2.2.1. Azadirachta indica A. Juss. (Indian Lilac, Hawaiian Name: Neem)
Azadirachta indica belonging to the Meliaceae family, is known as, or commonly referred to as neem, nim tree, or Indian lilac. It has been widely distributed in tropical and subtropical regions. Anxiety may occur as a symptom of clinical major depression. In 2019, Falaki et al. examined the anxiolytic effects of the leaf extract of
Tapinanthus globiferus grown on
A. indica and found that it had a significant anxiolytic activity. This finding supported the traditional use of this plant for anxiety relief and was consistent with the outcomes of previous studies that showed antidepressant activity in methanol extracts of the same plant and anxiolytic and antidepressant activity in a crude aqueous extract from the stem bark of another specie,
T. dodoneifolius [
31].
In 2022, Hawiset et al. investigated the potential anxiolytic and antidepressant-like effects of an aqueous extract obtained from the flowers of A. indica in rats subjected to stress. Male Wistar rats were divided randomly into two main groups: a control group and a stress-induced group. Rats in the stress group were exposed to a 3-h restraint stress. Over a duration of 30 days, stressed rats were administered different substances: a control solution, diazepam, fluoxetine, and A. indica extract at doses of 250, 500, and 1000 mg/kg body weight. To assess anxiolytic and antidepressant-like behaviors, the researchers employed three behavioral tests: the elevated plus-maze test (EPMT), the forced swimming test (FST), and the open field test (OFT). The EPMT involved measuring the percentage of open-arm entries and the time spent in open arms. Notably, stressed rats treated with diazepam or A. indica flower extract at a dosage of 500 mg/kg BW exhibited significant improvements in these measurements. Moreover, rats exposed to stress and subsequently administered fluoxetine or A. indica flower extract at all tested doses displayed a noteworthy reduction in immobility time during the FST, indicating potential antidepressant-like effects. Notably, no notable variance in spontaneous locomotor activity was observed across any of the experimental groups.
Further analysis revealed that rats subjected to stress and treated with either standard positive control drugs or
A. indica flower extract showed elevated levels of brain dopamine (DA) and serotonin (5-HT), accompanied by reduced blood cortisol levels compared to stressed rats given the control solution. Additionally, administration of
A. indica flower extract did not result in any harmful effects on the rats’ liver tissue, as determined through examination. This study highlighted that
A. indica flower extract, particularly at a dose of 500 mg/kg BW, could potentially have anxiolytic and antidepressant-like effects in stressed rats, as demonstrated through behavioral tests and biochemical analyses of neurotransmitter levels. Furthermore, the extract showed no detrimental impact on the rats’ liver tissue. Administration of the extract could attenuate the stress-induced behavioral impairment by regulating dopaminergic and serotonergic functions in the rats, indicating potential anxiolytic and antidepressant applications of the extract [
32].
2.2.3. Curcuma longa L. (Turmeric, Indian Saffron, Hawaiian Name: Ōlena)
In a 2020 review, Ramaholimihaso et al. reported that curcumin (
Figure 2) from
C. longa has been found to possess anti-inflammatory and antioxidant properties that may provide therapeutic benefits in the treatment of depression. A patient with depression often experiences a lot of anxiety. Curcumin has demonstrated promising effects in reversing depressive-like behaviors induced by chronic stress in mice. It appears to enhance serotoninergic and dopaminergic transmission while inhibiting the activity of the monoamine oxidase (MAO-A) enzyme. Observation of similar outcomes in rats found that curcumin increased serotonin and dopamine levels in a dose-dependent manner and inhibited monoamine oxidase enzymes. Recent investigations also revealed that curcumin elevated norepinephrine, serotonin, and dopamine levels in the frontal cortex, hippocampus, and striatum of rats. In a study of curcumin’s effects on serotonin (5-HT) receptors, an antidepressant-like effect was linked to interactions with serotonin receptors 5-HT1A/1B and 5-HT2C. Correspondingly, the antidepressant effect of curcumin was associated with increased expression of 5-HT1A receptors, supported by elevated 5-HT1A receptor mRNA levels across various hippocampal subfields after curcumin administration. These findings noted increased 5-HT1A receptor expression following curcumin administration in chronically stressed mice. Notably, the antidepressant-like effect was countered when a 5-HT1B receptor antagonist was administered. Collectively, these animal studies provide robust evidence that curcumin has the potential to modulate monoaminergic systems in preclinical rodent models, offering insights into its antidepressant properties. It has been suggested that curcumin may be used as a prophylactic agent due to its neuroprotective effects against stress-induced toxicity. Nevertheless, more clinical trials are needed to determine its efficacy and optimal dosage [
35].
In addition, in 2019, Bhat et al. reviewed that curcumin (
Figure 2) has demonstrated potent antidepressant effects in animal models of depression. Its mechanism involves the inhibition of monoamine oxidase (MAO-A and MAO-B) enzymes, leading to elevated levels of norepinephrine, serotonin, and dopamine. By modulating extracellular regulated kinase (ERK) activity, curcumin enhances the expression of brain-derived neurotrophic factor (BDNF) in the amygdala of mice, contributing to its antidepressant actions. Additionally, curcumin has been found to stimulate hippocampal neurogenesis and elevate BDNF levels in a mouse model of chronic stress. Its anxiolytic-like effects are thought to stem from the reduction in pro-inflammatory mediators like iNOS and COX-2 mRNA, achieved through the NF-κB signaling pathway. The inhibition of proinflammatory cytokine IL-1β through the NF-κB pathway further confirms curcumin’s anti-inflammatory potential in the context of depression. Thus, curcumin promoted hippocampal neurogenesis, improved brain-derived neurotrophic factor (BDNF) levels in a mouse model of chronic stress and exerted anxiolytic effects via decreased levels of pro-inflammatory mediators and inhibition of proinflammatory cytokines IL-1β via NF-κB pathway [
36].
A random effects meta-analysis involving five studies with a total of 284 participants (160 in the curcumin group and 124 in the comparison group) revealed a significant and substantial positive effect of curcumin on anxiety symptoms. The effect size was large, indicated by Hedge’s g value of −2.62, with a 95% confidence interval ranging from −4.06 to −1.17 (p < 0.001). The level of variability among the studies (heterogeneity) was low, with an I² value of 23.89%. Further analysis, by systematically excluding each individual study from the overall effect size, did not yield any statistically significant differences in the results. This finding contrasts with a recent rat model study, which reported no anxiolytic effects or changes in behavioral despair associated with curcumin. Additionally, this earlier research did not identify any interactions between curcumin and the benzodiazepine site of the γ-aminobutyric acid (GABA)-A receptor.
