
Exploring the Potent Anticancer Activity of Essential Oils and Their Bioactive Compounds: Mechanisms and Prospects for Future Cancer Therapy

 , ,
, , 
Abstract
:1. Introduction
1.1. Essential Oils
1.2. Chemical Composition
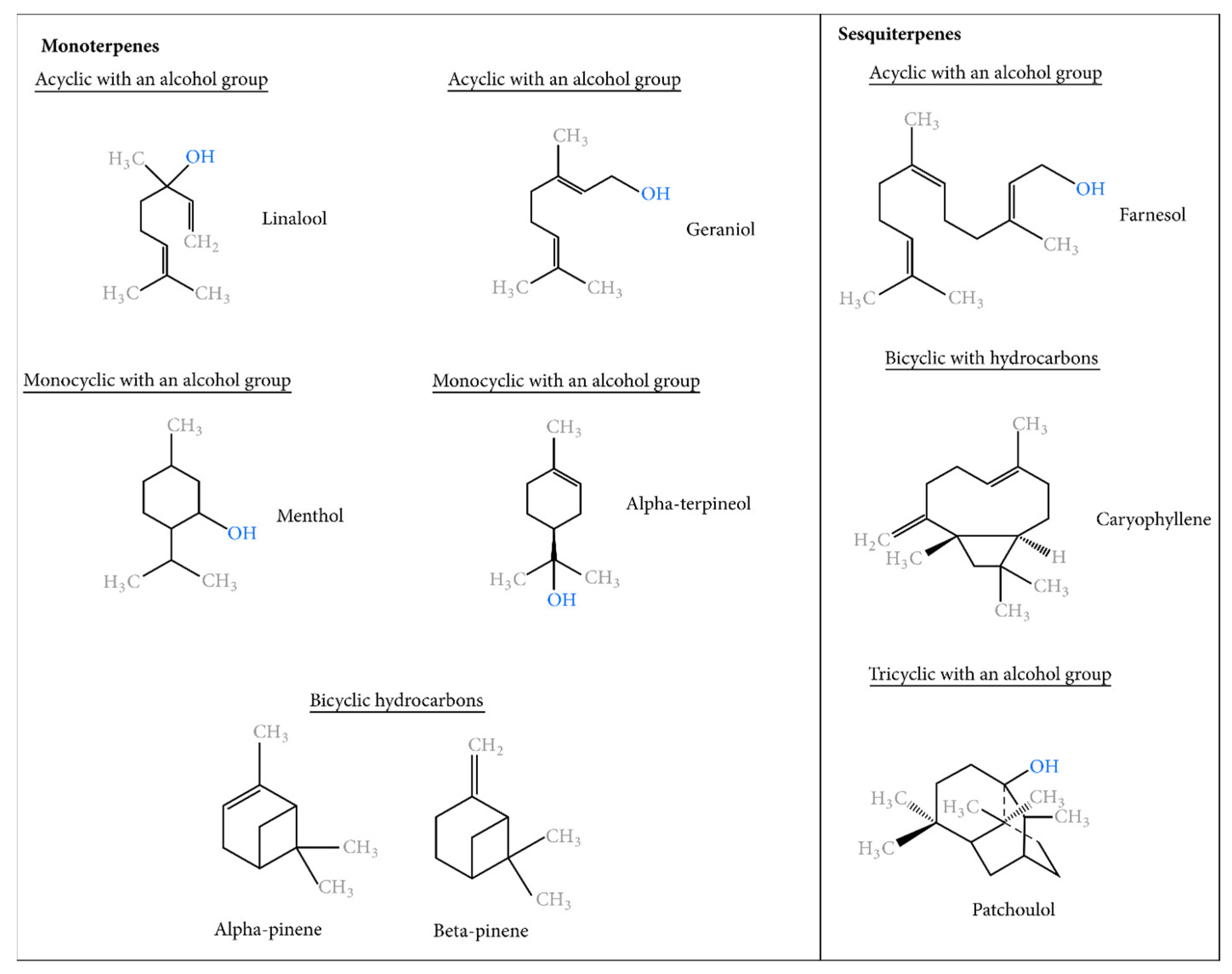
1.3. The Essential Groups of Essential Oils
1.4. Anti-Cancer Properties of Essential Oils
1.5. Antitumor Properties of Essential Oils
1.6. Antiproliferative Mechanisms of Action of Essential Oils
1.7. Cancer Cell Specificity of Essential Oils
1.8. Synergism of Essential Oils Extracts with Conventional Chemotherapeutic Agents: Potential of Combination Therapy Using Essential Oils
1.9. Mechanisms of Anti-Cancer Action
1.10. Suppression of Inflammation and Reduction of Oxidative Stress
1.11. Generation of Reactive OXYGEN Species within Cancer Cells
1.12. Overexpression and Detoxification of the Liver
1.13. Alteration of Mitochondrial Membrane Potential
1.14. Activation of Apoptosis by Caspases
1.15. Cell Cycle Arrest
1.16. Modification of Signaling Pathways (Disabling the PI3K/Akt/NF-κB Pathway)
1.17. Modification of the AMPK Pathway
1.18. Modulation in the Expression of MAPK/ERK Proteins
1.19. Inhibition of the Activity of 3-Hydroxy-3-Methylglutaryl-Coenzyme A
1.20. Anti-Angiogenic Effect
1.21. Modification of Histones
1.22. Inhibition of Other Factors Involved in the Induction of Tumorigenesis
1.23. Routes of Administration of Essential Oils
1.24. Toxicity of Essential Oils
2. Conclusions
3. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cammarota, G.; Ianiro, G.; Ahern, A.; Carbone, C.; Temko, A.; Claesson, M.J.; Gasbarrini, A.; Tortora, G. Gut microbiome, big data and machine learning to promote precision medicine for cancer. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Kushi, L.H.; Doyle, C.; McCullough, M.; Rock, C.L.; Demark-Wahnefried, W.; Bandera, E.V.; Gapstur, S.; Patel, A.V.; Andrews, K.; Gansler, T.; et al. American Cancer Society Guidelines on Nutrition and Physical Activity for cancer prevention: Reducing the risk of cancer with healthy food choices and physical activity. CA Cancer J. Clin. 2012, 62, 30–67. [Google Scholar] [CrossRef] [Green Version]
- Czarnecka, A.M.; Bartnik, E.; Fiedorowicz, M.; Rutkowski, P. Targeted therapy in melanoma and mechanisms of resistance. Int. J. Mol. Sci. 2020, 21, 4576. [Google Scholar] [CrossRef] [PubMed]
- Cristini, V.; Lowengrub, J. Multiscale Modeling of Cancer: An Integrated Experimental and Mathematical Modeling Approach; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Ladeiro, Y.; Couchy, G.; Balabaud, C.; Bioulac-Sage, P.; Pelletier, L.; Rebouissou, S.; Zucman-Rossi, J. MicroRNA profiling in hepatocellular tumors is associated with clinical features and oncogene/tumor suppressor gene mutations. Hepatology 2008, 47, 1955–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. WHO Report on the Global Tobacco Epidemic, 2008: The MPOWER Package; World Health Organization: Geneva, Switzerland, 2008. [Google Scholar]
- Cao, W.; Chen, H.D.; Yu, Y.W.; Li, N.; Chen, W.Q. Changing profiles of cancer burden worldwide and in China: A secondary analysis of the global cancer statistics 2020. Chin. Med. J. 2021, 134, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, N.; Chakrabarty, A.M.; Fialho, A.M. Engineering of bacterial strains and their products for cancer therapy. Appl. Microbiol. Biotechnol. 2013, 97, 5189–5199. [Google Scholar] [CrossRef]
- Cragg, G.M.; Grothaus, P.G.; Newman, D.J. Impact of natural products on developing new anti-cancer agents. Chem. Rev. 2009, 109, 3012–3043. [Google Scholar] [CrossRef]
- Abdoul-Latif, F.M.; Oumaskour, K.; Boujaber, N.; Ainane, A.; Mohamed, J.; Ainane, T. Formulations of a cosmetic product for hair care based on extract of the microalga Isochrysis galbana: In vivo and in vitro activities. J. Anal. Sci. Appl. Biotechnol. 2021, 3, 15–19. [Google Scholar]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A review of its effects on human health. Foods 2017, 6, 92. [Google Scholar] [CrossRef] [Green Version]
- Balandrin, M.F.; Kinghorn, A.D.; Farnsworth, N.R. Plant-derived natural products in drug discovery and development: An overview. In Human Medicinal Agents from Plants; American Chemical Society: Washington, DC, USA, 1993. [Google Scholar]
- Mohamed Abdoul-Latif, F.; El Montassir, Z.; Ainane, A.; Gharby, S.; Sakar, E.H.; Merito, A.; Mohamed, J.; Ainane, T. Use of Thymus Plants as an Ecological Filler in Urea-Formaldehyde Adhesives Intended for Bonding Plywood. Processes 2022, 10, 2209. [Google Scholar] [CrossRef]
- Al-Radadi, N.S. Biogenic proficient synthesis of (Au-NPs) via aqueous extract of Red Dragon Pulp and seed oil: Characterization, antioxidant, cytotoxic properties, anti-diabetic anti-inflammatory, anti-Alzheimer and their anti-proliferative potential against cancer cell lines. Saudi J. Biol. Sci. 2022, 29, 2836–2855. [Google Scholar] [PubMed]
- Majdalawieh, A.F.; Fayyad, M.W.; Nasrallah, G.K. Anti-cancer properties and mechanisms of action of thymoquinone, the major active ingredient of Nigella sativa. Crit. Rev. Food Sci. Nutr. 2017, 57, 3911–3928. [Google Scholar] [CrossRef] [PubMed]
- Bhavaniramya, S.; Vishnupriya, S.; Al-Aboody, M.S.; Vijayakumar, R.; Baskaran, D. Role of essential oils in food safety: Antimicrobial and antioxidant applications. Grain Oil Sci. Technol. 2019, 2, 49–55. [Google Scholar] [CrossRef]
- Rios, J.L. Essential oils: What they are and how the terms are used and defined. In Essential Oils in Food Preservation, Flavor and Safety; Academic Press: Cambridge, MA, USA, 2016; pp. 3–10. [Google Scholar]
- Alam, M.; Kamal, A.; Upadhyay, T.K.; Upadhye, V.J. The role and effects of aroma: Status and trends. In Aromatic Plants: The Technology, Human Welfare and Beyond; Nova Science Publishers: Hauppauge, NY, USA, 2010; p. 169. [Google Scholar]
