

Global Health Priority Box: Discovering Flucofuron as a Promising Antikinetoplastid Compound
 ,
,
Abstract
:1. Introduction
2. Results
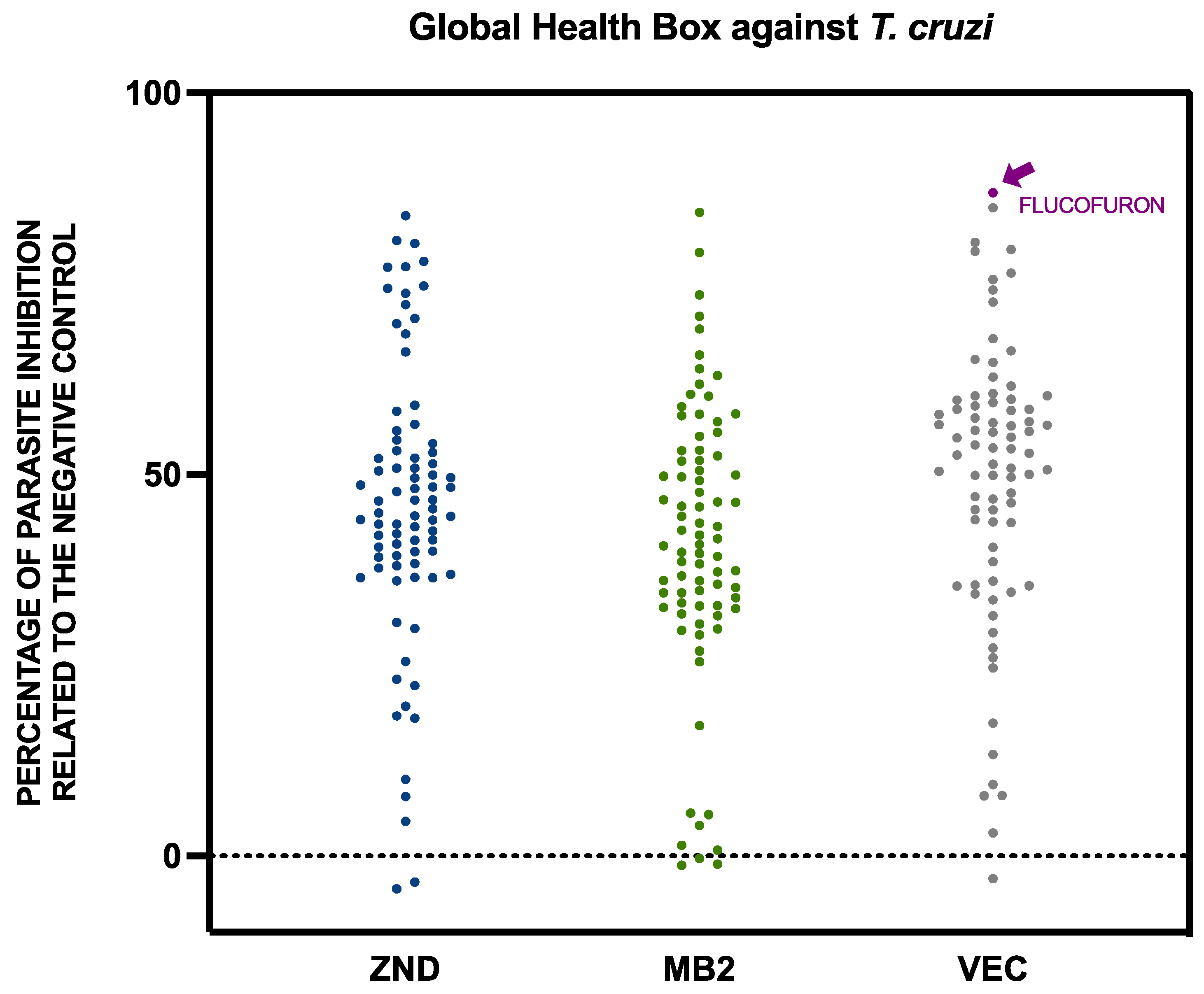
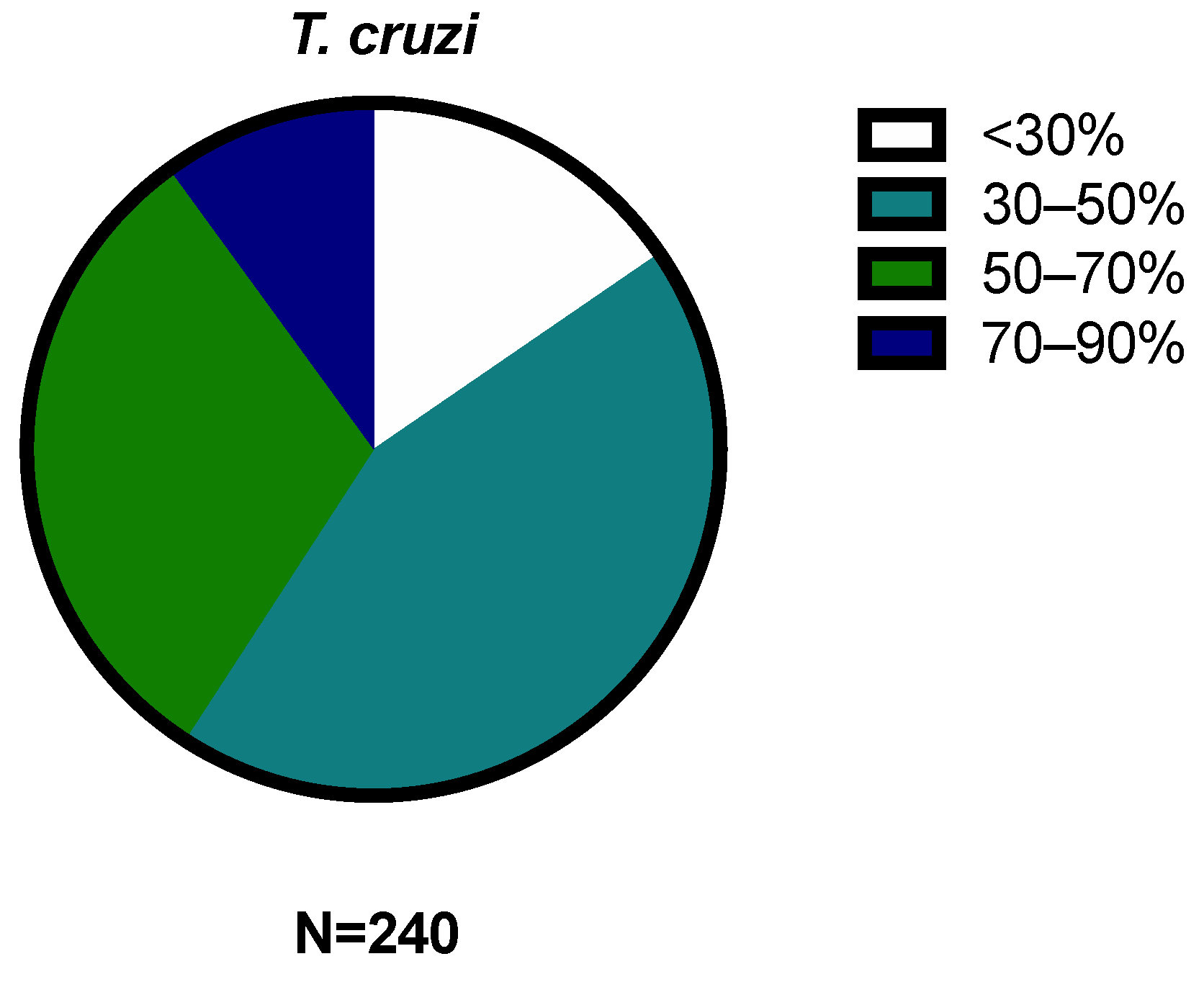
2.1. Activity against Trypanosoma cruzi and Leishmania amazonensis
2.2. Cytotoxicity against Murine Macrophages
2.3. Mechanisms of Cell Death
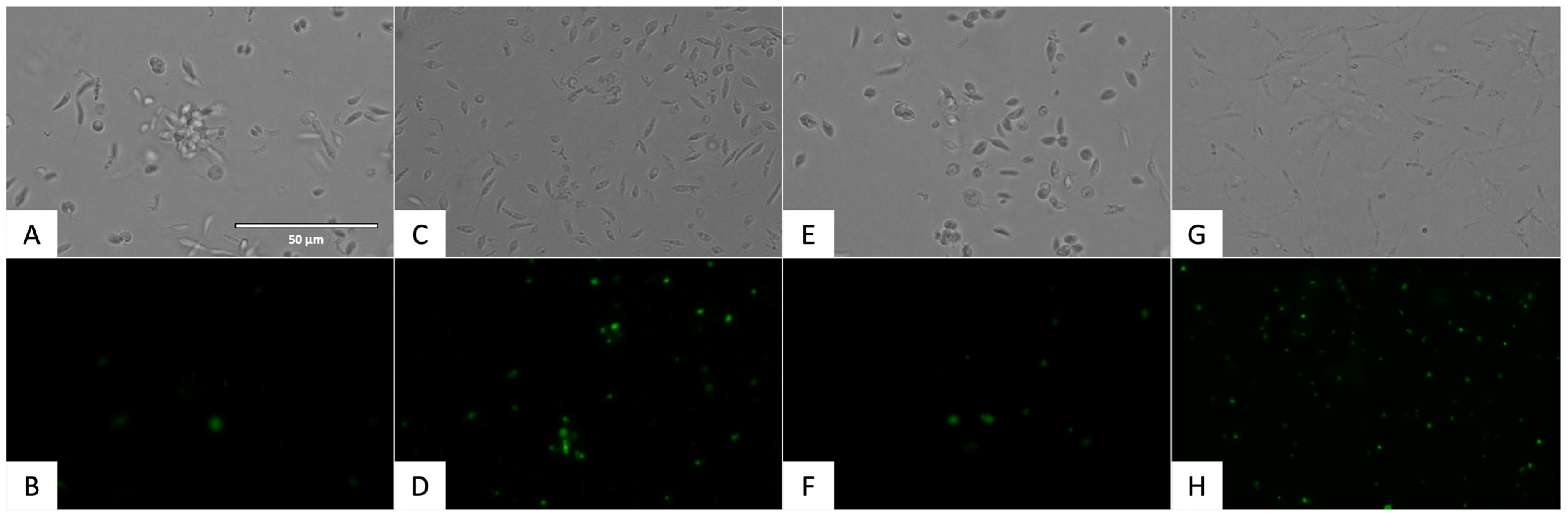
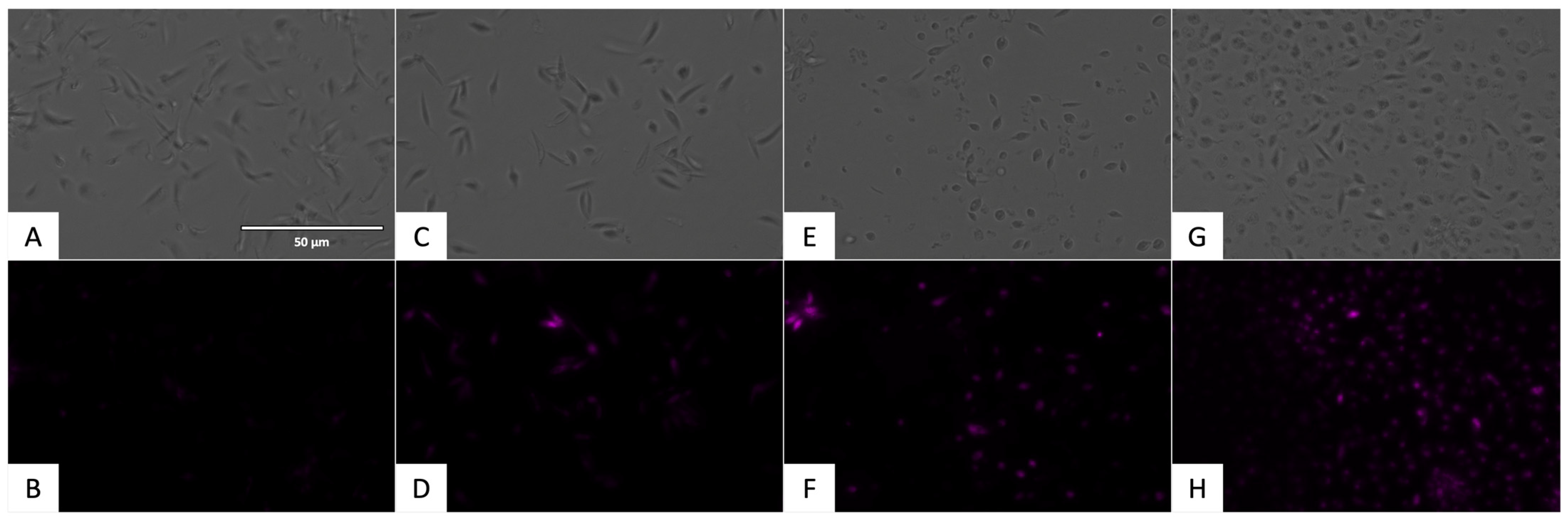
2.3.1. Chromatin Condensation Analysis
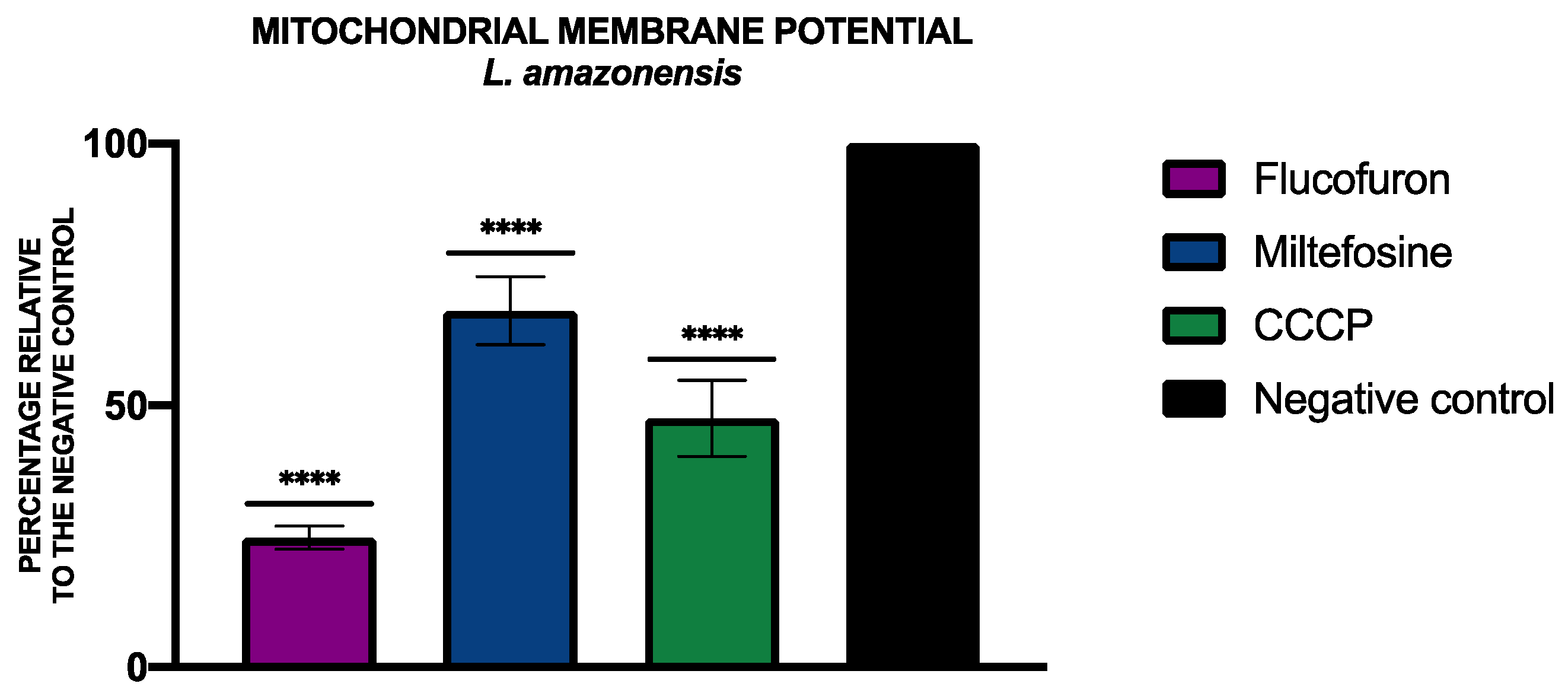
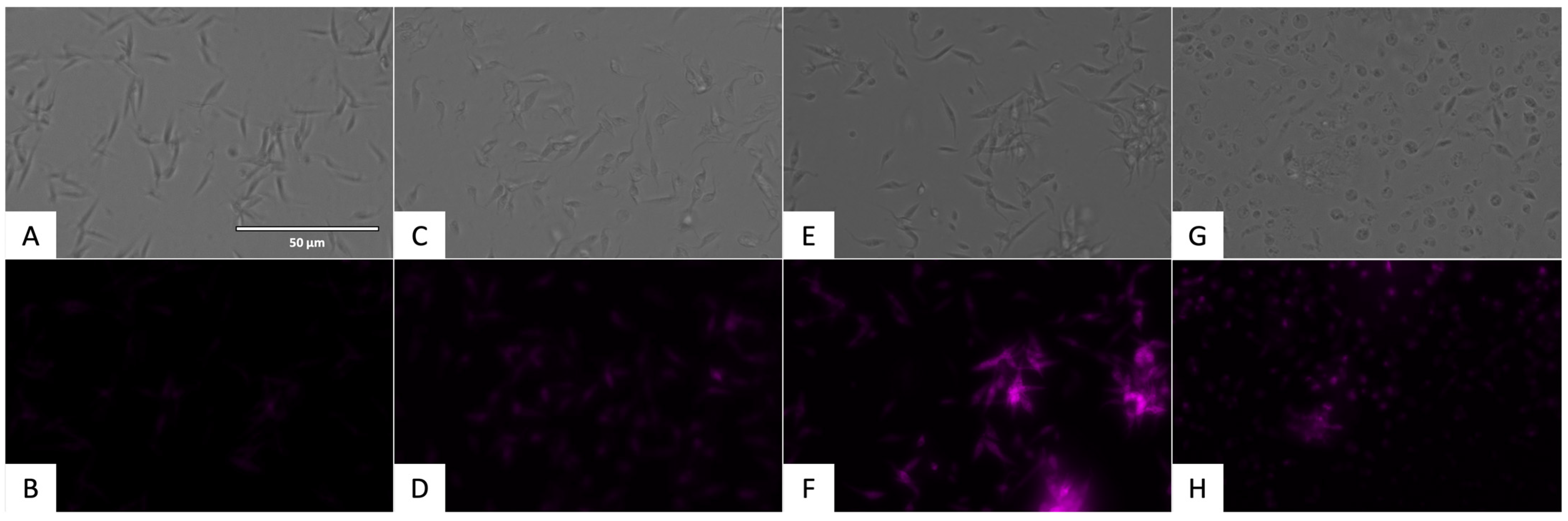
2.3.2. Mitochondrial Membrane Potential Analysis
2.3.3. Analysis of ATP Levels
2.3.4. Integrity of the Membrane Permeability Analysis
2.3.5. Analysis of the Presence of Reactive Oxygen Species
3. Discussion
4. Materials and Methods
4.1. Products
4.2. Cultures
4.3. Antiparasitic Activity
4.4. Cytotoxicity against Murine Macrophages
4.5. Mechanisms of Cell Death
4.6. Chromatin Condensation Analysis
4.7. Mitochondrial Membrane Potential Analysis
4.8. Analysis of ATP Levels
4.9. Integrity of the Membrane Permeability Analysis
4.10. Analysis of the Presence of Reactive Oxygen Species