Nonetheless, various potential mechanisms could explain the observed anxiety-reducing effects of curcumin. For instance, curcumin may enhance serotoninergic transmission in rats by inhibiting monoamine oxidase (MAO), leading to increased serotonin levels in the brain’s medial prefrontal cortex. Other studies suggested that curcumin promotes the conversion of hepatic α-linoleic acid into docosahexaenoic acid (DHA), an omega-3 fatty acid with anxiolytic-like properties and enhances its accumulation in the brain. Additionally, curcumin has been found to suppress the synthesis of a specific form of nitric oxide synthase (iNOS), which tends to increase during stress in the brain cortex, thereby producing effects akin to anxiolysis. It is worth noting that these mechanistic insights are derived solely from preclinical studies, and as of now, no dedicated trials assessing the efficacy of curcumin for anxiety disorders in humans have been conducted, as suggested by Fusar-Poli et al. [
37].
In 2021, Moragrega et al. described that curcumin, an active compound found in
C. longa, exhibits antidepressant effects through intricate mechanisms involving both the serotonergic system and the AC-cAMP pathway. Additionally, curcumin enhances the antidepressant actions of subliminal doses of drugs like fluoxetine, venlafaxine, and bupropion. When combined with piperine, a bioavailability enhancer, curcumin’s impact is further amplified. This combination not only boosts the neurotransmitter serotonin (5-HT) systems mediate dopamine (DA) but also intensifies the inhibitory effects on MAO-A, surpassing the effects of curcumin administered alone. In studies conducted on rats subjected to bilateral olfactory bulbectomy, a procedure that induces depression-like behavioral changes, curcumin effectively counteracts these alterations, similar to conventional antidepressants. This correction is attributed to curcumin’s ability to mitigate immobility time and reverse the behavioral anomalies triggered by bulbectomy. Moreover, curcumin demonstrates a protective effect on rat hippocampal neurons against damage caused by chronic stress. This safeguarding mechanism involves the positive regulation of receptors like serotonin 1A (5-HT1A) and brain-derived neurotrophic factor (BDNF) expression, both crucial for hippocampal neurogenesis. Exploration of curcumin’s anxiolytic and antidepressant-like properties through various selective tests. Their findings dismiss the idea that curcumin’s effects are tied to the benzodiazepine site on the GABAA receptor. Instead, these effects likely arise from interactions with other receptor subunits or central nervous system neurotransmitter systems, corroborating the previously mentioned mechanisms underlying curcumin’s antidepressant actions. This study examined the anxiolytic and antidepressant-like effects of curcumin in various tests and demonstrated that its effects were not likely due to interaction with the GABAa receptor benzodiazepine site, suggesting that curcumin may modulate other receptor subunits or interact with other central nervous system neurotransmitter systems [
38].
In 2021, Latif et al. conducted a study to assess the impact of consuming natural turmeric on cardiovascular risk factors, mental well-being, and serum homocysteine levels in overweight and obese women. The study utilized a pre-post, single-arm design carried out at the Department of Physiology in Imam Abdulrahman Bin Faisal University, located in Dammam, Saudi Arabia. The study included 36 female university students, all of whom had a body mass index (BMI) of 23 kg/m² or higher. Over a span of 90 days, participants were provided with a daily dosage of 2 g of C. longa in the form of capsules. The study collected initial and post-intervention data on various aspects, including anthropometric measurements, blood pressure, serum homocysteine levels, and indicators of mental health such as stress, anxiety, and depression scores.
Additionally, the potential confounding variables of dietary intake and physical activity were also assessed. The results indicated noteworthy reductions in multiple anthropometric measurements after the intervention, encompassing body weight, body mass index, waist circumference, hip circumference, body fat percentage, and systolic blood pressure. There was also a significant decrease in anxiety scores. However, there were no significant alterations observed in homocysteine levels, stress, or depression scores. Dietary intake and physical activity remained relatively constant during the study duration. Thus, this study demonstrated that daily consumption of 2 g of C. longa over 90 days could potentially lead to weight loss, lowered body fat percentage, reduced systolic blood pressure, and decreased anxiety levels in young, overweight and obese women.
Nonetheless, there were no significant changes in homocysteine levels, stress, or depression scores throughout the study period. The effects of
C. longa supplementation (2 g/d for 90 days) on obesity-related and cardiovascular-disease risk factors in overweight or obese females. Results showed that
C. longa was able to reduce weight, body fat percentage, systolic blood pressure, and anxiety levels in the participants. These findings suggested that
C. longa may be a promising treatment for obesity-related and cardiovascular disease risk factors besides anxiety in this population [
39].
Current evidence suggests that curcumin (
Figure 2) may have antidepressant properties, as evidenced by its ability to improve depressive and anxiety behavior, increase monoamines and brain-derived neurotrophic factor levels, inhibit pro-inflammatory cytokines, and reduce neuronal apoptosis. Additionally, curcumin has been reported to improve insulin sensitivity, reduce cortisol levels, and reverse metabolic abnormalities. Further research is needed to evaluate the efficacy of curcumin in depression treatment, as the dose and formulation of curcumin vary, as described and reviewed by Matias et al. in 2021 [
40]. Later, Lopresti et al. studied the impact of a curcumin extract called Curcugen™ on gastrointestinal symptoms, mood, and overall quality of life in adults who reported digestive issues. Additionally, the study aimed to uncover potential mechanisms of curcumin’s effects by investigating its impact on gut microbiota and small intestinal bacterial overgrowth (SIBO). The research was conducted over 8 weeks, employing a double-blind, randomized controlled trial involving 79 adults with self-reported digestive complaints. Participants were randomly assigned to receive either a placebo or 500 mg of the curcumin extract, Curcugen™. Multiple measurements were taken, including the Gastrointestinal Symptom Rating Scale (GSRS), Depression, Anxiety, and Stress Scale-21 (DASS-21), Short Form-36 (SF-36), as well as a SIBO breath test. The study also analyzed the participants’ intestinal microbial profiles using 16S rRNA sequencing. The results, derived from self-reported data of 77 participants, demonstrated that those who received curcumin experienced a notable reduction in the total GSRS score, indicating an improvement in gastrointestinal symptoms when compared to the placebo group. Furthermore, there was a significant decrease in the anxiety score measured by DASS-21 in the curcumin group. However, no other substantial differences were observed in self-reported data between the two groups. Notably, curcumin did not seem to have a significant impact on intestinal microbial composition or SIBO test results. Importantly, curcumin was well-tolerated without any significant adverse events. To conclude, administering the Curcugen™ curcumin extract at a dosage of 500 mg once daily over an 8-week period led to greater enhancements in both digestive complaints and anxiety levels among adults reporting such issues. While the study did not identify significant changes in gut microbiota or SIBO when compared to the placebo, further research using larger participant samples and more comprehensive microbial analysis methods will be crucial. Exploring additional potential mechanisms underlying curcumin’s beneficial effects on gastrointestinal symptoms, such as its influence on factors like intestinal barrier function, inflammation, neurotransmitter activity, and visceral sensitivity, also warrants investigation. Further research with larger sample sizes and methods that allow for more detailed microbial analyses is needed to further explore the potential benefits of curcumin on gastrointestinal health and anxiety. Additionally, future studies should focus on other potential mechanisms associated with curcumin’s gastrointestinal-relieving effects, such as its influence on intestinal barrier function, inflammation, neurotransmitter activity, and visceral sensitivity [
41].
