
Recombinant Expression and Characterization of α-Conotoxin LvIA in Escherichia coli


Abstract
:
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| α m/n | Name | Species | Sequence | Target | Reference |
|---|---|---|---|---|---|
| α 3/5 | GI | C. geographus | ECCNPACGRHYSC * | α1β1γδ | [10] |
| MI | C. magus | GRCCHPACGKNYSC * | α1β1γδ | [11] | |
| α 4/3 | ImI | C. imperialis | GCCSDPRCAWRC * | α3β2, α7 | [12] |
| ImII | C. imperialis | ACCSDRRCRWRC * | α7 | [12] | |
| α 4/4 | BuIA | C. bullatus | GCCSTPPCAVLYC * | α6/α3β2, α6/α3β4, α3β2, α3β4 | [13] |
| α 4/6 | TxIA | C. textile | GCCSROOCIANNPDLC * | α3β2, α7 | [14] |
| TxID | C. textile | GCCSHPVCSAMSPIC * | α3β4, α6/α3β4 | [15] | |
| α 4/7 | LvIA | C. lividus | GCCSHPACNVDHPEIC * | α3β2, α3β4, α6β4 | [9] |
| GIC | C. geographus | GCCSHPACAGNNQHIC * | α3β2 | [16] | |
| Vc1.1 | C. victoriae | GCCSDPRCNYDHPEIC * | α9α10 | [17] |
2. Results
2.1. Construction of the LvIA Recombinant Vector


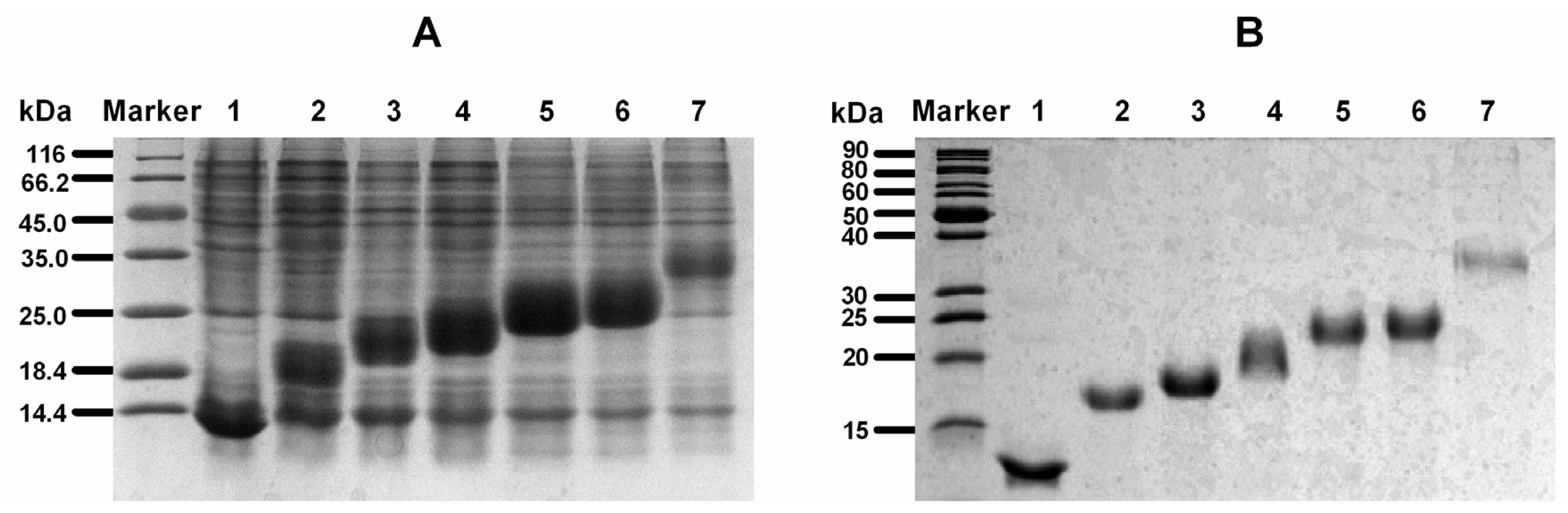
2.2. Expression and Purification of Recombinant Protein

2.3. CNBr Cleavage and Purification of Recombinant Protein
2.4. Bioassay of rLvIA in Pain Model


2.5. Effect of rLvIA on ACh-Evoked nAChR-Mediated Currents


3. Discussion
| KSI(rLvIA)nHis6 a | The Yield of Fusion Protein of KSI(rLvIA)nHis6 (mg/L) b | The Yield of Purified rLvIA (mg/L) c |
|---|---|---|
| KSI(rLvIA)1His6 | 114.1 | 3.8 |
| KSI(rLvIA)2His6 | 553.2 | 15.3 |
| KSI(rLvIA)3His6 | 115.1 | 7.8 |
| KSI(rLvIA)4His6 | 121.3 | 9.3 |
| KSI(rLvIA)5His6 | 156.5 | 11.2 |
| KSI(rLvIA)6His6 | 290.9 | 40.8 |
| KSI(rLvIA)11His6 | 534.5 | 45.3 |
| Conotoxin | Origin | Native Peptide Sequence | Fused Partner | Recombinant Peptide | Yield (mg/L) | Activity/Target | IC50 | Reference |
|---|---|---|---|---|---|---|---|---|
| LvIA | C. lividus | GCCSHPACNVDHPEIC * | KSI | rLvIA | 45 | Analgesic/α3β2 nAChRs | 160.8 nM(rα32β)/46.8 nM(hα32β) | This work |