- Khammour, F.; Abdoul-Latif, F.M.; Ainane, A.; Mohamed, J.; Ainane, T. Eco-friendly adsorbent from waste of mint: Application for the removal of hexavalent chromium. J. Chem. 2021, 2021, 8848964. [Google Scholar] [CrossRef]
- Mukhija, M.; Sundriyal, A.; Loshali, A. Essential Oils and Their Biological Applications: Extraction methods, Types, Biological Activities, Antimicrobial Fumes. In Handbook of Research on Advanced Phytochemicals and Plant-Based Drug Discovery; IGI Global: Hershey, PA, USA, 2022; pp. 395–412. [Google Scholar]
- Butnariu, M. Plants as source of essential oils and perfumery applications. In Bioprospecting of Plant Biodiversity for Industrial Molecules; John Wiley & Sons: Hoboken, NJ, USA, 2021; pp. 261–292. [Google Scholar]
- Rehman, R.; Hanif, M.A.; Mushtaq, Z.; Al-Sadi, A.M. Biosynthesis of essential oils in aromatic plants: A review. Food Rev. Int. 2016, 32, 117–160. [Google Scholar] [CrossRef]
- Atemni, I.; Touijer, H.; Hjouji, K.; Tlemcani, S.; Ainane, T.; Taleb, M.; Rais, Z. Effect of bone meal on growth traits, photosynthetic pigment content, and essential oil chemical composition of Pelargonium graveolens. Ind. Crops Prod. 2023, 202, 117021. [Google Scholar] [CrossRef]
- Zeroual, A.; Sakar, E.H.; Ibourki, M.; Bijla, L.; Ainane, A.; Mahjoubi, F.; Chaouch, M.; Gharby, S.; Chaqroune, A.; Ainane, T. Phytochemical screening and mineral profiling of wild and cultivated rosemary (Rosmarinus officinalis L.) from Taounate region (northern Morocco). Pharmacol. Online 2021, 2, 576–582. [Google Scholar]
- Cseke, L.J.; Kaufman, P.B.; Kirakosyan, A. The biology of essential oils in the pollination of flowers. Nat. Prod. Commun. 2007, 2, 1934578X0700201225. [Google Scholar] [CrossRef]
- Sharmeen, J.B.; Mahomoodally, F.M.; Zengin, G.; Maggi, F. Essential oils as natural sources of fragrance compounds for cosmetics and cosmeceuticals. Molecules 2021, 26, 666. [Google Scholar] [CrossRef]
- Ainane, T.; Elkouali, M.H.; Ainane, A.; Talbi, M. Moroccan traditional fragrance based essential oils: Preparation, composition and chemical identification. Der Pharma Chem. 2014, 6, 84–89. [Google Scholar]
- Bilia, A.R.; Guccione, C.; Isacchi, B.; Righeschi, C.; Firenzuoli, F.; Bergonzi, M.C. Essential oils loaded in nanosystems: A developing strategy for a successful therapeutic approach. Evid. -Based Complement. Altern. Med. 2014, 2014, 651593. [Google Scholar] [CrossRef] [Green Version]
- Abdoul-latif, F.M.; Ainane, A.; Abdoul-latif, T.M.; Ainane, T. Chemical study and evaluation of insectical properties of African Lippia citriodora essential oil. J. Biopestic. 2020, 13, 119–126. [Google Scholar] [CrossRef]
- Ignea, C.; Raadam, M.H.; Koutsaviti, A.; Zhao, Y.; Duan, Y.T.; Harizani, M.; Miettinen, K.; Georgantea, P.; Rosenfeldt, M.; Viejo-Ledesma, S.E.; et al. Expanding the terpene biosynthetic code with non-canonical 16 carbon atom building blocks. Nat. Commun. 2022, 13, 5188. [Google Scholar] [CrossRef]
- Hillier, S.G.; Lathe, R. Terpenes, hormones and life: Isoprene rule revisited. J. Endocrinol. 2019, 242, R9–R22. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.H.; Hsieh, F.L.; Ko, T.P.; Teng, K.H.; Liang, P.H.; Wang, A.H.J. Structure of a heterotetrameric geranyl pyrophosphate synthase from mint (Mentha piperita) reveals intersubunit regulation. Plant Cell 2010, 22, 454–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barja Afonso, M. Exploring the Diversity of Geranylgeranyl Diphosphate Synthases in Arabidopsis thaliana and Solanum lycopersicum; Universitat Autònoma de Barcelona: Bellaterra, Spain, 2019. [Google Scholar]
- Abdoul-Latif, F.M.; Ejjabraoui, M.; Ainane, A.; Hachi, T.; Mohamed, J.; Oumaskour, K.; Boujaber, N.; El Montassir, Z.; Ainane, T. Correlation of the Diffusion Parameters and the Biological Activities in the Formulation of Pinus halepensis Essential Oil in Phosphogypsum Material. Appl. Sci. 2023, 13, 5358. [Google Scholar] [CrossRef]
- Stephane, F.F.Y.; Jules, B.K.J. Terpenoids as important bioactive constituents of essential oils. In Essential Oils-Bioactive Compounds, New Perspectives and Applications; IntechOpen: London, UK, 2020. [Google Scholar]
- Sadgrove, N.J.; Padilla-González, G.F.; Phumthum, M. Fundamental chemistry of essential oils and volatile organic compounds, methods of analysis and authentication. Plants 2022, 11, 789. [Google Scholar] [CrossRef] [PubMed]
- Vassão, D.G.; Davin, L.B.; Lewis, N.G. Metabolic engineering of plant allyl/propenyl phenol and lignin pathways: Future potential for biofuels/bioenergy, polymer intermediates, and specialty chemicals? Adv. Plant Biochem. Mol. Biol. 2008, 1, 385–428. [Google Scholar]
- Mohamed Abdoul-Latif, F.; Ainane, A.; Houmed Aboubaker, I.; Merito Ali, A.; El Montassir, Z.; Kciuk, M.; Mohamed, J.; Ainane, T. Chemical Composition of the Essential Oil of Catha edulis Forsk from Djibouti and Its Toxicological Investigations In Vivo and In Vitro. Processes 2023, 11, 1324. [Google Scholar] [CrossRef]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential oils’ chemical characterization and investigation of some biological activities: A critical review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Pengelly, A. The Constituents of Medicinal Plants; CABI: Wallingford, UK, 2021. [Google Scholar]
- Bhatla, S.C.; Lal, A.M.; Bhatla, S.C. Secondary metabolites. In Plant Physiology Development and Metabolism; Springer: Berlin/Heidelberg, Germany; Cham, Switzerland, 2018; pp. 1099–1166. [Google Scholar]
- World Health Organization (WHO). Internacional Agency for Research on Cancer. Cancer Fact Sheets; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- World Health Organization. Cancer; WHO: Geneva, Switzerland, 2015; Available online: http://www.who.int/mediacentre/factsheets/fs297/en (accessed on 1 May 2023).
