Biogenic Polyphosphate Nanoparticles from a Marine Cyanobacterium Synechococcus sp. PCC 7002: Production, Characterization, and Anti-Inflammatory Properties In Vitro

 and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
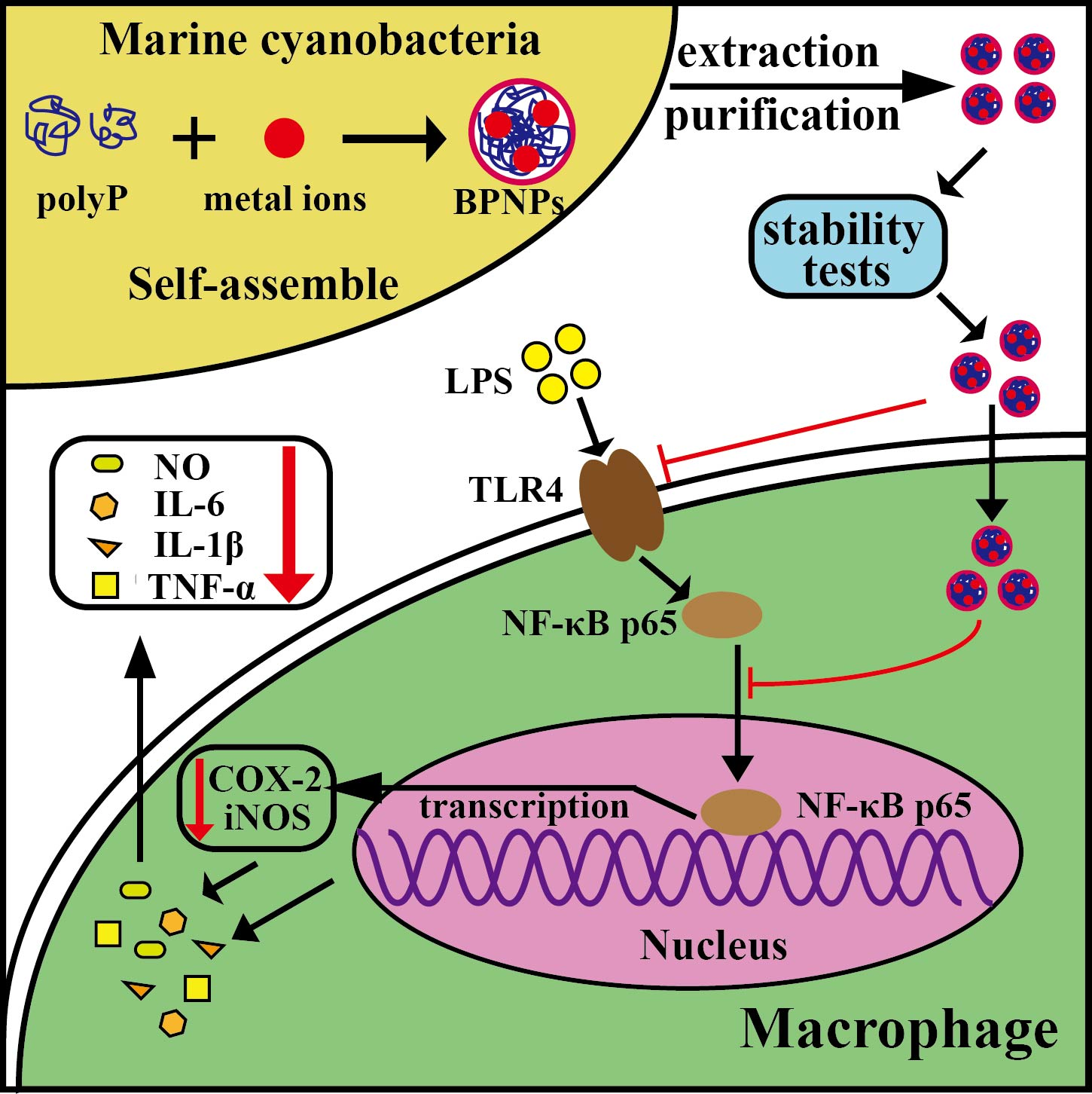
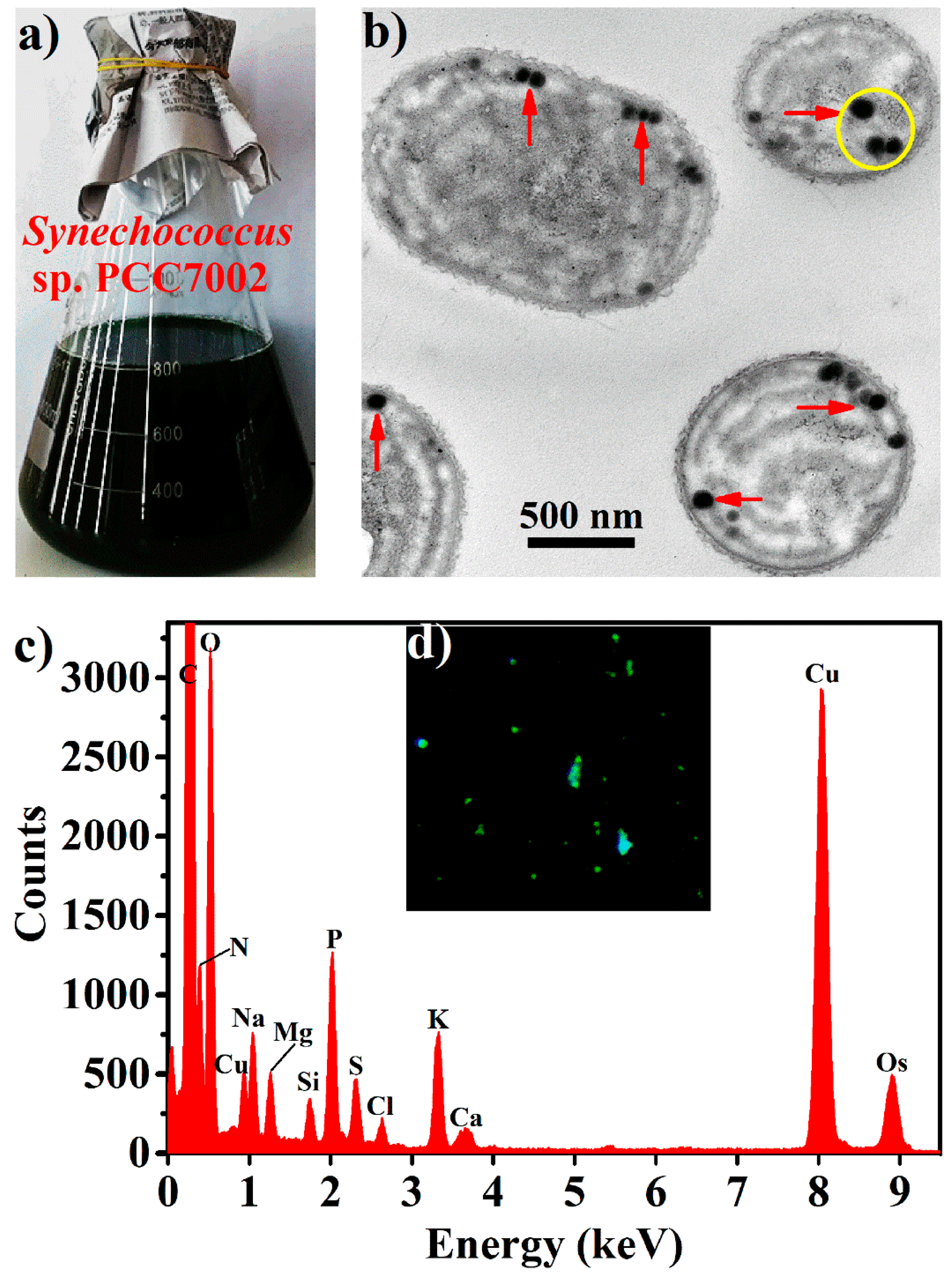
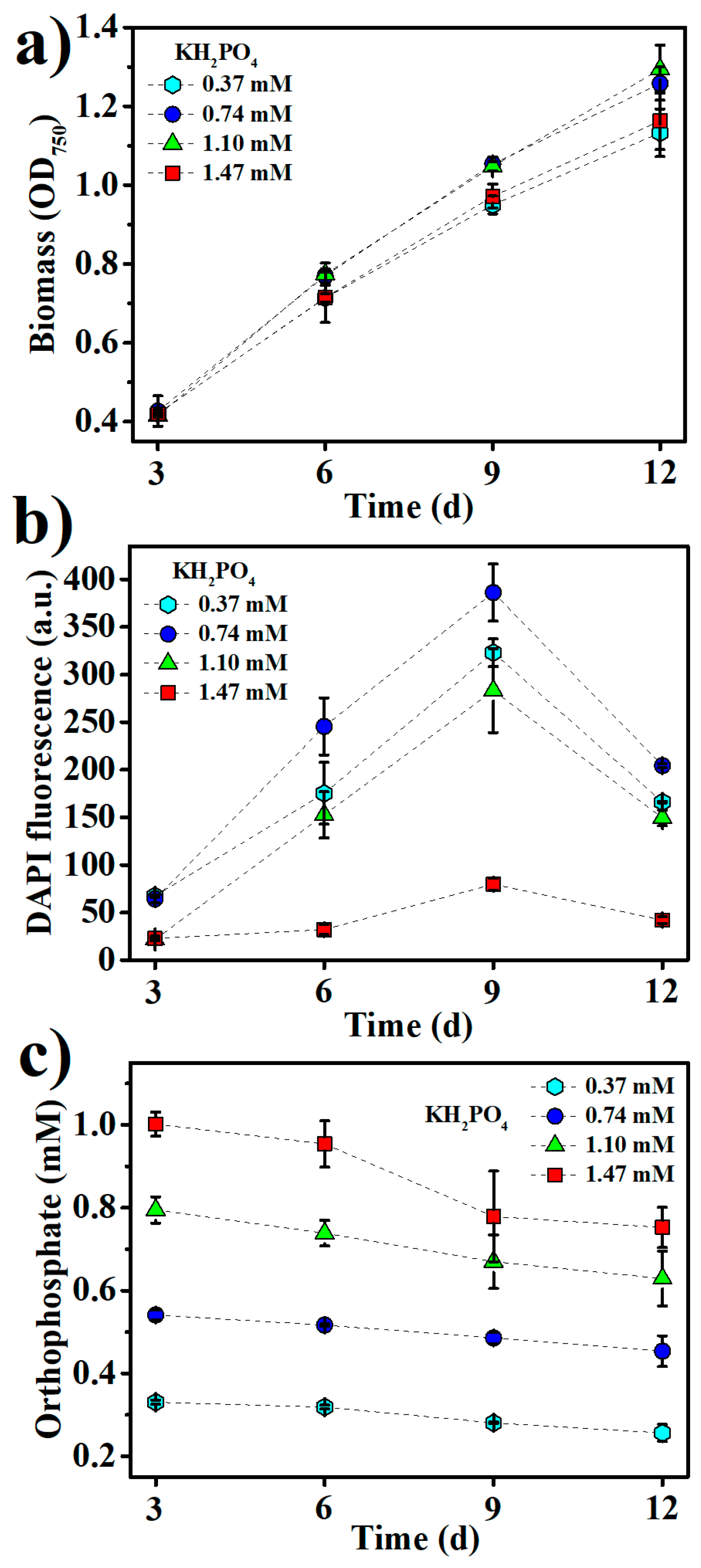
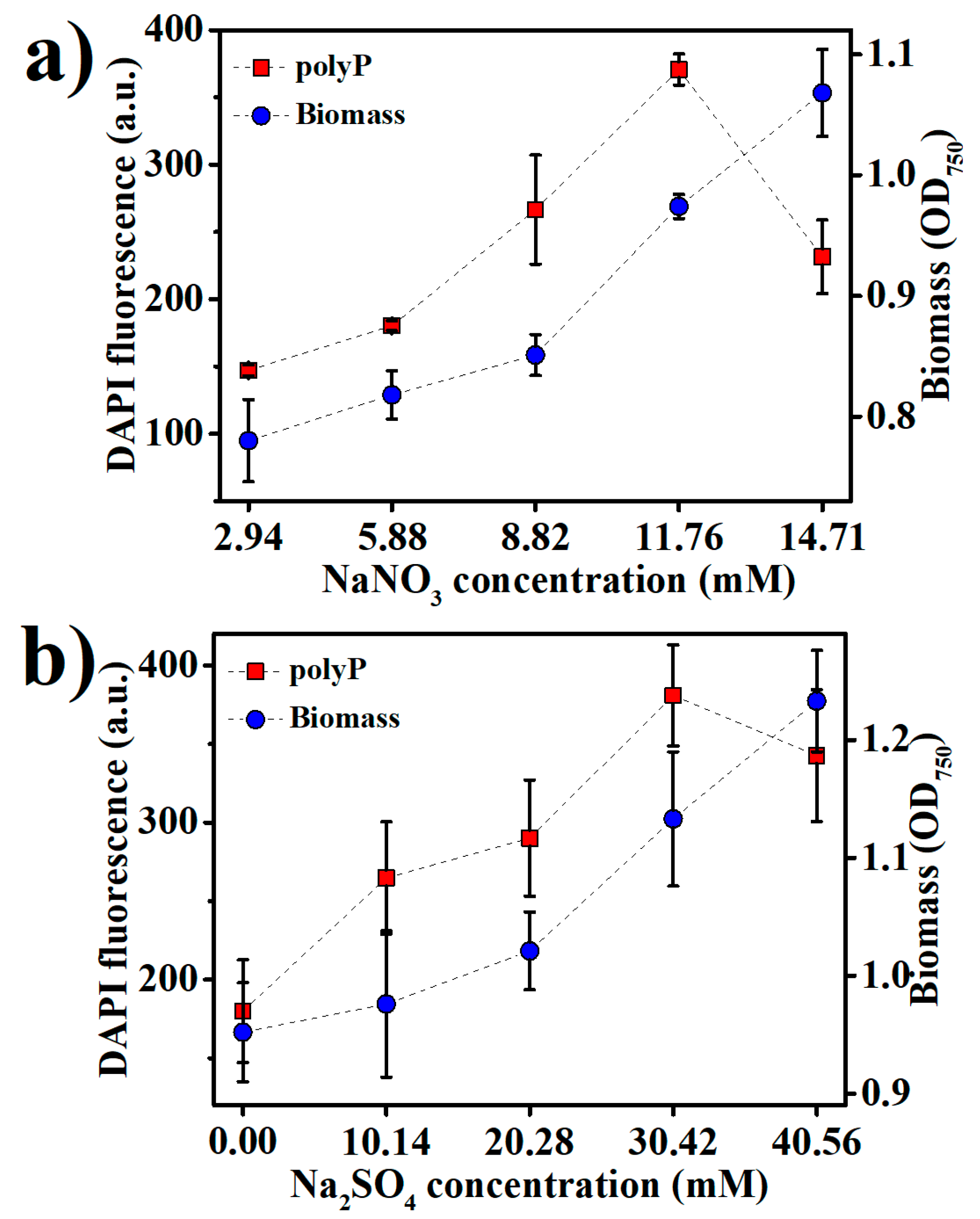
2.1. Production of BPNPs
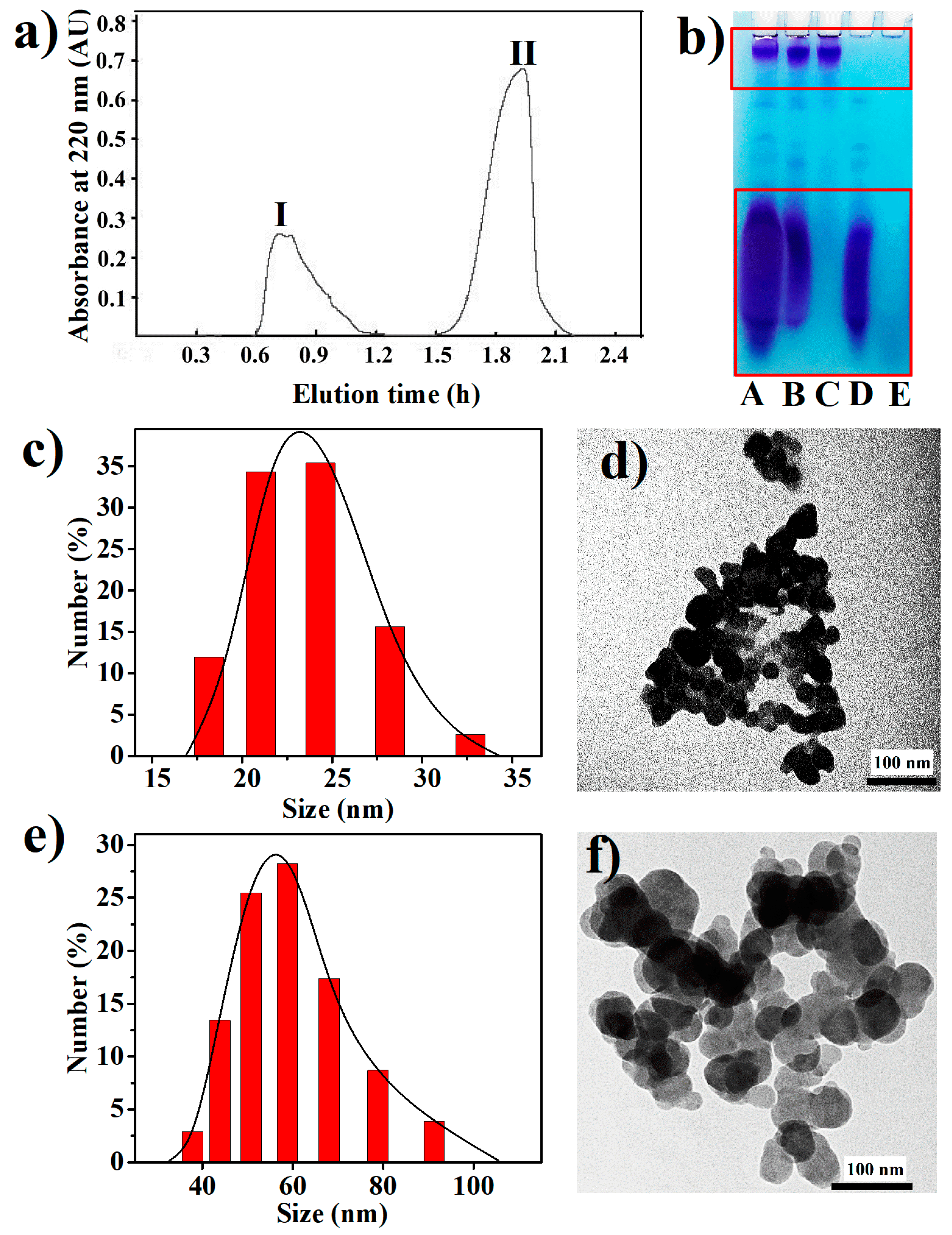
2.2. Purification and Characterization of BPNPs
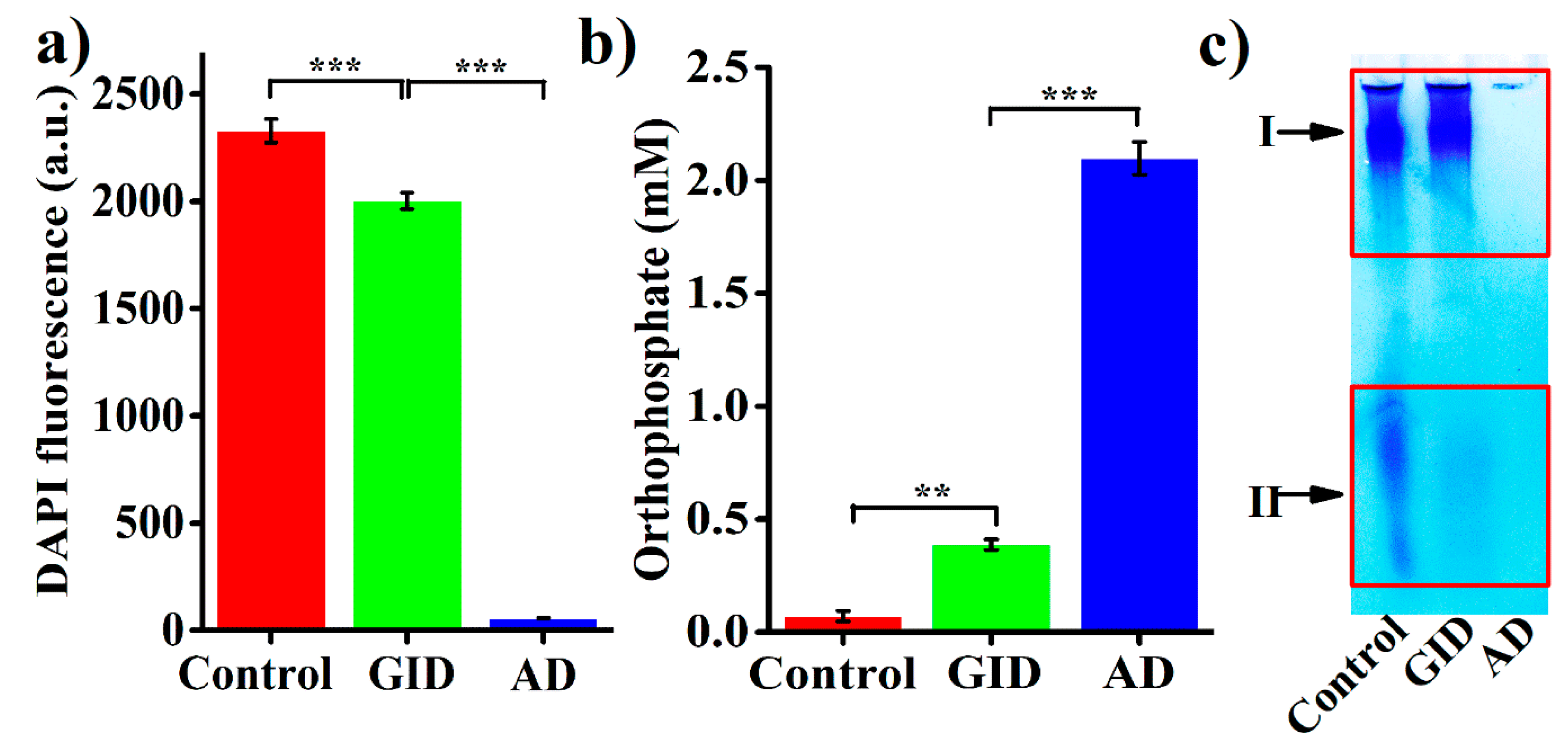
2.3. Stability of BPNPs in Simulated Gastrointestinal Digestion (GID)
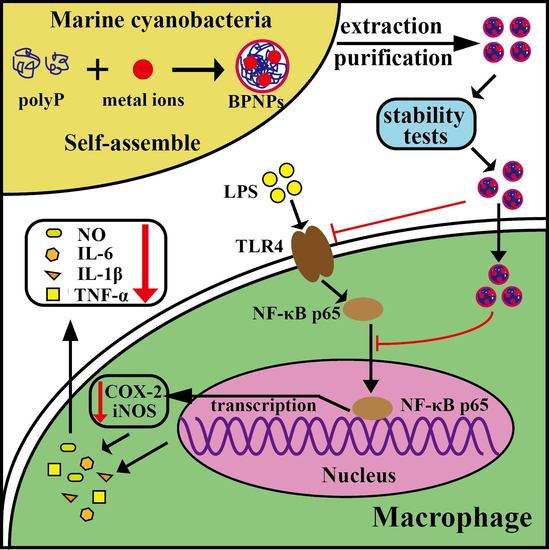
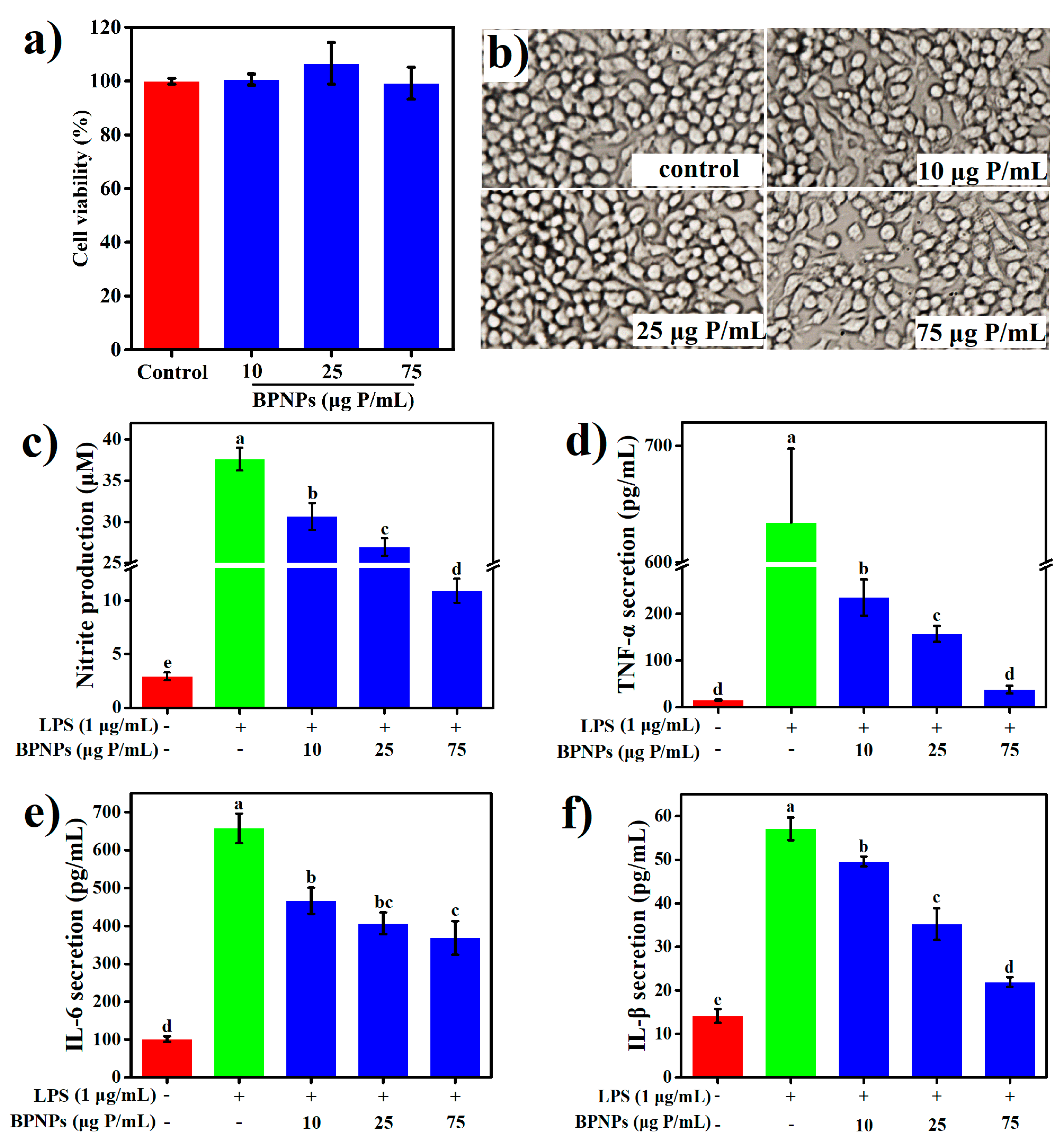
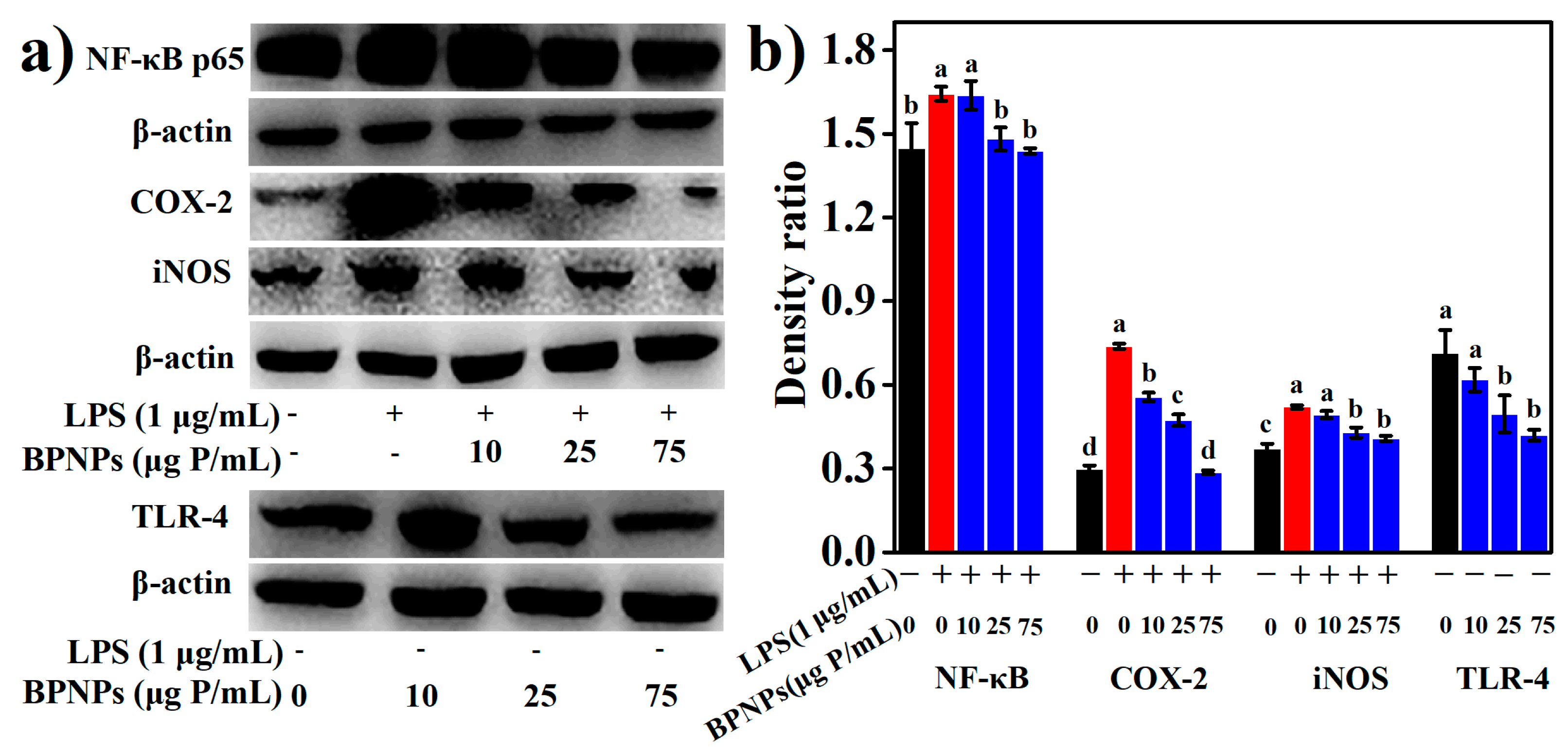
2.4. Anti-Inflammatory Effects of BPNPs in Lipopolysaccharide (LPS)-Activated RAW264.7 Cells
3. Conclusions
4. Experimental Section
4.1. Chemicals
4.2. Cyanobacterial Strains and Culture Conditions
4.3. PolyP Measurements
4.4. Molybdenum Blue Assay for Orthophosphate
4.5. Extraction and Purification of BPNPs
4.6. Electrophoretic Analysis of BPNPs
4.7. Characterization of BPNPs
4.8. Simulated GID
4.9. Cellular Experiments
4.10. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Kulaev, I.S.; Vagabov, V.M.; Kulakovskaya, T.V. The Biochemistry of Inorganic Polyphosphates; Springer International Publishing: Cham, Switzerland, 2005; pp. 3–35. [Google Scholar]