Norwitz et al. reviewed that curcumin has been investigated as a possible remedy for various brain-related conditions such as Alzheimer’s disease, Parkinson’s disease, depression, anxiety, and related conditions. The mechanisms of curcumin modulation of neurotransmitters like dopamine, serotonin, and cortisol, and regulation of microRNAs and histone deacetylases (HDACs). Preclinical assessments of curcumin’s impact on anxiety in animal models have reinforced its potential as an anti-anxiety agent. In studies involving rodents exposed to anxiety-inducing factors, curcumin intervention effectively mitigated anxiety-related behaviors. These findings showed that curcumin not only significantly reduced anxiety-like behaviors but also positively influenced neurotransmitter and hormone levels. Several rigorously conducted trials involving human participants, characterized by randomization, double-blinding, and placebo controls, have substantiated the anxiety-reducing effects of curcumin supplementation. For instance, in patients with diabetes, an eight-week curcumin supplementation regimen led to decreased anxiety. Likewise, a crossover trial involving obese individuals demonstrated that curcumin supplementation over a 30-day period resulted in lowered anxiety scores. A meta-analysis encompassing five studies further revealed an overall significant reduction in anxiety due to curcumin supplementation, with a substantial effect size (Hedge’s g = −2.62). However, it is important to acknowledge the limitations inherent in curcumin research. Critics have raised concerns about the potential overhyping of the health advantages associated with turmeric and its active constituents. A comprehensive analysis highlighted the chemical instability of curcumin and its potential to interfere with assay results, suggesting that positive findings in experimental models might be skewed.
Furthermore, variations in supplement purity and formulations among different studies have hindered reproducibility. Curcuminoids, being fat-soluble, exhibit limited bioavailability when administered alone or in aqueous solutions. To enhance bioavailability, curcuminoids should be consumed with fats, and approaches such as liposomes and nanoparticles are being explored for their administration. Notably, the positive curcumin-anxiety studies mentioned earlier utilized techniques to boost curcumin bioavailability, including nano-curcumin and co-administration of bioperine, which amplifies curcumin absorption by 20-fold. As such, future research efforts should concentrate on investigating these more bioavailable forms of curcumin, as well as examining curcumin’s impact on the human microbiome, a pathway not reliant on systemic absorption. The connection between
C. longa and the previous section on omega-3 fatty acids is noteworthy. Humans struggle to efficiently convert alpha-linolenic acid (ALA) into eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA). Curcumin could enhance this conversion by elevating levels of the enzymes responsible for DHA synthesis. This not only heightens DHA levels in the brain but also holds implications for anxiety regulation. Rodents treated with a combination of ALA and curcumin displayed reduced anxiety [
42].
In 2022, Lopresti et al. suggested that disturbances in the functioning of neurotransmitters like serotonin (5-HT), dopamine, noradrenaline, and glutamate have consistently been observed in cases of depression. Animal trials have shown that curcumin, a compound found in
C. longa, can influence the levels and activity of these neurotransmitters. For instance, when administered to mice, curcumin quickly improved depressive behaviors by positively affecting the 5-HT1A/2A receptor. In mice that had their ovaries removed, curcumin changed depressive behaviors and increased serotonin levels in various brain regions by promoting the expression of key genes involved in serotonin regulation while reducing the activity of a specific enzyme called monoamine oxidase A. In rats subjected to chronic stress, curcumin lessened anxious behaviors and reversed stress-related reductions in serotonin levels in brain areas such as the hippocampus, amygdala, and striatum. Additionally, curcumin had effects on dopamine and noradrenaline levels in different brain regions in rats that had their ovaries removed, counteracted diabetes-induced changes in dopamine receptors, and mitigated dopamine depletion triggered by a pesticide called rotenone. Furthermore, curcumin exhibited the ability to protect against damage caused by excess glutamate, a neurotransmitter linked to neurotoxicity. It achieved this by influencing the activity of
N-methyl-
D-aspartate (NMDA) receptors, which are important for glutamate signaling. Curcumin boosted the expression of a specific subunit of NMDA receptors known as GluN2A and encouraged the activation of a specific protein in α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors, termed GluR1. These actions collectively indicate that curcumin has the potential to impact various neurotransmitter systems and pathways, offering promise as a potential therapeutic agent for depression and related conditions. There was evidence supporting the efficacy of curcumin as a treatment for depression, as demonstrated by animal and human trials and confirmed by multiple meta-analyses. However, further research is necessary to assess the safety, tolerability, and effectiveness of different curcumin extracts, given their varying levels of oral bioavailability [
43].
2.2.4. Morinda citrifolia L. (Noni–Indian Mulberry, Great Morinda, Cheese Fruit)
Morinda citrifolia (noni) is a perennial, fruit-bearing tree belonging to the Rubiaceae family, found in Southeast Asia, consumed as both food and medicine for over 2000 years [
44,
45,
46]. In 2019, West et al. reported the HPLC analysis and the effects of
M. citrifolia fruit puree juice on the two major endocannabinoid degradation enzymes, fatty acid amide hydrolase (FAAH) and monoacylglycerol lipase (MAGL). HPLC analysis unveiled the presence of phytochemical components, including deacetylasperulosidic acid (DAA), asperulosidic acid (AA), scopoletin, rutin, and quercetin (
Figure 7) within
M. citrifolia juice samples. Disruption of fatty acid amide hydrolase (FAAH) activity prevents the effects of chronic stress on anxiety. Deacetylasperulosidic acid (DAA) displayed a moderate inhibition of FAAH activity, reducing it by approximately 28.26% with a standard deviation of 4.61%. It is possible that additional compounds present in
M. citrifolia juice could have also played a role in influencing the observed activity in our experimental tests. An experiment involving
M. citrifolia juice and its effects on monoacylglycerol lipase (MAGL) activity in a concentration-dependent manner. MAGL is an enzyme responsible for breaking down 2-arachidonoylglycerol (2-AG). The results revealed that as the concentration of noni juice increased, the activity of MAGL decreased, at the lowest concentration of 2.78 μL/mL of
M. citrifolia juice, was reduced by approximately 32.88% ± 3.62, at a concentration of 5.56 μL/mL of
M. citrifolia juice, MAGL activity was reduced by approximately 55.71% ± 4.40%, at the highest concentration of
M. citrifolia juice tested was 18.52 μL/mL, MAGL activity was completely prevented. Hence, these compounds from
M. citrifolia juice may contribute to the FAAH and MAGL inhibitory activity, potentially supporting mental health, joint health, relieving discomfort, and modulating the immune system [
45].