| MVIIA | C. magus | CKGKGAKCSRLMYDCCTGSCRSGKC | TRX | Trx-CTX MVIIA | 40 | analgesic function | No data | [33] |
| MVIIA | C.magus | CKGKGAKCSRLMYDCCTGSCRSGKC | GST | GST-CTX MVIIA | No data | Analgesic Activity | No data | [34] |
| Mol659 | C.monile | FHGGSWYRFPWGY | Cytochrome b5 | Mol659 | 6–8 | K+ channel | No data | [32] |
| Conkunitzin-S1 | C. striatus | KDRPSLCDLPADSGSGTKAEKRIYYNSAR KQCLRFDYTGQGGNENNFRRTYDCQRTCLYT | Ssp DnaB intein | Conk-S1 | No data | K+ channel | 1.33 nM | [35] |
| lt7a | C. litteratus | CLGWSNYCTSHSICCSGECILSYCDIW | TRX | lt7a | 6 | Na+ channel | No data | [36] |
| lt6c | C. litteratus | WPCKVAGSPCGLVSECCGTCNVLRNRCV | TRX | lt6c | 12 | Na+ channel | No data | [37] |
| TxVIA | C. textile | WCKQSGEMCNLLDQNCCDGYCIVLVCT | Alpha factor | Pro-TxVIA | 10 | insecticidal activity | No data | [8] |
| PrIIIE | C. parius | AARCCTYHGSCLKEKCRRKYCCG | SUMO | PrIIIE | 1.5 | α1β1δε/α1β1γδ | 2.8 μM | [7] |
| Vn2 | C. ventricosus | EDCIAVGQLCVFWNIGRPCCSGLCVFACTVKLP | GST | GST-wtCTX GST-mtCTX | No data | Toxicity to insects | No data | [38] |
| SIIIA | C. striatus | ENCCNGGCSSKWCRDHARCC | FlgM | rSIIIA | No data | Na+ channel | No data | [39] |
| GeXIVAWT | C. generalis | TCRSSGRYCRSPYDCRRRYCRRITDACV | PelB | rPelB-GeXIVAWT-His | 61.6 | cytotoxicity | No data | [40] |
| MrVIB | C. marmoreus | ACSKKWEYCIVPILGFVYCCPGLICGPFVCV | PelB | rMrVIB-His | 5.9 | Analgesic activity | No data | [29] |
| Lt15a | C. litteratus | ECTTKHRRCEKDEECCPNLECKCLTSPDCQSGYKCKP | TRX | Lt15a | No data | mice coma state, crouched. | No data | [41] |
| lt16a | C. litteratus | TGEDFLEECMGGCAFDFCCKR | TRX | lt16a | No data | Na+ channel | No data | [42] |
4. Experimental Section
4.1. Materials
4.2. Construction of Recombinant Exptession Vectors of LvIAn-His·Tag Genes
- (1)
- 5’-GGTTGCTGCTCTCACCCGGCTTGCAACGTTGACCACCCGGAAATCTGCATG-3’,
- (2)
- 3’-TACCCAACGACGAGAGTGGGCCGAACGTTGCAACTGGTGGGCCTTTAGACG-5’.