- Docampo, R.; de Souza, W.; Miranda, K.; Rohloff, P.; Moreno, S.N.J. Acidocalcisomes-Conserved from bacteria to man. Nat. Rev. Microbiol. 2005, 3, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Bonting, C.F.C.; Kortstee, G.J.J.; Boekestein, A.; Zehnder, A.J. The elemental composition dynamics of large polyphosphate granules in Acinetobacter strain 210A. Arch. Microbiol. 1993, 159, 428–434. [Google Scholar] [CrossRef]
- Racki, L.R.; Tocheva, E.I.; Dieterle, M.G.; Sullivan, M.C.; Jensen, G.J.; Newman, D.K. Polyphosphate granule biogenesis is temporally and functionally tied to cell cycle exit during starvation in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2017, 114, E2440–E2449. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, J.; Bru, S.; Ribeiro, M.P.C.; Clotet, J. Polyphosphate: Popping up from oblivion. Curr. Genet. 2017, 63, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Albi, T.; Serrano, A. Inorganic polyphosphate in the microbial world. Emerging roles for a multifaceted biopolymer. World J. Microbiol. Biotechnol. 2016, 32, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsilingiri, K.; Rescigno, M. Postbiotics: What else? Benef. Microbes 2013, 4, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Tsilingiri, K.; Barbosa, T.; Penna, G.; Caprioli, F.; Sonzogni, A.; Viale, G.; Rescigno, M. Probiotic and postbiotic activity in health and disease: Comparison on a novel polarised ex-vivo organ culture model. Gut 2012, 61, 1007–1015. [Google Scholar] [CrossRef] [PubMed]
- Segawa, S.; Fujiya, M.; Konishi, H.; Ueno, N.; Kobayashi, N.; Shigyo, T.; Kohgo, Y. Probiotic-Derived polyphosphate enhances the epithelial barrier function and maintains intestinal homeostasis through integrin-p38 MAPK pathway. PLoS ONE 2011, 6, e23278. [Google Scholar] [CrossRef] [PubMed]