A pilot clinical study involving Tahitian
M. citrifolia juice also reported improved mental health scores. The underlying biological mechanism for these effects is related to Gamma-aminobutyric acid (GABA), a vital inhibitory neurotransmitter in the central nervous system. GABA binds to specific receptors on both pre- and post-synaptic neurons in the plasma membrane. Among potential targets for various central nervous system disorders, the GABA receptor stands out, particularly in anxiety-related conditions. Extensive clinical experience and well-designed experiments have established the significance of GABA in mediating different anxiety disorders. Furthermore, neurotransmitters like norepinephrine and serotonin also play crucial roles in these processes. Alterations in these neurotransmitters have been observed in association with
M. citrifolia treatment. Medications that modulate the GABAergic or serotonergic systems or that reduce adrenergic activity are commonly used for treating anxiety disorders. Recent research conducted on rats demonstrated a significant reduction in norepinephrine levels in the amygdala and hippocampus, further supporting the potential benefits of
M. citrifolia in anxiety management, as reviewed by Ramya et al. in 2021 [
46]. In addition, epidemiological studies showed that anxiety disorders are more common among people diagnosed with psychotic disorders compared to the general population.
M. citrifolia fruits have demonstrated neuropharmacological activities, such as anti-depressant and anti-anxiety effects, in both in vivo and in vitro models. The research aimed to investigate the potential antipsychotic effect of
M. citrifolia fruits using mouse models of climbing activity. The findings of this study revealed an antidopaminergic effect of
M. citrifolia in mice, suggesting that
M. citrifolia may exhibit antipsychotic-like activity and could be a viable candidate for the treatment of mental conditions. Furthermore, to evaluate the potential antipsychotic activity of
M. citrifolia juice, the researchers administered various doses (5, 10, and 100 mL/kg) equivalent to dried
M. citrifolia juice powder doses (450, 900, and 1800 mg/kg) to mice. The results indicated that
M. citrifolia juice significantly reduced amphetamine-induced climbing behavior and methamphetamine-induced stereotypy at these different doses. The
M. citrifolia extract was orally given in combination with apomorphine and methamphetamine over a period of 21 days, and it demonstrated antipsychotic activity. This study adds to the understanding of the neurochemical basis of schizophrenia and highlights the potential antipsychotic properties of
M. citrifolia extract. However, further research is warranted to fully explore its mechanism of action and its efficacy compared to standard antipsychotic drugs for human clinical use [
46].
In various in vivo studies, a considerable body of research has indicated that
M. citrifolia holds promise for potential pharmacological applications in treating depression and anxiety. A study delved into the possible role of serotonergic and noradrenergic pathways in the antidepressant effects of
M. citrifolia. This was explored through interactions with substances like a depletory of adrenergic and dopaminergic compounds (AMPT), a serotonergic depletor (PCPA), and a 5HT1A receptor antagonist in mice. The study investigated the influence of the serotonergic system on the antidepressant-like activity of noni in the tail suspension test (TST), a well-accepted animal model for depression research due to its reliability and sensitivity. In the TST mouse model, the administration of noni extract at doses of 0.5 and 0.75 g/kg significantly reduced the duration of immobility, akin to the effect seen after intraperitoneal administration of the standard antidepressant desipramine (30 mg/kg). The activity of biogenic and dietary amines, regulated by key isoenzymes like monoamine oxidase (MAO-A and MAO-B), plays a crucial role. While MAO-A predominantly oxidizes serotonin (5-HT) and noradrenaline (NA), MAO-B primarily oxidizes phenyl ethylamine (PEA), and both can influence dopamine levels. Tranylcypromine, a non-selective and irreversible inhibitor of both MAO-A and MAO-B, has shown effectiveness in treating severe depression.
M. citrifolia extract, like tranylcypromine, demonstrated the ability to inhibit both MAO-A and MAO-B enzymes, thus potentially contributing to its antidepressant-like effects in mice and subsequently increasing the levels of biogenic amines like 5-HT and NA. Earlier studies indicated that
M. citrifolia’s antidepressant-like effects are mediated through its interaction with serotonergic and noradrenergic systems. Building upon this, in vitro investigations were conducted to study the inhibitory effects of noni fruit extract and its identified components on MAO-A and MAO-B enzymes, suggesting their therapeutic potential for antidepressant activity. Additionally, ex vivo studies have revealed that noni fruit contains various chemical constituents such as rutin, scopoletin, and coumarin derivatives (
Figure 7). Rutin, with its pharmacological properties encompassing antidepressant, antioxidant, neuroprotective, and antianxiety effects, has shown promise. Moreover, compounds like scopoletin and rutin have demonstrated sequential antidepressant activity, potentially interacting with α1- and α2-adrenoreceptors [
47].
The study investigated the anxiolytic and antidepressant-like properties of M. citrifolia’s potential mechanisms of action. The effects of M. citrifolia were examined through drug interaction studies involving selective benzodiazepine receptor antagonist (flumazenil), GABAA receptor antagonist (bicuculine), and 5HT1A receptor antagonist to understand its interaction with neurotransmitter systems associated with anxiety. The findings indicated that M. citrifolia extract (1 g/kg, p.o.) exhibited anxiolytic effects in mouse models, similar to the well-known anxiolytic drug diazepam (1 mg/kg, i.p.). This was observed in various tests, such as the elevated plus maze (EPM) and light–dark transition (LDT) tests. Notably, M. citrifolia increased the time spent in the open arms of the EPM, similar to the effects of diazepam. Previous in vitro studies suggested that the methanolic extract of M. citrifolia fruit binds preferentially to GABAA receptors, which play a critical role in controlling anxiety. Benzodiazepine-GABAergic neurotransmission was highlighted as vital in anxiety control. Further studies involving noni juice administration to rats for 15 days showed increased exploration of uncomfortable open arms in the EPM. This behavior, akin to benzodiazepine treatment, suggested potential anxiolytic effects. Interestingly, motor activity and rearing behavior remained consistent between treatments. In vitro studies demonstrated that the methanolic extract of noni fruit exhibited 75% binding suppression to GABAA receptors, indicating anxiolytic activity. This extract inhibited the binding of the agonist radioligand (3H) muscimol to GABAA receptors at a concentration of 100 mg/mL, thereby highlighting its GABAergic activity. This receptor binding plays a role in regulating the inhibitory neurotransmitter GABA, contributing to anxiolytic and sedative effects. Neurochemical evaluations indicated modifications in the monoaminergic system in M. citrifolia-treated rats compared to controls. The changes included alterations in neurotransmitters like norepinephrine (NE), serotonin (5-HT), 3,4-dihydroxyphenylacetic acid (DOPAC), and homovanillic acid (HVA). M. citrifolia’s potential to lower these neurotransmitter levels, particularly 5-HT, suggests anxiolytic properties. The effects on norepinephrine (NE), dopamine (DA), and its intermediates point to reduced anxiety levels. The modulation of different neurotransmitters by M. citrifolia appears to be mediated through the GABAergic system. M. citrifolia’s GABAA receptor binding affinity further supports its potential to regulate monoaminergic neurons in various brain regions. In vitro findings from an elevated plus maze test highlighted noni juice’s anxiolytic effect mediated in part by alterations in monoaminergic neurotransmitter levels.