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Ending the Neglect to Attain the Sustainable Development Goals: A Road Map for Neglected Tropical Diseases 2021–2030; WHO: Geneva, Switzerland, 2020; ISBN 9789240010352. [Google Scholar]
- de Vries, H.J.C.; Schallig, H.D. Cutaneous Leishmaniasis: A 2022 Updated Narrative Review into Diagnosis and Management Developments. Am. J. Clin. Dermatol. 2022, 23, 823–840. [Google Scholar] [CrossRef] [PubMed]
- Bern, C. Chagas’ Disease. N. Engl. J. Med. 2015, 373, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Organizacion Mundial de la Salud (OMS). Hoja de Ruta Sobre Enfermedades Tropicales Desatendidas 2021–2030; Organizacion Mundial de la Salud (OMS): Geneva, Switzerland, 2021; ISBN 9789240026520. [Google Scholar]
- Pérez-Molina, J.A.; Crespillo-Andújar, C.; Bosch-Nicolau, P.; Molina, I. Trypanocidal treatment of Chagas disease. Enfermedades Infecc. Y Microbiol. Clin. 2021, 39, 458–470. [Google Scholar] [CrossRef]
- de Fernandes, V.S.; da Rosa, R.; Zimmermann, L.A.; Rogério, K.R.; Kümmerle, A.E.; Bernardes, L.S.C.; Graebin, C.S. Antiprotozoal agents: How have they changed over a decade? Arch. Pharm. 2022, 355, e2100338. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, A.C.; de Souza, M.V.N. Current leishmaniasis drug discovery. RSC Med. Chem. 2022, 13, 1029–1043. [Google Scholar] [CrossRef] [PubMed]
- Askarizadeh, A.; Badiee, A.; Khamesipour, A. Development of nano-carriers for Leishmania vaccine delivery. Expert Opin. Drug Deliv. 2020, 17, 167–187. [Google Scholar] [CrossRef]
- Guerra, R.O.; do Carmo Neto, J.R.; da Silva, P.E.F.; Franco, P.I.R.; Barbosa, R.M.; de Albuquerque Martins, T.; Costa-Madeira, J.; de Assunção, T.S.F.; de Oliveira, C.J.F.; Machado, J.R.; et al. Metallic nanoparticles and treatment of cutaneous leishmaniasis: A systematic review. J. Trace Elem. Med. Biol. 2024, 83, 127404. [Google Scholar] [CrossRef]
- Yadagiri, G.; Singh, A.; Arora, K.; Mudavath, S.L. Immunotherapy and immunochemotherapy in combating visceral leishmaniasis. Front. Med. 2023, 10, 1096458. [Google Scholar] [CrossRef] [PubMed]