- Kashima, S.; Fujiya, M.; Konishi, H.; Ueno, N.; Inaba, Y.; Moriichi, K.; Tanabe, H.; Ikuta, K.; Ohtake, T.; Kohgo, Y. Polyphosphate, an active molecule derived from probiotic Lactobacillus brevis, improves the fibrosis in murine colitis. Transl. Res. 2015, 166, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Fujiya, M.; Konishi, H.; Ueno, N.; Kashima, S.; Sasajima, J.; Moriichi, K.; Ikuta, K.; Tanabe, H.; Kohgo, Y. Probiotic-derived polyphosphate improves the intestinal barrier function through the caveolin-dependent endocytic pathway. Biochem. Biophys. Res. Commun. 2015, 467, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Sakatani, A.; Fujiya, M.; Ueno, N.; Kashima, S.; Sasajima, J.; Moriichi, K.; Ikuta, K.; Tanabe, H.; Kohgo, Y. Polyphosphate derived from lactobacillus brevis inhibits colon cancer progression through induction of cell apoptosis. Anticancer Res. 2016, 36, 591–598. [Google Scholar] [PubMed]
- Oehmen, A.; Zeng, R.J.; Yuan, Z.G.; Keller, J. Anaerobic metabolism of propionate by polyphosphate-accumulating organisms in enhanced biological phosphorus removal systems. Biotechnol. Bioeng. 2005, 91, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Dyhrman, S.T. Nutrients and their acquisition: Phosphorus physiology in microalgae. In Developments in Applied Phycology; Borowitzka, M., Beardall, J., Raven, J., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 6, pp. 155–183. [Google Scholar]