Consequently, recent research on
M. citrifolia suggests potential anxiolytic and antidepressant-like effects. These effects are likely mediated through interactions with the GABAergic and serotonergic systems.
M. citrifolia’s ability to regulate neurotransmitter levels and its GABAA receptor binding affinity underpins its potential therapeutic applications for anxiety and depression. These findings suggest the potential therapeutic applications of
M. citrifolia in various neurological and psychiatric conditions, including anxiety, as summarized by Begum et al. in 2022 [
47].
2.2.5. Passiflora edulis Sims (Passionfruit, Hawaiian Name: Lilikoi)
Passiflora edulis (Passifloraceae), commonly known as passion fruit, is popular due to its balanced nutrition, juiciness, and attractive nutritional value. It is widely consumed for its essential benefits for health and is popular among consumers [
48,
49].
In 2019, Gross et al. summarized that
P. edulis has been traditionally utilized in Mexico for addressing conditions such as nervousness, anxiety, and depression. Extracts derived from its leaves through percolation have demonstrated a depressant impact on the central nervous system in mice. Both hydroalcoholic and aqueous leaf extracts have exhibited anxiolytic effects in rat experiments conducted using the high cross labyrinth.
P. edulis is recognized for its potential as an anxiolytic and mild sedative. However, it’s important to note that its consumption might lead to drowsiness, and caution is advised against chronic usage or simultaneous administration with other nervous system sedatives and depressants [
50].
In 2020, He et al. demonstrated the therapeutic effects of
P. edulis, with the crude extracts (butanol, methanol, ethanol, hydroethanolic, and aqueous extract) exhibiting anxiolytic-like effects in rat models. When administered orally, the aqueous extract of
P. edulis (at 50, 100, and 150 mg/kg) showed anxiolytic-like effects in the elevated plus-maze and inhibitory avoidance tests without impairing memory processes. Of greater significance, administration of the aqueous extract of
P. edulis did not disturb the memory function of rats during their adaptation to an open-field test. Conversely, the habituation of rats was hindered by diazepam, albeit through a basic alteration of the open-field apparatus. The methanol extract derived from the aerial parts of
P. edulis demonstrated anxiolytic effects in mice when administered orally at a dose of 75 mg/kg, as evidenced by its performance in the elevated plus-maze anxiety model. However, an oral dose of 125 mg/kg did not yield a significant anxiolytic response. Alternatively, higher oral doses of 200 and 300 mg/kg resulted in a mild sedative impact. In a similar vein, prior administration of hydroethanolic extracts at 50, 100, and 150 mg/kg, as well as spray-dried leaf powders at 400 and 800 mg/kg of
P. edulis, displayed anxiolytic properties in the elevated plus-maze test involving mice. These effects were attributed to the presence of a diverse array of flavonoids and their glycosides, underscoring the potential therapeutic value of these extracts [
48].
Evaluate the protective effects of hydroalcoholic extract derived from
P. edulis leaves against anxiety induced by sub-acute immobilization stress in mice. The anxiety induction was achieved through sub-acute immobilization stress, followed by an 11-day treatment period. Subsequently, behavioral assessments were conducted using the elevated plus maze (EPM) and open field (OF) tests, followed by the analysis of biochemical parameters, including malondialdehyde (MDA), glutathione (GSH), superoxidedismutase (SOD), catalase (CAT), GABA, GABA-T, and serotonin (5-HT). The findings demonstrate that the administration of
P. edulis extract at doses of 100 and 200 mg/kg resulted in a significant increase in the number of entries and time spent in the open arms of the EPM, while decreasing entries and time in the closed arms. In the OF test, the extract led to a significant increase in measures such as lines crossed, passages through the center, and center time. These outcomes suggest that
P. edulis extract potentially offers protection against anxiety. This effect could be attributed to its ability to combat oxidative stress and counteract hyperexcitability by enhancing GABA’s action. The most effective dose, 100 mg/kg, notably increased GSH activity to 4.44 ± 0.24 µmol/g. In mice treated with this dose, the extract significantly reduced three oxidative stress markers: MDA, catalase, and SOD levels were lowered to 0.22 ± 0.01 µmol/g, 1.05 ± 0.15 mmol H
2O
2/min/g, and 19.46 ± 0.00 unit/min/mg, respectively, compared to the negative control. Moreover, animals treated with 100 mg/kg of P. edulis extract exhibited a substantial increase (
p < 0.001) in GABA and 5-HT levels, reaching 4.62 ± 0.28 µg/g and 31.47 ± 1.58 ng/mL, respectively. The activity of GABA-T was also influenced by the treatment with P. edulis; the GABA-T activity, which was 1.27 ± 0.10 in the negative control, significantly (
p < 0.001) decreased to 0.37 ± 0.00 in the group treated with the 100 mg/kg dose. Ultimately, the findings of the study revealed the beneficial effects of
P. edulis extract in mitigating anxiety-like behavior in mice. These effects were linked to the extract’s ability to counteract oxidative stress and enhance GABAergic neurotransmission, thereby suggesting its potential as a natural remedy for anxiety-related conditions, as mentioned by Espoir et al. in 2020 [
51].