- Ferguson, L.R.; Chen, H.; Collins, A.R.; Connell, M.; Damia, G.; Dasgupta, S.; Malhotra, M.; Meeker, A.K.; Amedei, A.; Amin, A.; et al. Genomic instability in human cancer: Molecular insights and opportunities for therapeutic attack and prevention through diet and nutrition. In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2015; Volume 35, pp. S5–S24. [Google Scholar]
- Fresco, P.; Borges, F.I.G.M.; Diniz, C.; Marques, M.P.M. New insights on the anticancer properties of dietary polyphenols. Med. Res. Rev. 2006, 26, 747–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morse, M.A.; Stoner, G.D. Cancer chemoprevention: Principles and prospects. Carcinogenesis 1993, 14, 1737–1746. [Google Scholar] [CrossRef]
- Amin, A.; Gali-Muhtasib, H.; Ocker, M.; Schneider-Stock, R. Overview of major classes of plant-derived anticancer drugs. Int. J. Biomed. Sci. IJBS 2009, 5, 1. [Google Scholar] [PubMed]
- Weaver, B.A. How Taxol/paclitaxel kills cancer cells. Mol. Biol. Cell 2014, 25, 2677–2681. [Google Scholar] [CrossRef] [PubMed]
- Holton, R.A.; Kim, H.B.; Somoza, C.; Liang, F.; Biediger, R.J.; Boatman, P.D.; Shindo, M.; Smith, C.C.; Kim, S. First total synthesis of taxol. 2. Completion of the C and D rings. J. Am. Chem. Soc. 1994, 116, 1599–1600. [Google Scholar] [CrossRef]
- Altshuler, O.; Abu-Abied, M.; Chaimovitsh, D.; Shechter, A.; Frucht, H.; Dudai, N.; Sadot, E. Enantioselective effects of (+)-and (−)-citronellal on animal and plant microtubules. J. Nat. Prod. 2013, 76, 1598–1604. [Google Scholar] [CrossRef]
- Edris, A.E. Pharmaceutical and therapeutic potentials of essential oils and their individual volatile constituents: A review. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2007, 21, 308–323. [Google Scholar] [CrossRef]
- Sitarek, P.; Rijo, P.; Garcia, C.; Skała, E.; Kalemba, D.; Białas, A.J.; Szemraj, J.; Pytel, D.; Toma, M.; Wysokińska, H.; et al. Antibacterial, anti-inflammatory, antioxidant, and antiproliferative properties of essential oils from hairy and normal roots of Leonurus sibiricus L. and their chemical composition. Oxidative Med. Cell. Longev. 2017, 2017, 7384061. [Google Scholar] [CrossRef] [Green Version]
- Abdoul-Latif, F.M.; Elmi, A.; Merito, A.; Nour, M.; Risler, A.; Ainane, A.; Bignon, J.; Ainane, T. Essential oils of Tagetes minuta and Lavandula coronopifolia from Djibouti: Chemical composition, antibacterial activity and cytotoxic activity against various human cancer cell lines. Int. J. Plant Biol. 2022, 13, 26. [Google Scholar] [CrossRef]
- Mohamed Abdoul-Latif, F.; Elmi, A.; Merito, A.; Nour, M.; Risler, A.; Ainane, A.; Bignon, J.; Ainane, T. Chemical Analysis of Essential Oils of Cymbopogon schoenanthus (L.) Spreng. and Nepeta azurea R. Br. ex Benth from Djbouti, In-Vitro Cytotoxicity against Cancer Cell Lines and Antibacterial Activities. Appl. Sci. 2022, 12, 8699. [Google Scholar] [CrossRef]
- Mohamed Abdoul-Latif, F.; Elmi, A.; Merito, A.; Nour, M.; Risler, A.; Ainane, A.; Bignon, J.; Ainane, T. Essential Oils of Ocimum basilicum L. and Ocimum americanum L. from Djibouti: Chemical Composition, Antimicrobial and Cytotoxicity Evaluations. Processes 2022, 10, 1785. [Google Scholar] [CrossRef]
- Elmi, A.; Spina, R.; Risler, A.; Philippot, S.; Mérito, A.; Duval, R.E.; Abdoul-Latif, F.M.; Laurain-Mattar, D. Evaluation of antioxidant and antibacterial activities, cytotoxicity of Acacia seyal Del bark extracts and isolated compounds. Molecules 2020, 25, 2392. [Google Scholar] [CrossRef]
- Swain, S.S.; Paidesetty, S.K.; Padhy, R.N.; Hussain, T. Nano-technology platforms to increase the antibacterial drug suitability of essential oils: A drug prospective assessment. OpenNano 2023, 9, 100115. [Google Scholar] [CrossRef]
- Hallin, J.; Bowcut, V.; Calinisan, A.; Briere, D.M.; Hargis, L.; Engstrom, L.D.; Laguer, J.; Medwid, J.; Vanderpool, D.; Lifset, E.; et al. Anti-tumor efficacy of a potent and selective non-covalent KRASG12D inhibitor. Nat. Med. 2022, 28, 2171–2182. [Google Scholar] [CrossRef] [PubMed]
- Valizadeh, A.; Khaleghi, A.A.; Roozitalab, G.; Osanloo, M. High anticancer efficacy of solid lipid nanoparticles containing Zataria multiflora essential oil against breast cancer and melanoma cell lines. BMC Pharmacol. Toxicol. 2021, 22, 52. [Google Scholar] [CrossRef] [PubMed]
- Abdoul-Latif, F.M.; Elmi, A.; Merito, A.; Nour, M.; Risler, A.; Ainane, A.; Bignon, J.; Ainane, T. Essential oil of Ruta chalepensis L. from Djibouti: Chemical Analysis and Modeling of In Vitro Anticancer Profiling. Separations 2022, 9, 387. [Google Scholar] [CrossRef]
- Topçu, G.; Gören, A.C.; Bilsel, G.; Bilsel, M.; Çakmak, O.; Schilling, J.; Kingston, D.G. Cytotoxic Activity and Essential Oil Composition of Leaves and Berries of Juniperus excelsa. Pharm. Biol. 2005, 43, 125–128. [Google Scholar] [CrossRef]
- Buhagiar, J.A.; Podesta, M.T.; Wilson, A.P.; Micallef, M.J.; Ali, S. The induction of apoptosis in human melanoma, breast and ovarian cancer cell lines using an essential oil extract from the conifer Tetraclinis articulata. Anticancer. Res. 1999, 19, 5435–5443. [Google Scholar]
- Sylvestre, M.; Pichette, A.; Longtin, A.; Nagau, F.; Legault, J. Essential oil analysis and anticancer activity of leaf essential oil of Croton flavens L. from Guadeloupe. J. Ethnopharmacol. 2006, 103, 99–102. [Google Scholar] [CrossRef]
- Gharby, S.; Asdadi, A.; Ibourki, M.; Hamdouch, A.; Ainane, T.; Hassani, L.A.I. Chemical characterization of the essential oil from aerial parts of Lavandula rejdalii Upson & Jury, a medicinal plant endemic to Morocco. J. Essent. Oil Bear. Plants 2020, 23, 1422–1427. [Google Scholar]
- De Lima, V.T.; Vieira, M.C.; Kassuya, C.A.L.; Cardoso, C.A.L.; Alves, J.M.; Foglio, M.A.; De Carvalho, J.E.; Formagio, A.S.N. Chemical composition and free radical-scavenging, anticancer and anti-inflammatory activities of the essential oil from Ocimum kilimandscharicum. Phytomedicine 2014, 21, 1298–1302. [Google Scholar] [CrossRef] [PubMed]
- Mitropoulou, G.; Fitsiou, E.; Stavropoulou, E.; Papavassilopoulou, E.; Vamvakias, M.; Pappa, A.; Oreopoulou, A.; Kourkoutas, Y. Composition, antimicrobial, antioxidant, and antiproliferative activity of Origanum dictamnus (dittany) essential oil. Microb. Ecol. Health Dis. 2015, 26, 26543. [Google Scholar] [PubMed]
- Nikolić, M.; Glamočlija, J.; Ferreira, I.C.; Calhelha, R.C.; Fernandes, Â.; Marković, T.; Marković, D.; Giweli, A.; Soković, M. Chemical composition, antimicrobial, antioxidant and antitumor activity of Thymus serpyllum L.; Thymus algeriensis Boiss. and Reut and Thymus vulgaris L. essential oils. Ind. Crops Prod. 2014, 52, 183–190. [Google Scholar] [CrossRef]
- Russo, A.; Formisano, C.; Rigano, D.; Senatore, F.; Delfine, S.; Cardile, V.; Rosselli, S.; Bruno, M. Chemical composition and anticancer activity of essential oils of Mediterranean sage (Salvia officinalis L.) grown in different environmental conditions. Food Chem. Toxicol. 2013, 55, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Ainane, T.; Abdoul-Latif, F.M.; Baghouz, A.; El Montassir, Z.; Attahar, W.; Ainane, A.; Giuffrè, A.M. Correction: Essential oils rich in pulegone for insecticide purpose against legume bruchus species: Case of Ziziphora hispanica L. and Mentha pulegium L. AIMS Agric. Food 2023, 8, 236–238. [Google Scholar] [CrossRef]
- Sylvestre, M.; Pichette, A.; Lavoie, S.; Longtin, A.; Legault, J. Composition and cytotoxic activity of the leaf essential oil of Comptonia peregrina (L.) Coulter. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2007, 21, 536–540. [Google Scholar]