- Ludwig, M.; Bryant, D.A. Transcription profiling of the model cyanobacterium Synechococcus sp. strain PCC 7002 by Next-Gen (SOLiD™) sequencing of cDNA. Front. Microb. 2011, 2, 41. [Google Scholar] [CrossRef] [PubMed]
- Marcus, L.; Bryant, D.A. Acclimation of the global transcriptome of the cyanobacterium Synechococcus sp. strain pcc 7002 to nutrient limitations and different nitrogen sources. Front. Microb. 2012, 3, 145. [Google Scholar]
- Nomura, C.T.; Sakamoto, T.; Bryant, D.A. Roles for heme-copper oxidases in extreme high-light and oxidative stress response in the cyanobacterium Synechococcus sp. PCC 7002. Arch. Microbiol. 2006, 185, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, M.; Bryant, D.A. Synechococcus sp strain PCC 7002 transcriptome: Acclimation to temperature, salinity, oxidative stress, and mixotrophic growth conditions. Front. Microb. 2012, 3, 354. [Google Scholar] [CrossRef] [PubMed]
- Quintana, N.; Van der Kooy, F.; Van de Rhee, M.D.; Voshol, G.P.; Verpoorte, R. Renewable energy from Cyanobacteria: Energy production optimization by metabolic pathway engineering. Appl. Microbiol. Biotechnol. 2011, 91, 471–490. [Google Scholar] [CrossRef] [PubMed]
- Kulakova, A.N.; Hobbs, D.; Smithen, M.; Pavlov, E.; Gilbert, J.A.; Quinn, J.P.; McGrath, J.W. Direct quantification of inorganic polyphosphate in microbial cells using 4′-6-diamidino-2-phenylindole (DAPI). Environ. Sci. Technol. 2011, 45, 7799–7803. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, F.; Leppard, G.G.; Kushnert, D.J. Copper resistance in Anabaena variabilis: Effects of phosphate nutrition and polyphosphate bodies. Microb. Ecol. 1994, 27, 159–176. [Google Scholar] [CrossRef] [PubMed]