Additionally, the anxiolytic-like effect of
P. edulis extract was tested in a randomized trial using an anxiety model in rats. The elevated plus-maze (EPM) test, incorporating Diazepam as a positive control, demonstrated that diazepam produced an anxiety-reducing effect in mice. This effect was characterized by a notable increase in the percentage of open-arm entries (OE) and total time spent in the open arms (OT) of the EPM without altering the number of entries into enclosed arms. This outcome was consistent with prior research. In a similar manner, treatment with an ethanol extract (EE) derived from
P. edulis at doses of 200 and 400 mg/kg resulted in a significant rise in the percentage of open-arm entries and total time spent in the open arms, indicating an anxiety-reducing effect. Interestingly, the group treated with 400 mg/kg of EE showed a decrease in the number of entries into enclosed arms, implying a reduction in motor activity, suggesting that at this higher dose, the EE had more of a sedative-like effect. Additionally, administration of various fractions of
P. edulis extract, such as chloroform (CF), ethylacetate (EAE), and butanol (BF), at a dose of 200 mg/kg, also led to a noteworthy increase in open arm entries and total time spent in open arms, indicating an anxiolytic effect. However, no significant distinctions were observed among these treatment groups. Notably, BF exhibited the highest activity, followed by EAE and CF. Analysis of the animals’ brain GABA levels revealed that after administering
P. edulis extract and its fractions, there was a substantial increase in GABA neurotransmitter concentration compared to the negative control. This suggests that the anxiolytic-like effects of
P. edulis are linked to the GABA-ergic system’s activity. The extracts may act as positive allosteric modulators of GABA, and their effects might be influenced by the presence or absence of endogenous neurotransmitters. The effects of CF and BF were particularly pronounced at an oral dose of 200 mg/kg, followed by the EAE fraction. These findings mirrored the results obtained from the elevated plus-maze model. Although there was no significant difference among the different
P. edulis treatment groups, it implies that the plant’s anxiolytic effects are likely due to a combination of active constituents, such as flavonoids, rather than a single chemical entity. These results align with Appel’s previous research, which suggested that
P. edulis’s effects on the central nervous system are mediated by GABA system modulation, including interactions with GABAA and GABAB receptors, as well as effects on GABA uptake. To sum up, the extract was found to increase GABA concentrations in the brain, suggesting that it functioned as a positive allosteric modulator of GABA. Metabolomics analyses did not reveal a correlation between the different metabolites and the observed activities, suggesting that the anxiolytic effect was likely due to an additive or synergistic effect of multiple compounds, as reported by Humer et al. in 2020 [
52].
2.2.6. Piper methysticum G. Forst. (Hawaiian Name: Awa/Kava)
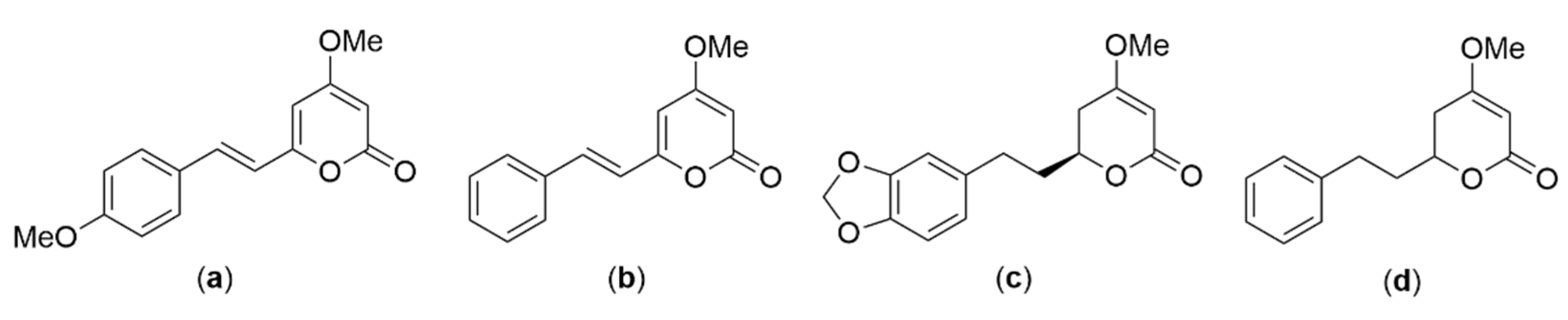
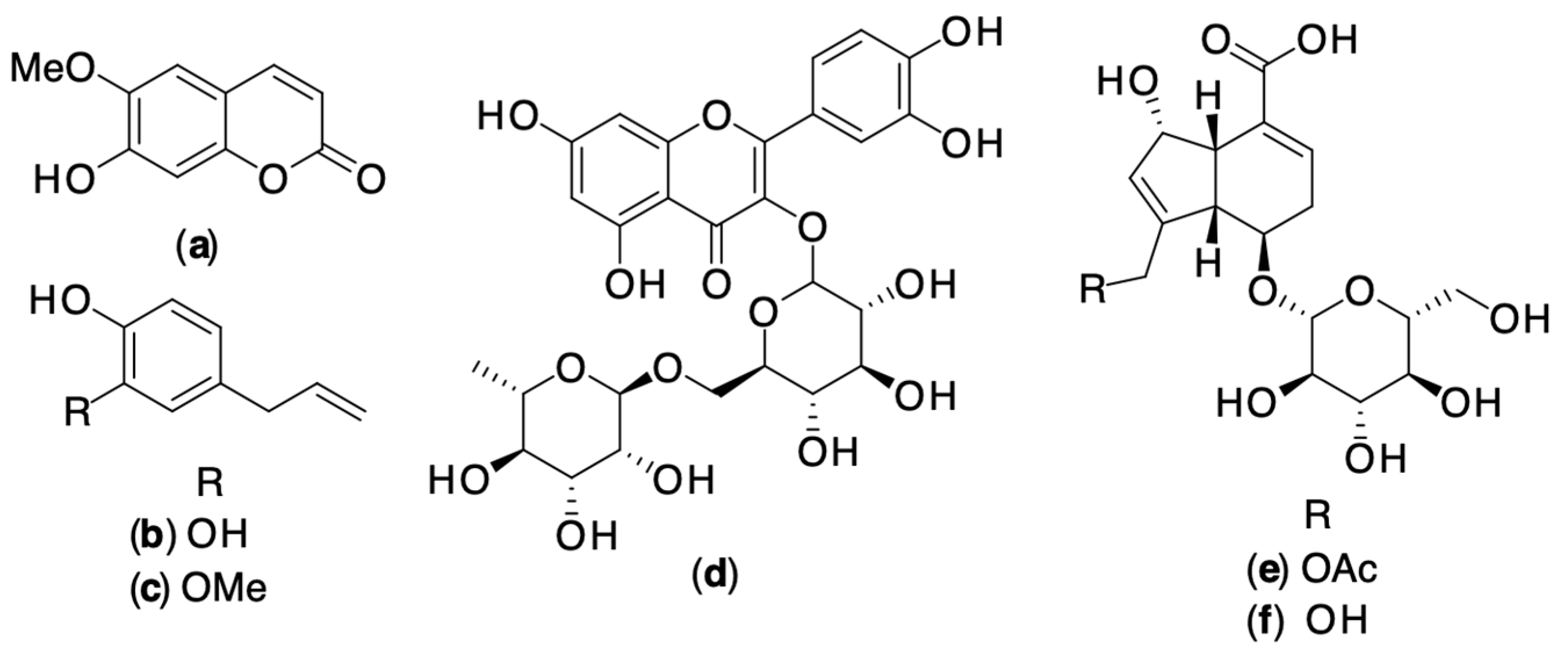
Piper methysticum or kava (Piperaceae) is native to islands of the Pacific Ocean. In 2020, Sarris et al. reported that an aqueous extract of dried
P. methysticum root, when taken in standardized doses of 120, 240, or 250 mg of kavalactones (
Figure 5), could not be used as a psychotropic medication to treat generalized anxiety disorder. In a study comprising 171 participants, an analysis was conducted to examine the impact of
P. methysticum versus placebo on anxiety reduction in 16 weeks. The results indicated that there was no significant difference in anxiety reduction between the two groups. However, it may be effective as an anxiolytic prior to a potential situational anxiogenic event or as an additional management of non-clinical anxiety and stress [
53].