- García-Huertas, P.; Olmo, F.; Sánchez-Moreno, M.; Dominguez, J.; Chahboun, R.; Triana-Chávez, O. Activity in vitro and in vivo against Trypanosoma cruzi of a furofuran lignan isolated from Piper jericoense. Exp. Parasitol. 2018, 189, 34–42. [Google Scholar] [CrossRef]
- Zeouk, I.; Sifaoui, I.; López-Arencibia, A.; Reyes-Batlle, M.; Bethencourt-Estrella, C.J.; Bazzocchi, I.L.; Bekhti, K.; Lorenzo-Morales, J.; Jiménez, I.A.; Piñero, J.E. Sesquiterpenoids and flavonoids from Inula viscosa induce programmed cell death in kinetoplastids. Biomed. Pharmacother. 2020, 130, 110518. [Google Scholar] [CrossRef]
- Cartuche, L.; Sifaoui, I.; López-Arencibia, A.; Bethencourt-Estrella, C.J.; San Nicolás-Hernández, D.; Lorenzo-Morales, J.; Piñero, J.E.; Díaz-Marrero, A.R.; Fernández, J.J. Antikinetoplastid Activity of Indolocarbazoles from Streptomyces sanyensis. Biomolecules 2020, 10, 657. [Google Scholar] [CrossRef] [PubMed]
- Núñez, M.J.; Martínez, M.L.; López-Arencibia, A.; Bethencourt-Estrella, C.J.; San Nicolás-Hernández, D.; Jiménez, I.A.; Lorenzo-Morales, J.; Piñero, J.E.; Bazzocchi, I.L. In Vitro Susceptibility of Kinetoplastids to Celastroloids from Maytenus chiapensis. Antimicrob. Agents Chemother. 2021, 65, e02236-20. [Google Scholar] [CrossRef] [PubMed]