4.3. Expression and Purification of KSI-LvIAn-His·Tag Fusion Protein
4.4. CNBr Cleavage and Purification of Recombinant Protein
4.5. Bioassays of rLvIA in Pain Model
4.6. Electrophysiology and Data Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CTx | conotoxin |
| nAChRs | nicotinic acetylcholine receptors |
| KSI | ketosteroid isomerase |
| CNBr | cyanogen bromide |
| CIAP | calf intestine alkaline phosphatase |
| IPTG | isopropyl β- d-1-thiogalactopyranoside |
| RP-HPLC | reversed phase high-performance liquid chromatography |
| ESI-MS | electrospray ionization-mass spectrometer |
| SDS-PAGE | sodium dodecyl sulfate polyacrylamide gel electropheresis |
| TFA | trifluoroacetic acid |
| ACN | acetonitrile |
| NSS | normal saline solution |
| LB | Lysogeny Broth |
References
- Olivera, B.M.; Teichert, R.W. Diversity of the neurotoxic Conus peptides: A model for concerted pharmacological discovery. Mol. Interv. 2007, 7, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.D.; Norton, R.S. Conotoxin gene superfamilies. Mar. Drugs 2014, 12, 6058–6101. [Google Scholar] [CrossRef] [PubMed]
- Azam, L.; McIntosh, J.M. Alpha-conotoxins as pharmacological probes of nicotinic acetylcholine receptors. Acta Pharmacol. Sin. 2009, 30, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Janes, R.W. α-Conotoxins as selective probes for nicotinic acetylcholine receptor subclasses. Curr. Opin. Pharmacol. 2005, 5, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Essack, M.; Bajic, V.B.; Archer, J.A. Conotoxins that confer therapeutic possibilities. Mar. Drugs 2012, 10, 1244–1265. [Google Scholar] [CrossRef] [PubMed]
- Armishaw, C.J.; Jensen, A.A.; Skonberg, C.; Liljefors, T.; Stromgaard, K. Synthesis and biological activity of novel alpha-conotoxin analogues incorporating substituted proline derivatives. Adv. Exp. Med. Biol. 2009, 611, 179–180. [Google Scholar] [PubMed]
- Hernandez-Cuebas, L.M.; White, M.M. Expression of a biologically-active conotoxin PrIIIE in Escherichia coli. Protein Expr. Purif. 2012, 82, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Bruce, C.; Fitches, E.C.; Chougule, N.; Bell, H.A.; Gatehouse, J.A. Recombinant conotoxin, TxVIA, produced in yeast has insecticidal activity. Toxicon 2011, 58, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Zhangsun, D.; Schroeder, C.I.; Zhu, X.; Hu, Y.; Wu, Y.; Weltzin, M.M.; Eberhard, S.; Kaas, Q.; Craik, D.J.; et al. A novel α4/7-conotoxin LvIA from Conus lividus that selectively blocks α3β2 vs. α6/α3β2β3 nicotinic acetylcholine receptors. FASEB J. 2014, 28, 1842–1853. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.S.; Martinez, J.; Elgoyhen, A.B.; Heinemann, S.F.; McIntosh, J.M. α-Conotoxin ImI exhibits subtype-specific nicotinic acetylcholine receptor blockade: Preferential inhibition of homomeric alpha 7 and alpha 9 receptors. Mol. Pharmacol. 1995, 48, 194–199. [Google Scholar] [PubMed]
- Jacobsen, R.B.; DelaCruz, R.G.; Grose, J.H.; McIntosh, J.M.; Yoshikami, D.; Olivera, B.M. Critical residues influence the affinity and selectivity of alpha-conotoxin MI for nicotinic acetylcholine receptors. Biochemistry 1999, 38, 13310–13315. [Google Scholar] [CrossRef] [PubMed]
- Ellison, M.; Gao, F.; Wang, H.L.; Sine, S.M.; McIntosh, J.M.; Olivera, B.M. Alpha-conotoxins ImI and ImII target distinct regions of the human alpha7 nicotinic acetylcholine receptor and distinguish human nicotinic receptor subtypes. Biochemistry 2004, 43, 16019–16026. [Google Scholar] [CrossRef] [PubMed]