Investigation of the potential impact of
P. methysticum extract on changes in behavior caused by amphetamine (AMPH) and its potential relationship with alterations in monoamine oxidase (MAO) activity. In the experiment, mice were administered either a vehicle or
P. methysticum extract through gavage, followed by either a vehicle or AMPH through intraperitoneal injection, with a two-hour interval. Behavioral tests (elevated plus maze, open field, stereotyped behavior, social interaction, and Y maze) and biochemical assessments (MAO-A and MAO-B activity in the cortex, hippocampus, and striatum) were conducted sequentially. The findings revealed that
P. methysticum extract demonstrated anxiolytic effects in the elevated plus maze test, increased locomotor activity in the open field test, and reduced MAO-A activity in the cortex, along with decreased MAO-B activity in the hippocampus.
P. methysticum extract counteracted the impact of AMPH on stereotyped behavior. When
P. methysticum extract and AMPH were combined, there was an increase in arm entries in the Y maze test, along with elevated MAO-B activity in the striatum. However,
P. methysticum extract did not mitigate the hyperlocomotion triggered by AMPH in the open field test. Notably,
P. methysticum extract did not influence social interaction, both in isolation and in combination with AMPH. In closing,
P. methysticum extract could prevent the appearance of stereotyped behavior induced by amphetamine (AMPH) in mice, suggesting it might be useful in alleviating psychotic symptoms in patients. Furthermore, the same dose of
P. methysticum extract that promoted anxiolytic effects was found to also reduce the stereotyped behavior, indicating its potential as a therapeutic for both anxiety and psychotic symptoms, as reported by Krum et al. in 2021 [
54].
The Committee on Herbal Medicinal Products (HMPC) assessment report on
P. methysticum has assumed that the true indication of
P. methysticum is generalized anxiety disorder (GAD according to ICD-10 F41.1). The analysis of data to determine which symptoms are best alleviated by
P. methysticum treatment. The study involved 156 patients, and it utilized a five-item rating scale to assess twelve common symptoms associated with nervous anxiety, tension, and restlessness. Additionally, details were collected regarding the therapeutic setting, the perceived onset time of the effects, and the safety of the treatment. The treatment period for patients had a median duration of 28 days. Notably, all individual symptoms displayed significant and clinically meaningful improvements. The most remarkable results were observed in cases of nervous tension and restlessness, particularly in patients with acute symptoms rather than chronic ones. The study also determined that the application of kava was exceptionally safe, a conclusion drawn from the evaluation of laboratory data. To review, the study supports the efficacy and safety of short-term
P. methysticum use in addressing “nervous anxiety, tension, and restlessness”, particularly in instances that are not chronic. These findings suggest that
P. methysticum could be clinically beneficial for conditions like context-related phobias as per ICD-10 F40, nervous tension (ICD-10 R45.0), or restlessness and excitation (ICD-10 R45.1). However, a study demonstrated that this assumption is unfounded and should be corrected. The clinical evidence suggested that
P. methysticum should be indicated for short-term situational anxiety, as defined by the German Commission E’s indication of “nervous anxiety, nervous tension and nervous restlessness”. This evidence also supported the safety of noble
P. methysticum extracts, particularly regarding liver adverse events, as described by Kuchta et al. and Yadav et al., respectively [
24,
55].
In 2022, Kenda et al. reviewed that various
P. methysticum extracts and isolated compounds have demonstrated interactions with GABAA receptors, inhibition of monoamine uptake through MAO-B inhibition and modulation of serotonin 5-HT1A receptors. In animal experiments involving rats and mice, diverse
P. methysticum extracts and compounds displayed sedative, calming, and muscle-relaxing effects. Additionally, some studies indicated potential anticonvulsive, spasmolytic, neuroprotective, and analgesic properties. Clinical trials investigating the efficacy of
P. methysticum preparations for anxiety disorders have yielded mixed outcomes. One randomized, double-blind controlled trial involving 135 participants in the kava-kava group and 135 participants in the placebo group revealed enhancements in anxiety symptoms and sleep. However, no significant distinctions between the groups were noted, implying that
P. methysticum did not provide more significant relief than the placebo. Another analysis encompassing a combined sample from three randomized, double-blind controlled trials did not reveal any improvement in the kava-kava treatment group. Notably, there was an absence of observed hepatotoxicity in individuals treated with
P. methysticum. In contrast, a separate randomized, double-blind, controlled study demonstrated a noteworthy reduction in anxiety within the
P. methysticum treatment group. This effect was particularly pronounced among individuals experiencing moderate to severe generalized anxiety disorder. Genetic polymorphisms in GABA transporters rs2601126 and rs2697153 were linked to this outcome. However, the
P. methysticum treatment group exhibited an increased incidence of headaches, while no discernible differences in liver function tests emerged between treatment groups. Overall, the clinical trials assessing
P. methysticum’s effectiveness in generalized anxiety or anxiety during (peri)menopause, as reviewed by the European Medicines Agency in 2016, exhibited significant limitations. These included the trials’ brevity and inadequate follow-up periods, heterogeneity in the anxiety-afflicted population, insufficient data on the proportion of responders, variations in supplement extraction methods, diverse reference compounds, and discrepancies in dosages employed [
56].