- Seki, Y.; Nitta, K.; Kaneko, Y. Observation of polyphosphate bodies and DNA during the cell division cycle of Synechococcus elongatus PCC 7942. Plant Biol. 2014, 16, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.E.; Baxter, M.; Rachlin, J.W.; Jani, V. Uptake of heavy metals by Plectonema boryanum (cyanophyceae) into cellular components, especially polyphosphate bodies: An X-ray energy dispersive study. Environ. Pollut. 1982, 27, 119–127. [Google Scholar] [CrossRef]
- Jensen, T.E. Electron microscopy of polyphosphate bodies in a blue-green alga, Nostoc pruniforme. Arch. Microbiol 1968, 62, 144–152. [Google Scholar] [CrossRef]
- Mou, S.; Zhang, Y.; Li, G.; Li, H.; Liang, Y.; Tang, L.; Tao, J.; Xu, J.; Li, J.; Zhang, C. Effects of elevated CO2 and nitrogen supply on the growth and photosynthetic physiology of a marine cyanobacterium, Synechococcus sp. PCC7002. J. Appl. Phycol. 2017, 29, 1755–1763. [Google Scholar] [CrossRef]
- Acosta, E. Bioavailability of nanoparticles in nutrient and nutraceutical delivery. Curr. Opin. Colloid Interface Sci. 2009, 14, 3–15. [Google Scholar] [CrossRef]
- Kulakovskaya, T.V.; Andreeva, N.A.; Trilisenko, L.V.; Vagabov, V.M.; Kulaev, I.S. Two exopolyphosphatases in Saccharomyces cerevisiae cytosol at different culture conditions. Process Biochem. 2004, 39, 1625–1630. [Google Scholar] [CrossRef]
