliorated Cardiac Function via IRS1/GRB2/ SOS1, GSK3β/CREB Pathways and Metabolic Pathways in Senescent Mice. Marine Drugs. 2019; 17(1):69. https://doi.org/10.3390/md17010069
Chicago/Turabian StyleChang, Po-Ming, Kuan-Lun Li, and Yen-Chang Lin. 2019. "Fucoidan–Fucoxanthin Ameliorated Cardiac Function via IRS1/GRB2/ SOS1, GSK3β/CREB Pathways and Metabolic Pathways in Senescent Mice" Marine Drugs 17, no. 1: 69. https://doi.org/10.3390/md17010069