- Cole, R.A.; Bansal, A.; Moriarity, D.M.; Haber, W.A.; Setzer, W.N. Chemical composition and cytotoxic activity of the leaf essential oil of Eugenia zuchowskiae from Monteverde, Costa Rica. J. Nat. Med. 2007, 61, 414–417. [Google Scholar] [CrossRef]
- Hou, J.; Sun, T.; Hu, J.; Chen, S.; Cai, X.; Zou, G. Chemical composition, cytotoxic and antioxidant activity of the leaf essential oil of Photinia serrulata. Food Chem. 2007, 103, 355–358. [Google Scholar] [CrossRef]
- Abdoul-Latif, F.M.; Ainane, A.; Merito, A.; Ainane, T. Chemical composition and biological activities of essential oils from Djibouti. J. Anal. Sci. Appl. Biotechnol. 2022, 4, 1–9. [Google Scholar]
- Gudi, V.A.; Singh, S.V. Effect of diallyl sulfide, a naturally occurring anti-carcinogen, on glutathione-dependent detoxification enzymes of female CD-1 mouse tissues. Biochem. Pharmacol. 1991, 42, 1261–1265. [Google Scholar] [CrossRef]
- Girola, N.; Figueiredo, C.R.; Farias, C.F.; Azevedo, R.A.; Ferreira, A.K.; Teixeira, S.F.; Capello, T.M.; Martins, E.G.; Matsuo, A.L.; Travassos, L.R.; et al. Camphene isolated from essential oil of Piper cernuum (Piperaceae) induces intrinsic apoptosis in melanoma cells and displays antitumor activity in vivo. Biochem. Biophys. Res. Commun. 2015, 467, 928–934. [Google Scholar] [CrossRef]
- Arunasree, K.M. Anti-proliferative effects of carvacrol on a human metastatic breast cancer cell line, MDA-MB 231. Phytomedicine 2010, 17, 581–588. [Google Scholar] [CrossRef]
- Suhail, M.M.; Wu, W.; Cao, A.; Mondalek, F.G.; Fung, K.M.; Shih, P.T.; Fang, Y.-T.; Woolley, C.; Young, G.; Lin, H.-K. Boswellia sacra essential oil induces tumor cell-specific apoptosis and suppresses tumor aggressiveness in cultured human breast cancer cells. BMC Complement. Altern. Med. 2011, 11, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qurishi, Y.; Hamid, A.; Zargar, M.A.; Singh, S.K.; Saxena, A.K. Potential role of natural molecules in health and disease: Importance of boswellic acid. J. Med. Plants Res. 2010, 4, 2778–2785. [Google Scholar]
- Queiroz, R.M.D.; Takiya, C.M.; Guimarães, L.P.T.P.; Rocha, G.D.G.; Alviano, D.S.; Blank, A.F.; Alviano, C.S.; Gattass, C.R. Apoptosis-inducing effects of Melissa officinalis L. essential oil in glioblastoma multiforme cells. Cancer Investig. 2014, 32, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, B.Y.; Sarker, M.M.R.; Kamarudin, M.N.A.; Mohan, G. Antiproliferative and apoptosis inducing effects of citral via p53 and ROS-induced mitochondrial-mediated apoptosis in human colorectal HCT116 and HT29 cell lines. Biomed. Pharmacother. 2017, 96, 834–846. [Google Scholar] [CrossRef]
- Thomas, M.L.; De Antueno, R.; Coyle, K.M.; Sultan, M.; Cruickshank, B.M.; Giacomantonio, M.A.; Giacomantonio, C.A.; Duncan, R.; Marcato, P. Citral reduces breast tumor growth by inhibiting the cancer stem cell marker ALDH1A3. Mol. Oncol. 2016, 10, 1485–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naz, F.; Khan, F.I.; Mohammad, T.; Khan, P.; Manzoor, S.; Hasan, G.M.; Lobb, K.A.; Luqman, S.; Islam, A.; Ahmad, F.; et al. Investigation of molecular mechanism of recognition between citral and MARK4: A newer therapeutic approach to attenuate cancer cell progression. Int. J. Biol. Macromol. 2018, 107, 2580–2589. [Google Scholar] [CrossRef]
- Fayard, E.; Tintignac, L.A.; Baudry, A.; Hemmings, B.A. Protein kinase B/Akt at a glance. J. Cell Sci. 2005, 118, 5675–5678. [Google Scholar] [CrossRef] [Green Version]
- Seal, S.; Chatterjee, P.; Bhattacharya, S.; Pal, D.; Dasgupta, S.; Kundu, R.; Mukherjee, S.; Bhattacharya, S.; Bhuyan, M.; Bhattacharyya, P.R.; et al. Vapor of volatile oils from Litsea cubeba seed induces apoptosis and causes cell cycle arrest in lung cancer cells. PLoS ONE 2012, 7, e47014. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.C.; Chung, J.G.; Tsai, S.J.; Yang, J.H.; Sheen, L.Y. Differential effects of allyl sulfides from garlic essential oil on cell cycle regulation in human liver tumor cells. Food Chem. Toxicol. 2004, 42, 1937–1947. [Google Scholar] [CrossRef]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehne, N.; Mora, J.; Namgaladze, D.; Weigert, A.; Brüne, B. Cancer cell and macrophage cross-talk in the tumor microenvironment. Curr. Opin. Pharmacol. 2017, 35, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Ben-Neriah, Y.; Karin, M. Inflammation meets cancer, with NF-κB as the matchmaker. Nat. Immunol. 2011, 12, 715–723. [Google Scholar] [CrossRef]
- Hassan, S.B.; Gali-Muhtasib, H.; Göransson, H.; Larsson, R. Alpha terpineol: A potential anticancer agent which acts through suppressing NF-κB signalling. Anticancer. Res. 2010, 30, 1911–1919. [Google Scholar]
- Deeb, S.J. Enhancement of Cell Death by Linalyl Acetate and [alpha]-Terpineol through Targeting the Nuclear Factor-[kappa] B Activation Pathway in Human Colon Cancer Cells-by Sally Joseph Deeb. Ph.D. Thesis, American University of Beirut, Beirut, Lebanon, 2019. [Google Scholar]
- Feng, S.S.; Chien, S. Chemotherapeutic engineering: Application and further development of chemical engineering principles for chemotherapy of cancer and other diseases. Chem. Eng. Sci. 2003, 58, 4087–4114. [Google Scholar] [CrossRef]
- Chari, R.V. Targeted cancer therapy: Conferring specificity to cytotoxic drugs. Acc. Chem. Res. 2008, 41, 98–107. [Google Scholar] [CrossRef]
- Çetinus, E.; Temiz, T.; Ergül, M.; Altun, A.; Çetinus, Ş.; Kaya, T. Thyme essential oil inhibits proliferation of DLD-1 colorectal cancer cells through antioxidant effect. Cumhur. Med. J. 2013, 35, 14–24. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yue, Y.; Runwei, Y.; Guolin, Z. Cytotoxic, apoptotic and antioxidant activity of the essential oil of Amomum tsao-ko. Bioresour. Technol. 2010, 101, 4205–4211. [Google Scholar] [CrossRef] [PubMed]
- Mesa-Arango, A.C.; Montiel-Ramos, J.; Zapata, B.; Durán, C.; Betancur-Galvis, L.; Stashenko, E. Citral and carvone chemotypes from the essential oils of Colombian Lippia alba (Mill.) NE Brown: Composition, cytotoxicity and antifungal activity. Memórias Inst. Oswaldo Cruz 2009, 104, 878–884. [Google Scholar] [CrossRef] [Green Version]
- Frank, M.B.; Yang, Q.; Osban, J.; Azzarello, J.T.; Saban, M.R.; Saban, R.; Ashley, R.A.; Welter, J.C.; Fung, K.-M.; Lin, H.-K. Frankincense oil derived from Boswellia carteri induces tumor cell specific cytotoxicity. BMC Complement. Altern. Med. 2009, 9, 6. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.L.D.; Chaar, J.D.S.; Figueiredo, P.D.M.S.; Yano, T. Cytotoxic evaluation of essential oil from Casearia sylvestris Sw on human cancer cells and erythrocytes. Acta Amaz. 2008, 38, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.L.D.; Figueiredo, P.M.; Yano, T. Cytotoxic evaluation of essential oil from Zanthoxylum rhoifolium Lam. leaves. Acta Amaz. 2007, 37, 281–286. [Google Scholar] [CrossRef] [Green Version]
- Amiel, E.; Ofir, R.; Dudai, N.; Soloway, E.; Rabinsky, T.; Rachmilevitch, S. β-Caryophyllene, a compound isolated from the biblical balm of gilead (Commiphora gileadensis), is a selective apoptosis inducer for tumor cell lines. Evid.-Based Complement. Altern. Med. 2012, 2012, 872394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sœur, J.; Marrot, L.; Perez, P.; Iraqui, I.; Kienda, G.; Dardalhon, M.; Meunier, J.-R.; Averbeck, D.; Huang, M.-E. Selective cytotoxicity of Aniba rosaeodora essential oil towards epidermoid cancer cells through induction of apoptosis. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2011, 718, 24–32. [Google Scholar] [CrossRef]
- Ni, X.; Suhail, M.M.; Yang, Q.; Cao, A.; Fung, K.M.; Postier, R.G.; Woolley, C.; Young, G.; Zhang, J.; Lin, H.-K. Frankincense essential oil prepared from hydrodistillation of Boswellia sacra gum resins induces human pancreatic cancer cell death in cultures and in a xenograft murine model. BMC Complement. Altern. Med. 2012, 12, 253. [Google Scholar] [CrossRef] [Green Version]