- Azam, L.; Dowell, C.; Watkins, M.; Stitzel, J.A.; Olivera, B.M.; McIntosh, J.M. Alpha-conotoxin BuIA, a novel peptide from Conus bullatus, distinguishes among neuronal nicotinic acetylcholine receptors. J. Biol. Chem. 2005, 280, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Ulens, C.; Buttner, R.; Fish, A.; van Elk, R.; Kendel, Y.; Hopping, G.; Alewood, P.F.; Schroeder, C.; Nicke, A.; et al. AChBP-targeted alpha-conotoxin correlates distinct binding orientations with nAChR subtype selectivity. EMBO J. 2007, 26, 3858–3867. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Zhangsun, D.; Zhu, X.; Wu, Y.; Hu, Y.; Christensen, S.; Harvey, P.J.; Akcan, M.; Craik, D.J.; McIntosh, J.M. Characterization of a novel alpha-conotoxin TxID from Conus textile that potently blocks rat alpha3beta4 nicotinic acetylcholine receptors. J. Med. Chem. 2013, 56, 9655–9663. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, J.M.; Dowell, C.; Watkins, M.; Garrett, J.E.; Yoshikami, D.; Olivera, B.M. Alpha-conotoxin GIC from Conus geographus, a novel peptide antagonist of nicotinic acetylcholine receptors. J. Biol. Chem. 2002, 277, 33610–33615. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.J.; Fischer, H.; Nevin, S.T.; Adams, D.J.; Craik, D.J. The synthesis, structural characterization, and receptor specificity of the alpha-conotoxin Vc1.1. J. Biol. Chem. 2006, 281, 23254–23263. [Google Scholar] [CrossRef] [PubMed]
- Muttenthaler, M.; Akondi, K.B.; Alewood, P.F. Structure-activity studies on alpha-conotoxins. Curr. Pharm. Des. 2011, 17, 4226–4241. [Google Scholar] [CrossRef] [PubMed]
- Hurst, R.; Rollema, H.; Bertrand, D. Nicotinic acetylcholine receptors: From basic science to therapeutics. Pharmacol. Ther. 2013, 137, 22–54. [Google Scholar] [CrossRef] [PubMed]
- Olivera, B.M.; Quik, M.; Vincler, M.; McIntosh, J.M. Subtype-selective conopeptides targeted to nicotinic receptors: Concerted discovery and biomedical applications. Channels (Austin) 2008, 2, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Zhangsun, D.; Wu, Y.; Zhu, X.; Hu, Y.; McIntyre, M.; Christensen, S.; Akcan, M.; Craik, D.J.; McIntosh, J.M. Characterization of a novel alpha-conotoxin from conus textile that selectively targets alpha6/alpha3beta2beta3 nicotinic acetylcholine receptors. J. Biol. Chem. 2013, 288, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Zhangsun, D.; Zhu, X.; Wu, Y.; Hu, Y.; Kaas, Q.; Craik, D.J.; McIntosh, J.M.; Luo, S. Key residues in the nicotinic acetylcholine receptor beta2 subunit contribute to alpha-conotoxin LvIA binding. J. Biol. Chem. 2015, 290, 9855–9862. [Google Scholar] [CrossRef] [PubMed]
- Young, T.; Wittenauer, S.; McIntosh, J.M.; Vincler, M. Spinal alpha3beta2* nicotinic acetylcholine receptors tonically inhibit the transmission of nociceptive mechanical stimuli. Brain Res. 2008, 1229, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Majerle, A.; Kidric, J.; Jerala, R. Production of stable isotope enriched antimicrobial peptides in Escherichia coli: An application to the production of a 15N-enriched fragment of lactoferrin. J. Biomol. NMR 2000, 18, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Kuliopulos, A.; Walsh, C.T. Production, purification, and cleavage of tandem repeats of recombinant peptides. J. Am. Chem. Soc. 1994, 116, 4599–4507. [Google Scholar] [CrossRef]
- Hwang, P.M.; Pan, J.S.; Sykes, B.D. Targeted expression, purification, and cleavage of fusion proteins from inclusion bodies in Escherichia coli. FEBS Lett. 2014, 588, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Forsberg, G.; Moks, T.; Hartmanis, M.; Nilsson, B. Fusion proteins in biotechnology. Curr. Opin. Biotechnol. 1992, 3, 363–369. [Google Scholar] [CrossRef]
- Menendez, L.; Lastra, A.; Hidalgo, A.; Baamonde, A. Unilateral hot plate test: A simple and sensitive method for detecting central and peripheral hyperalgesia in mice. J. Neurosci. Methods 2002, 113, 91–97. [Google Scholar] [CrossRef]
- Gao, B.; Zhangsun, D.; Hu, Y.; Wu, Y.; Sheng, L.; Fang, L.; Wu, X.; Yu, J.; Luo, S. Expression and secretion of functional recombinant µO-conotoxin MrVIB-His-tag in Escherichia coli. Toxicon 2013, 72, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.G.; Bandyopadhyay, P.; Olivera, B.M. Post-translationally modified neuropeptides from Conus venoms. Eur. J. Biochem. 1999, 264, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.S.; Vivekanandan, S.; Jois, S.D.; Kini, R.M. Effect of C-terminal amidation on folding and disulfide-pairing of alpha-conotoxin ImI. Angew. Chem. Int. Ed. Engl. 2005, 44, 6333–6337. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.S.; Ramasamy, P.; Sikdar, S.K.; Sarma, S.P. Overexpression, purification, and pharmacological activity of a biosynthetically derived conopeptide. Biochem. Biophys. Res. Commun. 2005, 335, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Zhan, J.; Chen, X.; Wang, C.; Qiu, J.; Ma, F.; Wang, K.; Zheng, S. A fusion protein of conotoxin MVIIA and thioredoxin expressed in Escherichia coli has significant analgesic activity. Biochem. Biophys. Res. Commun. 2003, 311, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Chen, Y.; Zhu, Y.; Wang, F.; Xu, X.; Zhan, J. Recombinant omega-conotoxin MVIIA possesses strong analgesic activity. BioDrugs 2006, 20, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Bayrhuber, M.; Graf, R.; Ferber, M.; Zweckstetter, M.; Imperial, J.; Garrett, J.E.; Olivera, B.M.; Terlau, H.; Becker, S. Production of recombinant Conkunitzin-S1 in Escherichia coli. Protein Expr. Purif. 2006, 47, 640–644. [Google Scholar] [CrossRef] [PubMed]
- Pi, C.; Liu, J.; Wang, L.; Jiang, X.; Liu, Y.; Peng, C.; Chen, S.; Xu, A. Soluble expression, purification and functional identification of a disulfide-rich conotoxin derived from Conus litteratus. J. Biotechnol. 2007, 128, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Pi, C.; Liu, J.; Chen, S.; Peng, C.; Sun, D.; Zhou, M.; Xiang, H.; Ren, Z.; Xu, A. Identification and characterization of a novel O-superfamily conotoxin from Conus litteratus. J. Pept. Sci. 2008, 14, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Spiezia, M.C.; Chiarabelli, C.; Polticelli, F. Recombinant expression and insecticidal properties of a Conus ventricosus conotoxin-GST fusion protein. Toxicon 2012, 60, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Singer, H.M.; Erhardt, M.; Steiner, A.M.; Zhang, M.M.; Yoshikami, D.; Bulaj, G.; Olivera, B.M.; Hughes, K.T. Selective purification of recombinant neuroactive peptides using the flagellar type III secretion system. mBio 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Zhangsun, D.; Wu, Y.; Lin, B.; Zhu, X.; Luo, S. Expression, renaturation and biological activity of recombinant conotoxin GeXIVAWT. Appl. Microbiol. Biotechnol. 2013, 97, 1223–1230. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, L.; Zhou, M.; Jiang, X.; Zhu, X.; Chen, Y.; Luo, S.; You, Y.; Ren, Z.; Xu, A. Soluble expression, purification and functional identification of the framework XV conotoxins derived from different Conus species. Peptides 2014, 56, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Wang, L.; Wu, Y.; Liu, J.; Sun, D.; Zhu, X.; Feng, Y.; Qin, M.; Chen, S.; Xu, A. Soluble expression and sodium channel activity of lt16a, a novel framework XVI conotoxin from the M-superfamily. Toxicon 2015, 98, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Zhangsun, D.; Zhang, B.; Quan, Y.; Wu, Y. Novel alpha-conotoxins identified by gene sequencing from cone snails native to Hainan, and their sequence diversity. J. Pept. Sci. 2006, 12, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Cartier, G.E.; Yoshikami, D.; Gray, W.R.; Luo, S.; Olivera, B.M.; McIntosh, J.M. A new alpha-conotoxin which targets alpha3beta2 nicotinic acetylcholine receptors. J. Biol. Chem. 1996, 271, 7522–7528. [Google Scholar] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, X.; Bi, J.; Yu, J.; Li, X.; Zhang, Y.; Zhangsun, D.; Luo, S. Recombinant Expression and Characterization of α-Conotoxin LvIA in Escherichia coli. Mar. Drugs 2016, 14, 11. https://doi.org/10.3390/md14010011
Zhu X, Bi J, Yu J, Li X, Zhang Y, Zhangsun D, Luo S. Recombinant Expression and Characterization of α-Conotoxin LvIA in Escherichia coli. Marine Drugs. 2016; 14(1):11. https://doi.org/10.3390/md14010011
Chicago/Turabian StyleZhu, Xiaopeng, Jianpeng Bi, Jinpeng Yu, Xiaodan Li, Yaning Zhang, Dongting Zhangsun, and Sulan Luo. 2016. "Recombinant Expression and Characterization of α-Conotoxin LvIA in Escherichia coli" Marine Drugs 14, no. 1: 11. https://doi.org/10.3390/md14010011