- Júnior, J.N.D.S.; da Silva, A.C.; Oliveira, K.K.D.S.; Moreira, L.R.; Caires, S.D.F.F.D.S.; da Silva, A.J.; Moura, Y.A.S.; de Marques, D.A.V.; Bezerra, R.P.; de Lorena, V.M.B.; et al. Green microalgae as a potential source of trypanocide compounds. Nat. Prod. Res. 2023, 1–7. [Google Scholar] [CrossRef]
- Papadopoulou, M.V.; Bloomer, W.D.; Rosenzweig, H.S.; Wilkinson, S.R.; Szular, J.; Kaiser, M. Nitrotriazole-based acetamides and propanamides with broad spectrum antitrypanosomal activity. Eur. J. Med. Chem. 2016, 123, 895–904. [Google Scholar] [CrossRef]
- Nefertiti, A.S.G.; Batista, M.M.; Da Silva, P.B.; Batista, D.G.J.; Da Silva, C.F.; Peres, R.B.; Torres-Santos, E.C.; Cunha-Junior, E.F.; Holt, E.; Boykin, D.W.; et al. In Vitro and In Vivo Studies of the Trypanocidal Effect of Novel Quinolines. Antimicrob. Agents Chemother. 2018, 62, e01936-17. [Google Scholar] [CrossRef]
- Martín-Escolano, R.; Cebrián, R.; Maqueda, M.; Romero, D.; Rosales, M.J.; Sánchez-Moreno, M.; Marín, C. Assessing the effectiveness of AS-48 in experimental mice models of Chagas’ disease. J. Antimicrob. Chemother. 2020, 75, 1537–1545. [Google Scholar] [CrossRef]