- Rabi, T.; Bishayee, A. d-Limonene sensitizes docetaxel-induced cytotoxicity in human prostate cancer cells: Generation of reactive oxygen species and induction of apoptosis. J. Carcinog. 2009, 8, 9. [Google Scholar]
- Legault, J.; Pichette, A. Potentiating effect of β-caryophyllene on anticancer activity of α-humulene, isocaryophyllene and paclitaxel. J. Pharm. Pharmacol. 2007, 59, 1643–1647. [Google Scholar] [CrossRef]
- Lustberg, M.B. Management of neutropenia in cancer patients. Clin. Adv. Hematol. Oncol. HO 2012, 10, 825. [Google Scholar]
- Zhuang, S.R.; Chen, S.L.; Tsai, J.H.; Huang, C.C.; Wu, T.C.; Liu, W.S.; Tseng, H.C.; Lee, H.S.; Huang, M.C.; Shane, G.T.; et al. Effect of citronellol and the Chinese medical herb complex on cellular immunity of cancer patients receiving chemotherapy/radiotherapy. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2009, 23, 785–790. [Google Scholar] [CrossRef]
- Carnesecchi, S.; Bras-Gonçalves, R.; Bradaia, A.; Zeisel, M.; Gossé, F.; Poupon, M.F.; Raul, F. Geraniol, a component of plant essential oils, modulates DNA synthesis and potentiates 5-fluorouracil efficacy on human colon tumor xenografts. Cancer Lett. 2004, 215, 53–59. [Google Scholar] [CrossRef]
- Ainane, A.; Abdoul-Latif, F.M.; Abdoul-Latif, T.M.; Ainane, T. Evaluation of biological activities of two essential oils as a safe environmental bioinsecticides: Case of Eucalyptus globulus and Rosmarinus officinalis. Przegląd Nauk. Inżynieria I Kształtowanie Środowiska 2020, 29, 544–556. [Google Scholar] [CrossRef]
- Vieira, A.; Heidor, R.; Cardozo, M.T.; Scolastici, C.; Purgatto, E.; Shiga, T.M.; Barbisan, L.F.; Ong, T.P.; Moreno, F.S. Efficacy of geraniol but not of β-ionone or their combination for the chemoprevention of rat colon carcinogenesis. Braz. J. Med. Biol. Res. 2011, 44, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, J.K.; Gupta, S. Tocotrienol-rich fraction of palm oil induces cell cycle arrest and apoptosis selectively in human prostate cancer cells. Biochem. Biophys. Res. Commun. 2006, 346, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Karamysheva, A.F. Mechanisms of angiogenesis. Biochemistry 2008, 73, 751–762. [Google Scholar] [CrossRef]
- Janmey, P.A.; Kinnunen, P.K. Biophysical properties of lipids and dynamic membranes. Trends Cell Biol. 2006, 16, 538–546. [Google Scholar] [CrossRef]
- Ainane, A.; Abdoul-Latif, F.M.; Mohamed, J.; Attahar, W.; Ouassil, M.; Shybat, Z.L.; El Yaacoubi, A.; Ainane, T. Behaviour desorption study of the essential oil of Cedrus atlantica in a porous clay versus insecticidal activity against Sitophilus granarius: Explanation of the phenomenon by statistical studies. Int. J. Metrol. Qual. Eng. 2021, 12, 12. [Google Scholar] [CrossRef]
- Maddocks-Jennings, W.; Wilkinson, J.M. Aromatherapy practice in nursing: Literature review. J. Adv. Nurs. 2004, 48, 93–103. [Google Scholar] [CrossRef]
- Okigbo, R.N.; Anuagasi, C.L.; Amadi, J.E. Advances in selected medicinal and aromatic plants indigenous to Africa. J. Med. Plants Res. 2009, 3, 86–95. [Google Scholar]
- Talbi, M.; Saadali, B.; Boriky, D.; Bennani, L.; Elkouali, M.H.; Ainane, T. Two natural compounds—A benzofuran and a phenylpropane–from Artemisia dracunculus. J. Asian Nat. Prod. Res. 2016, 18, 724–729. [Google Scholar] [CrossRef]
- Spreckelmeyer, S.; Orvig, C.; Casini, A. Cellular transport mechanisms of cytotoxic metallodrugs: An overview beyond cisplatin. Molecules 2014, 19, 15584–15610. [Google Scholar] [CrossRef]
- Sullivan, L.B.; Chandel, N.S. Mitochondrial reactive oxygen species and cancer. Cancer Metab. 2014, 2, 17. [Google Scholar] [CrossRef] [PubMed]
- Jaafari, A.; Mouse, H.A.; M’Bark, L.A.; Tilaoui, M.; Elhansali, M.; Lepoivre, M.; Aboufatima, R.; Melhaoui, A.; Chait, A.; Zyad, A. Differential antitumor effect of essential oils and their major components of Thymus broussonettii: Relationship to cell cycle and apoptosis induction. Herba Pol. 2009, 55, 36–50. [Google Scholar]
- Gedara, S.R. Terpenoid content of the leaves of Thymus algeriensis Boiss. Mansoura J. Pharm. Sci. 2008, 24, 133–143. [Google Scholar]
- Andonova, T.; Muhovski, Y.; Apostolova, E.; Naimov, S.; Petkova, Z.; Teneva, O.; Dimitrova-Dyulgerova, I. Koelreuteria paniculata Seed Oil—A Rich Natural Source of Unsaturated Fatty Acids and Phytocompounds with DNA Protective Potential. Foods 2023, 12, 2230. [Google Scholar] [CrossRef] [PubMed]
- Zeytinoğlu, M.; Aydin, S.; Öztürk, Y.; Başer, K.H.C. Inhibitory effects of carvacrol on DMBA induced pulmonary tumorigenesis in rats. ACTA Pharm. Sci. 1998, 40, 93–98. [Google Scholar]
- Özkan, A.; Erdoğan, A. A comparative evaluation of antioxidant and anticancer activity of essential oil from Origanum onites (Lamiaceae) and its two major phenolic components. Turk. J. Biol. 2011, 35, 735–742. [Google Scholar] [CrossRef]
- Abusnina, A.; Alhosin, M.; Keravis, T.; Muller, C.D.; Fuhrmann, G.; Bronner, C.; Lugnier, C. Down-regulation of cyclic nucleotide phosphodiesterase PDE1A is the key event of p73 and UHRF1 deregulation in thymoquinone-induced acute lymphoblastic leukemia cell apoptosis. Cell. Signal. 2011, 23, 152–160. [Google Scholar] [CrossRef]
- Carnesecchi, S.; Langley, K.; Exinger, F.; Gosse, F.; Raul, F. Geraniol, a component of plant essential oils, sensitizes human colonic cancer cells to 5-fluorouracil treatment. J. Pharmacol. Exp. Ther. 2002, 301, 625–630. [Google Scholar] [CrossRef] [Green Version]
- Polo, M.P.; De Bravo, M.G. Effect of geraniol on fatty-acid and mevalonate metabolism in the human hepatoma cell line Hep G2. Biochem. Cell Biol. 2006, 84, 102–111. [Google Scholar] [CrossRef]
- Cardozo, M.T.; de Conti, A.; Ong, T.P.; Scolastici, C.; Purgatto, E.; Horst, M.A.; Bassoli, B.K.; Moreno, F.S. Chemopreventive effects of β-ionone and geraniol during rat hepatocarcinogenesis promotion: Distinct actions on cell proliferation, apoptosis, HMGCoA reductase, and RhoA. J. Nutr. Biochem. 2011, 22, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Chidambara, K.N.M.; Jayaprakasha, G.K.; Shivappa, M.; Mantur, B.S.P. Citrus monoterpenes: Potential source of phytochemicals for cancer prevention. In Emerging Trends in Dietary Components for Preventing and Combating Disease; ACS Publications: Columbus, OH, USA, 2012; Volume 1093, pp. 545–558. [Google Scholar]
- Elegbede, J.A.; Maltzman, T.H.; Elson, C.E.; Gould, M.N. Effects of anticarcinogenic monoterpenes on phase II hepatic metabolizing enzymes. Carcinogenesis 1993, 14, 1221–1223. [Google Scholar] [CrossRef] [PubMed]
- Pattanayak, M.; Seth, P.K.; Smita, S.; Gupta, S.K. Geraniol and limonene interaction with 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA) reductase for their role as cancer chemopreventive agents. J. Proteom. Bioinform. 2009, 2, 466–474. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Shuzo, O.; Li, Y.; Han, R. Effect of d-limonene, Salvia miltiorrhiza and turmeric derivatives on membrane association of Ras gene product and gap junction intercellular communication. Yao Xue Xue Bao=Acta Pharm. Sin. 1998, 33, 821–827. [Google Scholar]
- Usta, J.; Kreydiyyeh, S.; Knio, K.; Barnabe, P.; Bou-Moughlabay, Y.; Dagher, S. Linalool decreases HepG2 viability by inhibiting mitochondrial complexes I and II, increasing reactive oxygen species and decreasing ATP and GSH levels. Chem.-Biol. Interact. 2009, 180, 39–46. [Google Scholar] [CrossRef]