- Ogawa, N.; Tzeng, C.-M.; Fraley, C.d.; Kornberg, A. Inorganic polyphosphate in Vibrio cholerae: Genetic, biochemical, and physiologic features. J. Bacteriol. 2000, 182, 6687–6693. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Litaker, R.W.; Sunda, W.G.; Wood, M. Phosphorus physiological ecology and molecular mechanisms in marine phytoplankton. J. Phycol. 2016, 52, 10–36. [Google Scholar] [CrossRef] [PubMed]
- Rao, N.N.; Gómez-García, M.R.; Kornberg, A. Inorganic polyphosphate: Essential for growth and survival. Annu. Rev. Biochem. 2009, 78, 605–647. [Google Scholar] [CrossRef] [PubMed]
- Lamarche, M.G.; Wanner, B.L.; Crépin, S.; Harel, J. The phosphate regulon and bacterial virulence: A regulatory network connecting phosphate homeostasis and pathogenesis. FEMS Microbiol. Rev. 2008, 32, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Gomezgarcia, M.R.; Fazeli, F.; Grote, A.; Grossman, A.R.; Bhaya, D. Role of polyphosphate in thermophilic Synechococcus sp. from microbial mats. J. Bacteriol. 2013, 195, 3309–3319. [Google Scholar] [CrossRef] [PubMed]
- Harold, F.M.; Sylvan, S. Accumulation of inorganic polyphosphate in Aerobacter aerogenes II. environmental control and the role of sulfur compounds. J. Bacteriol. 1963, 86, 216–221. [Google Scholar]
- Santosbeneit, F. The Pho regulon: A huge regulatory network in bacteria. Front. Microb. 2015, 6, 402. [Google Scholar]
- Tocheva, E.I.; Dekas, A.E.; Mcglynn, S.E.; Morris, D.; Orphan, V.J.; Jensen, G.J. Polyphosphate storage during sporulation in the gram-negative bacterium acetonema longum. J. Bacteriol. 2013, 195, 3940. [Google Scholar] [CrossRef] [PubMed]