2.2.8. Rhodiola rosea L. (Introduced to Hawaii; Traditional Chinese Medicine (TCM): Hóng Jǐng Tiān)
Rhodiola rosea, commonly referred to as “roseroot”, “golden root” or “arctic root”, is a medicinal plant belonging to the Crassulaceae family. It has been used in traditional and popular medicine in many European and Asian countries [
59,
60]. In 2019, Yu et al. investigated the effects of
R. rosea on oxidative stress, anxiety, and depression in patients with obstructive sleep apnea (OSA). Ninety patients diagnosed with moderate to severe obstructive sleep apnea (OSA) and exhibiting negative emotions based on polysomnography (PSG) as well as self-assessment scales for depression (SDS) and anxiety (SAS) were enrolled from the respiratory department of our hospital between February 2015 and February 2018. Employing a random number table method, the patients were allocated into three groups: the non-invasive ventilator group, the
R. rosea + non-invasive ventilator group, and the
R. rosea group, with each group containing 30 patients. The non-invasive ventilator group received continuous positive airway pressure (CPAP) treatment for 3 months,
R. rosea + non-invasive ventilator group received oral
R. rosea capsules in addition to CPAP, and
R. rosea treatment group received only oral
R. rosea capsules for 3 months. Changes in SDS and SAS scores were measured before and after the interventions, and alterations in serum superoxide dismutase (SOD) and malondialdehyde (MDA) levels were assessed using enzyme-linked immunosorbent assays. Results showed no significant differences in SDS and SAS scores among the three groups (
p > 0.05). However,
R. rosea + non-invasive ventilator group exhibited decreased SDS and SAS scores after treatment (
p < 0.05) compared to the non-invasive ventilator group. Conversely, the
R. rosea treatment group experienced increased SDS and SAS scores after treatment (
p < 0.05). Notably, when comparing the
R. rosea + non-invasive ventilator group to the
R. rosea treatment group after treatment, the former demonstrated reduced SDS and SAS scores (
p < 0.05). Initially, there were no significant differences in serum SOD and MDA levels among the three groups (
p > 0.05). However, post-treatment, all groups exhibited elevated SOD levels and decreased MDA levels (
p < 0.05). Furthermore, in comparison to the non-invasive ventilator group after treatment, the
R. rosea + non-invasive ventilator group displayed decreased MDA levels in patients with elevated serum SOD levels (
p < 0.05), as did the
R. rosea treatment group (
p < 0.05). Ultimately, the
R. rosea + non-invasive ventilator group, when compared with
R. rosea treatment group, showed decreased levels of serum SOD and MDA after treatment (
p < 0.05). The study found that
R. rosea may improve negative emotions such as anxiety and depression by inhibiting oxygen free radicals and lipid peroxidation in patients with OSA [
61].
In 2019, Dinel et al. reported that examined the impact of repeated doses of a hydroethanolic root extract (HRE) from
R. rosea on the hypothalamic-pituitary-adrenal (HPA) response in a murine model subjected to mild acute stress, as well as elucidate the underlying mechanisms. The experiment involved Balb/c mice administered either HRE (5 g/kg) or a control substance orally for two weeks before undergoing a mild acute stress regimen (consisting of open-field and elevated plus maze tests). Levels of corticosterone, a stress hormone, were assessed in plasma samples obtained from the mandibular vein prior to stress initiation, as well as 30, 60, and 90 min thereafter. At the 90 min mark, mice were euthanized, and the hippocampus, prefrontal cortex, and amygdala were harvested for high-frequency RT-PCR analysis to evaluate gene expression patterns. Results demonstrated that, following the introduction of mild acute stress, mice treated with HRE exhibited lower corticosterone levels after 30 min compared to the control group. Notably, the corticosterone levels in the HRE-treated stressed mice resembled those in non-stressed mice from the HRE group. Additionally, administration of HRE led to specific alterations in gene expression in various brain regions, particularly the hippocampus and prefrontal cortex. These genetic changes encompass stress-responsive genes related to neuronal architecture, activation of the HPA axis, and circadian rhythm. Collectively, the findings underscore the potential utility of
R. rosea HRE in regulating reactivity to mild acute stress, as evidenced by its ability to diminish corticosterone levels and enhance the expression of stress-responsive genes, especially within the hippocampus and prefrontal cortex. This upregulation by
R. rosea hydroethanolic root extract was associated with damping of corticosterone secretion and a faster return to the basal profile, indicating improved adaptation of the animals receiving
R. rosea hydroethanolic root extract to a new environment. Further research is needed to understand the signaling pathways and transcription factors involved, as well as the impact of hydroethanolic root extract under stress conditions [
62].
R. rosea supplementation may have beneficial effects on mild to moderate depression, mild anxiety, and mood in clinical trials. The therapeutic effects of
R. rosea were observed to align with the specific medical conditions of the participants. Individuals with mild anxiety experienced a reduction in anxiety symptoms along with an uplift in mood. Those dealing with mild to moderate depression exhibited a decrease in depressive symptoms. In the case of individuals with major depression, there was a reduction in depression symptoms, though not to the same extent as seen with participants treated with sertraline, a conventional antidepressant. This suggests that
R. rosea might be less effective in treating major depression compared to standard antidepressants. Additionally, individuals experiencing fatigue and treated with
R. rosea showed improvements in fatigue symptoms but not in depression. Similarly, participants undergoing mentally stressful situations reported a decrease in mental fatigue and an increase in overall well-being, as summarized by Konstantinos et al. in 2020 [
63]. Further research with randomized controlled trials is needed to confirm these effects.
The use of
R. rosea as a medicinal treatment for stress-induced conditions and disorders has been validated by extensive research, making it an effective remedy for stress-related issues.
R. rosea herbal preparations have demonstrated promising results in treating mild-to-moderate depression and generalized anxiety. Traditional uses of
R. rosea for stress and mood-related issues align with its potential to modulate anxiety and mood by hindering physiological stress responses. Clinical trials have indicated the effectiveness of
R. rosea in reducing symptoms of generalized anxiety disorder and mild anxiety. Additionally, studies have shown that
R. rosea can effectively alleviate symptoms of mild to moderate depression. Its antidepressant effects have been compared to conventional antidepressants like sertraline, with
R. rosea showing similar odds of improvement and better tolerability. Combining
R. rosea with sertraline has also led to improved outcomes in patients with major depressive disorder. Preclinical research suggests that
R. rosea’s mechanisms involve influencing neuropeptide-Y expression, gene regulation related to behavior and mood, modulation of stress response pathways, and inhibition of monoamine oxidases. Moreover,
R. rosea has shown potential for enhancing serotonin levels and neural stem cell proliferation in the hippocampus. Overall, these findings support the notion that
R. rosea holds promise as a natural remedy for anxiety and depression, offering potential benefits and a more favorable risk-to-benefit ratio compared to certain conventional antidepressants, as described by Ivanova et al. in 2022 [
60].




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