- Gu, Y.; Ting, Z.; Qiu, X.; Zhang, X.; Gan, X.; Fang, Y.; Xu, X.; Xu, R. Linalool preferentially induces robust apoptosis of a variety of leukemia cells via upregulating p53 and cyclin-dependent kinase inhibitors. Toxicology 2010, 268, 19–24. [Google Scholar] [CrossRef]
- Horváthová, E.; Sramková, M.; Lábaj, J.; Slamenová, D. Study of cytotoxic, genotoxic and DNA-protective effects of selected plant essential oils on human cells cultured in vitro. Neuro Endocrinol. Lett. 2006, 27, 44–47. [Google Scholar]
- Jaafari, A.; Tilaoui, M.; Mouse, H.A.; M’bark, L.A.; Aboufatima, R.; Chait, A.; Lepoivre, M.; Zyad, A. Comparative study of the antitumor effect of natural monoterpenes: Relationship to cell cycle analysis. Rev. Bras. Farmacogn. 2012, 22, 534–540. [Google Scholar] [CrossRef] [Green Version]
- Ferraz, R.P.; Bomfim, D.S.; Carvalho, N.C.; Soares, M.B.; da Silva, T.B.; Machado, W.J.; Prata, A.P.N.; Costa, E.V.; Moraes, V.R.S.; Nogueira, P.C.L.; et al. Cytotoxic effect of leaf essential oil of Lippia gracilis Schauer (Verbenaceae). Phytomedicine 2013, 20, 615–621. [Google Scholar] [CrossRef] [Green Version]
- Al-Shabanah, O.A.; Badary, O.A.; Nagi, M.N.; Al-Gharably, N.M.; Al-Rikabi, A.C.; Al-Bekairi, A.M. Thymoquinone protects against doxorubicin-induced cardiotoxicity without compromising its antitumor activity. J. Exp. Clin. Cancer Res. CR 1998, 17, 193–198. [Google Scholar] [PubMed]
- Effenberger-Neidnicht, K.; Schobert, R. Combinatorial effects of thymoquinone on the anti-cancer activity of doxorubicin. Cancer Chemother. Pharmacol. 2011, 67, 867–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dayem, A.A.; Choi, H.Y.; Kim, J.H.; Cho, S.G. Role of oxidative stress in stem, cancer, and cancer stem cells. Cancers 2010, 2, 859–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semlali, A.; Contant, C.; Al-Otaibi, B.; Al-Jammaz, I.; Chandad, F. The curcumin analog (PAC) suppressed cell survival and induced apoptosis and autophagy in oral cancer cells. Sci. Rep. 2021, 11, 11701. [Google Scholar] [CrossRef] [PubMed]
- Contant, C.; Rouabhia, M.; Loubaki, L.; Chandad, F.; Semlali, A. Anethole induces anti-oral cancer activity by triggering apoptosis, autophagy and oxidative stress and by modulation of multiple signaling pathways. Sci. Rep. 2021, 11, 13087. [Google Scholar] [CrossRef]
- Zhang, H.; Tikekar, R.V.; Ding, Q.; Gilbert, A.R.; Wimsatt, S.T. Inactivation of foodborne pathogens by the synergistic combinations of food processing technologies and food-grade compounds. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2110–2138. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Long, Y.; Li, D.; Shi, A.; Deng, J.; Ma, Y.; Wen, J.; Li, X.; Zhang, Y.; Liu, S.; et al. Natural essential oils efficacious in internal organs fibrosis treatment: Mechanisms of action and application perspectives. Pharmacol. Res. 2022, 182, 106339. [Google Scholar] [CrossRef]
- Das, I.; Acharya, A.; Berry, D.L.; Sen, S.; Williams, E.; Permaul, E.; Sengupta, A.; Bhattacharya, S.; Saha, T. Antioxidative effects of the spice cardamom against non-melanoma skin cancer by modulating nuclear factor erythroid-2-related factor 2 and NF-κB signalling pathways. Br. J. Nutr. 2012, 108, 984–997. [Google Scholar] [CrossRef] [Green Version]
- Pavithra, P.S.; Mehta, A.; Verma, R.S. Essential oils: From prevention to treatment of skin cancer. Drug Discov. Today 2019, 24, 644–655. [Google Scholar] [CrossRef]
- Islam, S.S.; Al-Sharif, I.; Sultan, A.; Al-Mazrou, A.; Remmal, A.; Aboussekhra, A. Eugenol potentiates cisplatin anti-cancer activity through inhibition of ALDH-positive breast cancer stem cells and the NF-κB signaling pathway. Mol. Carcinog. 2018, 57, 333–346. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Sailo, B.L.; Banik, K.; Harsha, C.; Prasad, S.; Gupta, S.C.; Bharti, A.C.; Aggarwal, B.B. Chronic diseases, inflammation, and spices: How are they linked? J. Transl. Med. 2018, 16, 14. [Google Scholar] [CrossRef] [Green Version]
- Attahar, W.; Mohamed Abdoul-Latif, F.; Mohamed, J.; Ainane, A.; Ainane, T. Antimicrobial and antioxidant activities of Trigonella foenum-graecum essential oil from the region of settat (Morocco). Pharmacologyonline 2021, 2, 434–442. [Google Scholar]
- Deb, D.D.; Parimala, G.; Devi, S.S.; Chakraborty, T. Effect of thymol on peripheral blood mononuclear cell PBMC and acute promyelotic cancer cell line HL-60. Chem.-Biol. Interact. 2011, 193, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Crowell, P.L. Monoterpenes in breast cancer chemoprevention. Breast Cancer Res. Treat. 1997, 46, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Giudice, A.; Montella, M. Activation of the Nrf2–ARE signaling pathway: A promising strategy in cancer prevention. Bioessays 2006, 28, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Jana, S.; Mandlekar, S. Role of phase II drug metabolizing enzymes in cancer chemoprevention. Curr. Drug Metab. 2009, 10, 595–616. [Google Scholar] [CrossRef]
- Kim, S.H.; Park, E.J.; Lee, C.R.; Chun, J.N.; Cho, N.H.; Kim, I.G.; Lee, S.; Kim, T.W.; Park, H.H.; So, I.; et al. Geraniol induces cooperative interaction of apoptosis and autophagy to elicit cell death in PC-3 prostate cancer cells. Int. J. Oncol. 2012, 40, 1683–1690. [Google Scholar]
- Ouassil, M.; Mohamed Abdoul-Latif, F.; Attahar, W.; Ainane, A.; Ainane, T. Chemical composition of bay laurel and rosemary essential oils from morocco and their antifungal activity against fusarium strains. Pharmacologyonline 2021, 2, 426–433. [Google Scholar]
- Kumar, S.; Mathew, S.O.; Aharwal, R.P.; Tulli, H.S.; Mohan, C.D.; Sethi, G.; Bishayee, A. Withaferin A: A Pleiotropic Anticancer Agent from the Indian Medicinal Plant Withania somnifera (L.) Dunal. Pharmaceuticals 2023, 16, 160. [Google Scholar] [CrossRef]
- Ainane, A.; Cherroud, S.; El Kouali, M.; Abba, E.H.; Ainane, T. Chemical compositions, insecticidal and antimicrobial activities of two moroccan essential oils of Citrus limonum and Syzygium aromaticum. Pharmacologyonline 2020, 2, 190–199. [Google Scholar]
- Lesgards, J.F.; Baldovini, N.; Vidal, N.; Pietri, S. Anticancer activities of essential oils constituents and synergy with conventional therapies: A review. Phytother. Res. 2014, 28, 1423–1446. [Google Scholar] [CrossRef] [PubMed]
- El Omari, N.; Bakrim, S.; Bakha, M.; Lorenzo, J.M.; Rebezov, M.; Shariati, M.A.; Aboulaghras, S.; Balahbib, A.; Khayrullin, M.; Bouyahya, A. Natural bioactive compounds targeting epigenetic pathways in cancer: A review on alkaloids, terpenoids, quinones, and isothiocyanates. Nutrients 2021, 13, 3714. [Google Scholar] [CrossRef]
- Rigo, A.; Ferrarini, I.; Lorenzetto, E.; Darra, E.; Liparulo, I.; Bergamini, C.; Sissa, C.; Cavalieri, E.; Vinante, F. BID and the α-bisabolol-triggered cell death program: Converging on mitochondria and lysosomes. Cell Death Dis. 2019, 10, 889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, N.; Zhu, K.; Czarnecka-Herok, J.; Vernier, M.; Bernard, D. Regulation and role of calcium in cellular senescence. Cell Calcium 2023, 110, 102701. [Google Scholar] [CrossRef]
- Tsujimoto, Y.; Shimizu, S. Another way to die: Autophagic programmed cell death. Cell Death Differ. 2005, 12, 1528–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvitz, H.R. Genetic control of programmed cell death in the nematode Caenorhabditis elegans. Cancer Res. 1999, 59 (Suppl. S7), 1701s–1706s. [Google Scholar]
- Fearnhead, H.O.; Dinsdale, D.; Cohen, G.M. An interleukin-1β-converting enzyme-like protease is a common mediator of apoptosis in thymocytes. FEBS Lett. 1995, 375, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Tsuruo, T.; Naito, M.; Tomida, A.; Fujita, N.; Mashima, T.; Sakamoto, H.; Haga, N. Molecular targeting therapy of cancer: Drug resistance, apoptosis and survival signal. Cancer Sci. 2003, 94, 15–21. [Google Scholar] [CrossRef]