- Strauss, U.P.; Treitler, T.L. Degradation of polyphosphates in solution. I. kinetics and mechanism of the hydrolysis at branching points in polyphosphate chains. J. Am. Chem. Soc. 1956, 78, 3553–3557. [Google Scholar] [CrossRef]
- Ivey, F.J.; Shaver, K. Enzymic hydrolysis of polyphosphate in the gastrointestinal tract. J. Agric. Food Chem. 1976, 25, 128. [Google Scholar] [CrossRef] [PubMed]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef] [PubMed]
- Fahmi, H.; Charon, D.; Mondange, M.; Chaby, R. Endotoxin-induced desensitization of mouse macrophages is mediated in part by nitric oxide production. Infect. Immun. 1995, 63, 1863–1869. [Google Scholar] [PubMed]
- Husebye, H.; Halaas, O.; Stenmark, H.; Tunheim, G.; Sandanger, O.; Bogen, B.; Brech, A.; Latz, E.; Espevik, T. Endocytic pathways regulate Toll-like receptor 4 signaling and link innate and adaptive immunity. Embo J. 2006, 25, 683–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.M.; Li, W.Q.; Wang, F.; Sun, C.Y.; Wang, L.; Wang, J.; Sun, F. Enhancement of lipopolysaccharide-induced nitric oxide and interleukin-6 production by PEGylated gold nanoparticles in RAW264.7 cells. Nanoscale 2012, 4, 7135–7142. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-Y.; Lu, S.-L.; Hu, C.-W.; Yeh, C.-S.; Lee, G.-B.; Lei, H.-Y. Size-Dependent attenuation of TLR9 signaling by gold nanoparticles in macrophages. J. Immunol. 2012, 188, 68. [Google Scholar] [CrossRef] [PubMed]
- Stevens Jr, S.E.; Patterson, C.O.P.; Myers, J. The production of hydrogen peroxide by blue-green algae: A survey. J. Phycol. 1973, 9, 427–430. [Google Scholar] [CrossRef]