- Bethencourt-Estrella, C.J.; Delgado-Hernández, S.; López-Arencibia, A.; San Nicolás-Hernández, D.; Sifaoui, I.; Tejedor, D.; García-Tellado, F.; Lorenzo-Morales, J.; Piñero, J.E. Acrylonitrile Derivatives against Trypanosoma cruzi: In Vitro Activity and Programmed Cell Death Study. Pharmaceuticals 2021, 14, 552. [Google Scholar] [CrossRef] [PubMed]
- Perin, L.R.; Parreira, L.A.; Barcelos, E.C.S.; Santos, M.F.C.; Menini, L.; de Gomes, D.O.; de Careta, F.P. In vitro effect of alpha-bisabolol and its synthetic derivatives on macrophages, promastigotes, and amastigotes of Leishmania amazonensis and Leishmania infantum. Nat. Prod. Res. 2023, 1–6. [Google Scholar] [CrossRef]
- Medicines for Malaria Venture (MMV). Approved/ERP|Medicines for Malaria Venture. Available online: https://www.mmv.org/about-us (accessed on 10 January 2024).
- Medicines for Malaria Venture (MMV). About the Global Health Priority Box|Medicines for Malaria Venture. Available online: https://www.mmv.org/mmv-open/global-health-priority-box/about-global-health-priority-box (accessed on 10 January 2024).