- Vara, J.Á.F.; Casado, E.; de Castro, J.; Cejas, P.; Belda-Iniesta, C.; González-Barón, M. PI3K/Akt signalling pathway and cancer. Cancer Treat. Rev. 2004, 30, 193–204. [Google Scholar] [CrossRef]
- Bouyahya, A.; Belmehdi, O.; Benjouad, A.; El Hassani, R.A.; Amzazi, S.; Dakka, N.; Bakri, Y. Pharmacological properties and mechanism insights of Moroccan anticancer medicinal plants: What are the next steps? Ind. Crops Prod. 2020, 147, 112198. [Google Scholar] [CrossRef]
- Khoo, B.Y.; Chua, S.L.; Balaram, P. Apoptotic effects of chrysin in human cancer cell lines. Int. J. Mol. Sci. 2010, 11, 2188–2199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ainane, T.; Abdoul-Latif, F.M.; Shybat, Z.L.; Mohamed, J.; Ainane, A. Antifungal activity of essential oil of Pistacia atlantica against Ascochyta rabiei and its correlation with antioxidant activity. Pharmacologyonline 2021, 3, 829–837. [Google Scholar]
- Yin, Q.H.; Yan, F.X.; Zu, X.Y.; Wu, Y.H.; Wu, X.P.; Liao, M.C.; Deng, S.-W.; Yin, L.-L.; Zhuang, Y.-Z. Anti-proliferative and pro-apoptotic effect of carvacrol on human hepatocellular carcinoma cell line HepG-2. Cytotechnology 2012, 64, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Rajput, S.; Kumar, B.P.; Dey, K.K.; Pal, I.; Parekh, A.; Mandal, M. Molecular targeting of Akt by thymoquinone promotes G1 arrest through translation inhibition of cyclin D1 and induces apoptosis in breast cancer cells. Life Sci. 2013, 93, 783–790. [Google Scholar] [CrossRef]
- Tundis, R.; Loizzo, M.R.; Bonesi, M.; Menichini, F.; Dodaro, D.; Passalacqua, N.G.; Statti, G.; Menichini, F. In vitro cytotoxic effects of Senecio stabianus Lacaita (Asteraceae) on human cancer cell lines. Nat. Prod. Res. 2009, 23, 1707–1718. [Google Scholar] [CrossRef]
- Banerjee, S.; Azmi, A.S.; Padhye, S.; Singh, M.W.; Baruah, J.B.; Philip, P.A.; Sarkar, F.H.; Mohammad, R.M. Structure-activity studies on therapeutic potential of Thymoquinone analogs in pancreatic cancer. Pharm. Res. 2010, 27, 1146–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sethi, G.; Ahn, K.S.; Aggarwal, B.B. Targeting nuclear factor-κB activation pathway by thymoquinone: Role in suppression of antiapoptotic gene products and enhancement of apoptosis. Mol. Cancer Res. 2008, 6, 1059–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazan, L.S.; Ng, W.K.; Al-Naqeeb, G.; Ismail, M. Cytotoxicity of thymoquinone (TQ) from Nigella sativa towards human cervical carcinoma cells (HeLa). J. Pharm. Res. 2009, 2, 585–589. [Google Scholar]
- Zhang, X.J.; Jia, S.S. Fisetin inhibits laryngeal carcinoma through regulation of AKT/NF-κB/mTOR and ERK1/2 signaling pathways. Biomed. Pharmacother. 2016, 83, 1164–1174. [Google Scholar] [CrossRef] [PubMed]
- Martelli, A.M.; Nyåkern, M.; Tabellini, G.; Bortul, R.; Tazzari, P.L.; Evangelisti, C.; Cocco, L. Phosphoinositide 3-kinase/Akt signaling pathway and its therapeutical implications for human acute myeloid leukemia. Leukemia 2006, 20, 911–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frauwirth, K.A.; Riley, J.L.; Harris, M.H.; Parry, R.V.; Rathmell, J.C.; Plas, D.R.; Elstrom, R.L.; June, C.H.; Thompson, C.B. The CD28 signaling pathway regulates glucose metabolism. Immunity 2002, 16, 769–777. [Google Scholar] [CrossRef] [Green Version]
- Monirujjaman, M.D.; Ferdouse, A. Metabolic and physiological roles of branched-chain amino acids. Adv. Mol. Biol. 2014, 2014, 364976. [Google Scholar] [CrossRef] [Green Version]
- Patra, K.C.; Hay, N. The pentose phosphate pathway and cancer. Trends Biochem. Sci. 2014, 39, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Abdoul-Latif, F.M.; Ainane, A.; Oumaskour, K.; Boujaber, N.; Mohamed, J.; Ainane, T. Effect of atlas cedar essential oil against pathogenic medical bacterial strains-in vitro test. Pharmacologyonline 2021, 3, 857–864. [Google Scholar]
- Abildgaard, C.; Guldberg, P. Molecular drivers of cellular metabolic reprogramming in melanoma. Trends Mol. Med. 2015, 21, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Wang, Z.; Sebastian Yakisich, J. Natural products targeting autophagy via the PI3K/Akt/mTOR pathway as anticancer agents. Anti-Cancer Agents Med. Chem. (Former. Curr. Med. Chem.-Anti-Cancer Agents) 2013, 13, 1048–1056. [Google Scholar] [CrossRef]
- Abdoul-Latif, F.M.; Ainane, A.; Oumaskour, K.; Boujaber, N.; Mohamed, J.; Ainane, T. In vitro antidiabetic activity of essential oil of two species of Artemisia: Artemisia heba-alba asso and Artemisia ifranensis. Pharmacologyonline 2021, 3, 812–820. [Google Scholar]
- Ramamurthy, S.; Ronnett, G.V. Developing a head for energy sensing: AMP-activated protein kinase as a multifunctional metabolic sensor in the brain. J. Physiol. 2006, 574, 85–93. [Google Scholar] [CrossRef]
- Chartoumpekis, D.V.; Kensler, T.W. New player on an old field; the keap1/Nrf2 pathway as a target for treatment of type 2 diabetes and metabolic syndrome. Curr. Diabetes Rev. 2013, 9, 137–145. [Google Scholar] [CrossRef]
- Ribas, V.; García-Ruiz, C.; Fernández-Checa, J.C. Mitochondria, cholesterol and cancer cell metabolism. Clin. Transl. Med. 2016, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Tomko, A.M.; Whynot, E.G.; Ellis, L.D.; Dupré, D.J. Anti-cancer potential of cannabinoids, terpenes, and flavonoids present in cannabis. Cancers 2020, 12, 1985. [Google Scholar] [CrossRef]
- Ballou, L.R.; Laulederkind, S.J.; Rosloniec, E.F.; Raghow, R. Ceramide signalling and the immune response. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metab. 1996, 1301, 273–287. [Google Scholar] [CrossRef]
- Ainane, T.; Abourriche, A.; Kabbaj, M.; Elkouali, M.; Bennamara, A.; Charrouf, M.; Lemrani, M. Biological activities of extracts from seaweed Cystoseira tamariscifolia: Antibacterial activity, antileishmanial activity and cytotoxicity. J. Chem. Pharm. Res. 2014, 6, 607–611. [Google Scholar]
- Maltese, W.A.; Defendini, R.; Green, R.A.; Sheridan, K.M.; Donley, D.K. Suppression of murine neuroblastoma growth in vivo by mevinolin, a competitive inhibitor of 3-hydroxy-3-methylglutaryl-coenzyme A reductase. J. Clin. Investig. 1985, 76, 1748–1754. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.S.; Goldstein, J.L. Multivalent feedback regulation of HMG CoA reductase, a control mechanism coordinating isoprenoid synthesis and cell growth. J. Lipid Res. 1980, 21, 505–517. [Google Scholar] [CrossRef]
- Karabín, M.; Hudcová, T.; Jelínek, L.; Dostálek, P. Biologically active compounds from hops and prospects for their use. Compr. Rev. Food Sci. Food Saf. 2016, 15, 542–567. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, J.; Sureda, A.; Tenore, G.C.; Daglia, M.; Sharifi-Rad, M.; Valussi, M.; Tundis, R.; Sharifi-Rad, M.; Loizzo, M.R.; Ademiluyi, A.O.; et al. Biological activities of essential oils: From plant chemoecology to traditional healing systems. Molecules 2017, 22, 70. [Google Scholar] [CrossRef]
- Kumar, M.; Tomar, M.; Amarowicz, R.; Saurabh, V.; Nair, M.S.; Maheshwari, C.; Sasi, M.; Prajapati, U.; Hasan, M.; Singh, S.; et al. Guava (Psidium guajava L.) leaves: Nutritional composition, phytochemical profile, and health-promoting bioactivities. Foods 2021, 10, 752. [Google Scholar] [CrossRef]
- Fenley, A.T.; Anandakrishnan, R.; Kidane, Y.H.; Onufriev, A.V. Modulation of nucleosomal DNA accessibility via charge-altering post-translational modifications in histone core. Epigenetics Chromatin 2018, 11, 11. [Google Scholar] [CrossRef]
- Bowman, G.D.; Poirier, M.G. Post-translational modifications of histones that influence nucleosome dynamics. Chem. Rev. 2014, 115, 2274–2295. [Google Scholar] [CrossRef] [Green Version]