- Martin, P.; Van Mooy, B.A. Fluorometric quantification of polyphosphate in environmental plankton samples: Extraction protocols, matrix effects, and nucleic acid interference. Appl. Environ. Microbiol. 2013, 79, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Kumble, K.D.; Kornberg, A. Inorganic polyphosphate in mammalian cells and tissues. J. Biol. Chem. 1995, 270, 5818–5822. [Google Scholar] [CrossRef] [PubMed]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.O.R.S.T.E.N.; Bourlieu, C.; Dufour, C. A standardised static in vitro digestion method suitable for food-an international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15N] nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, G.; Dong, S.; Huang, M.; Zeng, M.; Liu, Z.; Zhao, Y.; Wu, H. Biogenic Polyphosphate Nanoparticles from a Marine Cyanobacterium Synechococcus sp. PCC 7002: Production, Characterization, and Anti-Inflammatory Properties In Vitro. Mar. Drugs 2018, 16, 322. https://doi.org/10.3390/md16090322
Feng G, Dong S, Huang M, Zeng M, Liu Z, Zhao Y, Wu H. Biogenic Polyphosphate Nanoparticles from a Marine Cyanobacterium Synechococcus sp. PCC 7002: Production, Characterization, and Anti-Inflammatory Properties In Vitro. Marine Drugs. 2018; 16(9):322. https://doi.org/10.3390/md16090322
Chicago/Turabian StyleFeng, Guangxin, Shiyuan Dong, Min Huang, Mingyong Zeng, Zunying Liu, Yuanhui Zhao, and Haohao Wu. 2018. "Biogenic Polyphosphate Nanoparticles from a Marine Cyanobacterium Synechococcus sp. PCC 7002: Production, Characterization, and Anti-Inflammatory Properties In Vitro" Marine Drugs 16, no. 9: 322. https://doi.org/10.3390/md16090322