- Rocha, V.P.C.; Nonato, F.R.; Guimarães, E.T.; Rodrigues de Freitas, L.A.; Soares, M.B.P. Activity of antimalarial drugs in vitro and in a murine model of cutaneous leishmaniasis. J. Med. Microbiol. 2013, 62, 1001–1010. [Google Scholar] [CrossRef]
- Heurtault, B.; Legrand, P.; Mosqueira, V.; Devissaguet, J.P.; Barratt, G.; Bories, C. The antileishmanial properties of surface-modified, primaquine-loaded nanocapsules tested against intramacrophagic Leishmania donovani amastigotes in vitro. Ann. Trop. Med. Parasitol. 2001, 95, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.P.; Nascimento, M.S.; Franco, C.H.; Bortoluci, K.; Silva, M.N.; Zingales, B.; Gibaldi, D.; Castaño Barrios, L.; Lannes-Vieira, J.; Cariste, L.M.; et al. Drug Repurposing in Chagas Disease: Chloroquine Potentiates Benznidazole Activity against Trypanosoma cruzi In Vitro and In Vivo. Antimicrob. Agents Chemother. 2022, 66, e0028422. [Google Scholar] [CrossRef]
- Hata, Y.; Zimmermann, S.; Quitschau, M.; Kaiser, M.; Hamburger, M.; Adams, M. Antiplasmodial and antitrypanosomal activity of pyrethrins and pyrethroids. J. Agric. Food Chem. 2011, 59, 9172–9176. [Google Scholar] [CrossRef]
- Menna-Barreto, R.F.S. Cell death pathways in pathogenic trypanosomatids: Lessons of (over)kill. Cell Death Dis. 2019, 10, 93. [Google Scholar] [CrossRef]
- de Menezes, R.R.P.P.B.; Sampaio, T.L.; Lima, D.B.; Sousa, P.L.; de Azevedo, I.E.P.; Magalhães, E.P.; Tessarolo, L.D.; Marinho, M.M.; Dos Santos, R.P.; Martins, A.M.C. Antiparasitic effect of (−)-α-bisabolol against Trypanosoma cruzi Y strain forms. Diagn. Microbiol. Infect. Dis. 2019, 95, 114860. [Google Scholar] [CrossRef] [PubMed]
- de Castro Andreassa, E.; Santos, M.D.M.D.; Wassmandorf, R.; Wippel, H.H.; Carvalho, P.C.; Fischer, J.S.D.G.; Souza, T.A.C.B. Proteomic changes in Trypanosoma cruzi epimastigotes treated with the proapoptotic compound PAC-1. Biochim. Biophys Acta Proteins Proteom. 2021, 1869, 140582. [Google Scholar] [CrossRef]
- Soto-Sánchez, J. Bioactivity of Natural Polyphenols as Antiparasitic Agents and their Biochemical Targets. Mini Rev. Med. Chem. 2022, 22, 2661–2677. [Google Scholar] [CrossRef] [PubMed]
- Swiss Institute of Bioinformatics. SwissADME. Available online: http://www.swissadme.ch/index.php (accessed on 8 March 2024).
- Chang, H.-C.; Huang, Y.-T.; Chen, C.-S.; Chen, Y.-W.; Huang, Y.-T.; Su, J.-C.; Teng, L.-J.; Shiau, C.-W.; Chiu, H.-C. In vitro and in vivo activity of a novel sorafenib derivative SC5005 against MRSA. J. Antimicrob. Chemother. 2016, 71, 449–459. [Google Scholar] [CrossRef]
- Yao, H.; Liu, F.; Chen, J.; Li, Y.; Cui, J.; Qiao, C. Antischistosomal activity of N,N’-arylurea analogs against Schistosoma japonicum. Bioorg. Med. Chem. Lett. 2016, 26, 1386–1390. [Google Scholar] [CrossRef] [PubMed]
- López-Arencibia, A.; San Nicolás-Hernández, D.; Bethencourt-Estrella, C.J.; Sifaoui, I.; Reyes-Batlle, M.; Rodríguez-Expósito, R.L.; Rizo-Liendo, A.; Lorenzo-Morales, J.; Bazzocchi, I.L.; Piñero, J.E.; et al. Withanolides from Withania aristata as Antikinetoplastid Agents through Induction of Programmed Cell Death. Pathogens 2019, 8, 172. [Google Scholar] [CrossRef]
- San Nicolás-Hernández, D.; Hernández-Álvarez, E.; Bethencourt-Estrella, C.J.; López-Arencibia, A.; Sifaoui, I.; Bazzocchi, I.L.; Lorenzo-Morales, J.; Jiménez, I.A.; Piñero, J.E. Multi-target withaferin-A analogues as promising anti-kinetoplastid agents through the programmed cell death. Biomed. Pharmacother. 2023, 164, 114879. [Google Scholar] [CrossRef]
- Bethencourt-Estrella, C.J.; Delgado-Hernández, S.; López-Arencibia, A.; San Nicolás-Hernández, D.; Tejedor, D.; García-Tellado, F.; Lorenzo-Morales, J.; Piñero, J.E. In vitro activity and mechanism of cell death induction of cyanomethyl vinyl ethers derivatives against Trypanosoma cruzi. Int. J. Parasitol. Drugs Drug Resist. 2023, 22, 72–80. [Google Scholar] [CrossRef] [PubMed]
- San Nicolás-Hernández, D.; Bethencourt-Estrella, C.J.; López-Arencibia, A.; Hernández-Álvarez, E.; Sifaoui, I.; Bazzocchi, I.L.; Lorenzo-Morales, J.; Jiménez, I.A.; Piñero, J.E. Withaferin A-silyl ether analogs as potential anti-kinetoplastid agents targeting the programmed cell death. Biomed. Pharmacother. 2023, 157, 114012. [Google Scholar] [CrossRef] [PubMed]
- Bethencourt-Estrella, C.J.; Delgado-Hernández, S.; López-Arencibia, A.; San Nicolás-Hernández, D.; Tejedor, D.; García-Tellado, F.; Lorenzo-Morales, J.; Piñero, J.E. In vitro activity and cell death mechanism induced by acrylonitrile derivatives against Leishmania amazonensis. Bioorg. Chem. 2022, 124, 105872. [Google Scholar] [CrossRef] [PubMed]
- Bethencourt-Estrella, C.J.; Nocchi, N.; López-Arencibia, A.; San Nicolás-Hernández, D.; Souto, M.L.; Suárez-Gómez, B.; Díaz-Marrero, A.R.; Fernández, J.J.; Lorenzo-Morales, J.; Piñero, J.E. Antikinetoplastid Activity of Sesquiterpenes Isolated from the Zoanthid Palythoa aff. clavata. Pharmaceuticals 2021, 14, 1095. [Google Scholar] [CrossRef]
- García-Davis, S.; López-Arencibia, A.; Bethencourt-Estrella, C.J.; San Nicolás-Hernández, D.; Viveros-Valdez, E.; Díaz-Marrero, A.R.; Fernández, J.J.; Lorenzo-Morales, J.; Piñero, J.E. Laurequinone, a Lead Compound against Leishmania. Mar. Drugs 2023, 21, 333. [Google Scholar] [CrossRef]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CC50 Murine Macrophages (µM) | IC50 Leishmania amazonensis (µM) | SI | ||
|---|---|---|---|---|
| Flucofuron | 83.86 ± 20.76 | Promastigote | 6.07 ± 1.11 | 13.8 |
| Amastigote | 3.14 ± 0.39 | 26.7 | ||
| Miltefosine | 72.18 ± 8.85 | Promastigote | 6.48 ± 0.25 | 11.1 |
| Amastigote | 3.12 ± 0.30 | 23.1 | ||
| CC50 Murine Macrophages (µM) | IC50 Trypanosoma cruzi (µM) | SI | ||
| Flucofuron | 83.86 ± 20.76 | Epimastigote | 4.28 ± 0.83 | 19.6 |
| Amastigote | 3.26 ± 0.34 | 25.7 | ||
| Benznidazole | 399.91 ± 1.40 | Epimastigote | 6.92 ± 0.77 | 57.8 |
| Amastigote | 2.67 ± 0.39 | 149.8 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bethencourt-Estrella, C.J.; López-Arencibia, A.; Lorenzo-Morales, J.; Piñero, J.E. Global Health Priority Box: Discovering Flucofuron as a Promising Antikinetoplastid Compound. Pharmaceuticals 2024, 17, 554. https://doi.org/10.3390/ph17050554
Bethencourt-Estrella CJ, López-Arencibia A, Lorenzo-Morales J, Piñero JE. Global Health Priority Box: Discovering Flucofuron as a Promising Antikinetoplastid Compound. Pharmaceuticals. 2024; 17(5):554. https://doi.org/10.3390/ph17050554
Chicago/Turabian StyleBethencourt-Estrella, Carlos J., Atteneri López-Arencibia, Jacob Lorenzo-Morales, and José E. Piñero. 2024. "Global Health Priority Box: Discovering Flucofuron as a Promising Antikinetoplastid Compound" Pharmaceuticals 17, no. 5: 554. https://doi.org/10.3390/ph17050554