- Mariño-Ramírez, L.; Kann, M.G.; Shoemaker, B.A.; Landsman, D. Histone structure and nucleosome stability. Expert Rev. Proteom. 2005, 2, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.R.; Schulze, A. Lipid metabolism in cancer. FEBS J. 2012, 279, 2610–2623. [Google Scholar] [CrossRef] [PubMed]
- Alam, A.; Jawaid, T.; Alsanad, S.M.; Kamal, M.; Balaha, M.F. Composition, Antibacterial Efficacy, and Anticancer Activity of Essential Oil Extracted from Psidium guajava (L.) Leaves. Plants 2023, 12, 246. [Google Scholar] [CrossRef] [PubMed]
- Chinni, S.R.; Li, Y.; Upadhyay, S.; Koppolu, P.K.; Sarkar, F.H. Indole-3-carbinol (I3C) induced cell growth inhibition, G1 cell cycle arrest and apoptosis in prostate cancer cells. Oncogene 2001, 20, 2927–2936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.C.S.; Ko, P.J.; Pan, Y.S.; Lin, H.Y.; Whang-Peng, J.; Davis, P.J.; Wang, K. Role of thyroid hormone-integrin αvβ3-signal and therapeutic strategies in colorectal cancers. J. Biomed. Sci. 2021, 28, 24. [Google Scholar] [CrossRef]
- Di Martile, M.; Garzoli, S.; Ragno, R.; Del Bufalo, D. Essential oils and their main chemical components: The past 20 years of preclinical studies in melanoma. Cancers 2020, 12, 2650. [Google Scholar] [CrossRef] [PubMed]
- Santoro, A.; Zhao, J.; Wu, L.; Carru, C.; Biagi, E.; Franceschi, C. Microbiomes other than the gut: Inflammaging and age-related diseases. In Seminars in Immunopathology; Springer: Berlin/Heidelberg, Germany, 2020; Volume 42, pp. 589–605. [Google Scholar]
- Gurib-Fakim, A. Medicinal plants: Traditions of yesterday and drugs of tomorrow. Mol. Asp. Med. 2006, 27, 1–93. [Google Scholar] [CrossRef]
- Tajana, A. Stereoselective Pharmacokinetics. In New Trends in Pharmacokinetics; NATO ASI SERIES; Springer: Berlin/Heidelberg, Germany, 1991; pp. 227–255. [Google Scholar]
- Labiris, N.R.; Dolovich, M.B. Pulmonary drug delivery. Part I: Physiological factors affecting therapeutic effectiveness of aerosolized medications. Br. J. Clin. Pharmacol. 2003, 56, 588–599. [Google Scholar] [CrossRef]
- Menn, J.J. Comparative aspects of pesticide metabolism in plants and animals. Environ. Health Perspect. 1978, 27, 113–124. [Google Scholar] [CrossRef]
- Singletary, K. Oregano: Overview of the literature on health benefits. Nutr. Today 2010, 45, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Evangelista, L.; Urso, L.; Caracciolo, M.; Stracuzzi, F.; Panareo, S.; Cistaro, A.; Catalano, O. Fdg pet/ct volume-based quantitative data and survival analysis in breast cancer patients: A systematic review of the literature. Curr. Med. Imaging 2023, 19, 807–816. [Google Scholar] [CrossRef] [PubMed]
- Pati, S.; Irfan, W.; Jameel, A.; Ahmed, S.; Shahid, R.K. Obesity and cancer: A current overview of epidemiology, pathogenesis, outcomes, and management. Cancers 2023, 15, 485. [Google Scholar] [CrossRef]
- Api, A.M.; Ritacco, G.; Hawkins, D.R. The fate of dermally applied [14C] d-limonene in rats and humans. Int. J. Toxicol. 2013, 32, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Hegeman, G.D. Synthesis of the enzymes of the mandelate pathway by Pseudomonas putida I. Synthesis of enzymes by the wild type. J. Bacteriol. 1966, 91, 1140–1154. [Google Scholar] [CrossRef] [PubMed]
- Salgueiro, L.; Martins, A.P.; Correia, H. Raw materials: The importance of quality and safety. A review. Flavour Fragr. J. 2010, 25, 253–271. [Google Scholar] [CrossRef]
- Sinha, A.K.; Sharma, U.K.; Sharma, N. A comprehensive review on vanilla flavor: Extraction, isolation and quantification of vanillin and others constituents. Int. J. Food Sci. Nutr. 2008, 59, 299–326. [Google Scholar] [CrossRef]
- Valaes, T. Severe neonatal jaundice associated with glucose-6-phosphate dehydrogenase deficiency: Pathogenesis and global epidemiology. Acta Paediatr. 1994, 83, 58–76. [Google Scholar] [CrossRef]
- Fenichel, G.M. Clinical Pediatric Neurology: A Signs and Symptoms Approach; Elsevier Health Sciences: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Alvarez-Diez, T.M.; Zheng, J. Mechanism-based inactivation of cytochrome P450 3A4 by 4-ipomeanol. Chem. Res. Toxicol. 2004, 17, 150–157. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Family | Main Components | References |
|---|---|---|
| Cupressaceae | β-pinene; α-pinene. | [60] |
| Cupressaceae | D-verbenol; Cedrol; L-verbenol. | [61] |
| Cupressaceae | Camphor; Bornyl d’acetate. | [62] |
| Euphorbiaceae | Viridiflorene. | [63] |
| Flacourtiaceae | β-caryophyllene; α-humulene. | [64] |
| Lamiaceae | 1,8-cineole; | [65,66,67,68] |
| Borneol; | ||
| Camphor; | ||
| Carvacrol; | ||
| D-germacrene; | ||
| Limonene; | ||
| p-cymene; | ||
| Thymol; | ||
| α-thujone; | ||
| γ-terpinene. | ||
| Meliaceae | Caryophyllene; β-caryophyllene. | [69] |
| Myricaceae | β-caryophyllene; α-humulene. | [70] |
| Myrtaceae | α-pinene; β-caryophyllene. | [71] |
| Rosaceae | α-pinene; Sabinene; γ-eudesmol; α-thujene; α-humulene. | [72] |
| Rutaceae | β-caryophyllene; β-elemene. | [73] |
| Species | Major EO Constituent(s) | Cancer Cell Lines | Noncancer Cell Lines | Major Findings and EO Concentrations | Mechanisms | Reference |
|---|---|---|---|---|---|---|
| Thymus fallax | Carvacrol, p-cymene, thymol, γ-terpinene | DLD-1 (CRc) | Mouse fibroblast (L.929) | Cytotoxic to cancer cells (IC50 0.347 mg/mL) and noncytotoxic to normal cells (IC50 22 mg/mL) | Antioxidant activity | [93] |
| Boswellia sacra | α-pinene, α-thujene, β-pinene, myrcene, boswellic acid | T47D, MCF7, MDA-MB-231 (Bc) | Immortalized normal human breast (MCF10-2A) | Cytotoxic to cancer cells (EO dilution IC50 1:900 for TD47, 1:1000 for MCF7, 1:950 for MDA-MB-231) and noncytotoxic to immortalized normal cells (EO dilution IC50 1:680) | Antiproliferative | [77] |
| Amomum tsaoko | 1,8-cineole, ρ-propylbenzaldehyde, geraniol, geranial, α-terpineol, α-phellandrene, neral, β-pinene | HepG2 and Bel-7402 (Lc) HeLa (Cc), A549 (Lc), SGC-7901 (GAC), PC-3 (Pc) | Hepatocyte (HL-7702) and umbilical vein endothelial (HUVEC) | Cytotoxic to cancer cells, particularly HepG2 (IC50 31.8 μg/mL), Hela (IC50 66.46 μg/mL) and Bel-7402 (IC50 96.08 μg/mL), with less cytotoxicity towards HL-7702 (IC50 272.4 μg/mL) and HUVEC (IC50 163.91 μg/mL). No cytotoxicity towards A549 | Antiproliferative | [94] |
| Lippia alba (Citral chemotype) | Geranial, neral, geraniol, trans-β-caryophyllene, 6-methyl-5-hepten-2-one, limonene, linalool | HeLa (Cc) | African green monkey kidney (Vero) | Cytotoxic to cancer cells (CC50 3.5 μg/mL) and noncytotoxic to normal cells (CC50 > 100 μg/mL) | Citral-dependent cytotoxicity | [95] |
| Boswellia sp. (1200 mg/mL frankincense gum resin) | Duva-3,9,13-trien-1,5alpha-diol-1-acetate, octyl acetate, o-methyl anisole, naphthalene decahydro-1,1,4a-trimethyl-6-methylene-5-(3-methyl-2-pentenyl), thunbergol (Mikhaeil et al., 2003) | J82 (Blc) | Human urothelium (UROtsa) | Cytotoxic to cancer cells (no viable cells after EO dilution 1:1100 after 24 h) and noncytotoxic to normal cells (no viable cells after EO dilution 1:400) | Antiproliferative | [96] |
| Casearia sylvestris | Bicyclogermacrene, β-caryophyllene, spathulenol, α-humulene, α-pinene | HeLa (Cc), A549 (Lc) HT-29 (CRc) | Monkey kidney (Vero) and mice macrophages | Cytotoxic to HeLa (CD50 63.3 µg·ml−1), A549 (CD50 60.7 µg·ml−1) and HT-29 (CD50 90.6 µg·ml−1) with less cytotoxicity to Vero (CD50 210.1 µg·ml−1) and macrophages (CD50 234.0 µg·ml−1) | Cytotoxicity | [97] |
| Zanthoxylum rhoifolum Lam | ß-caryophyllene, α-humulene, α-pinene, myrcene, linalool | HeLa (Cc), A549 (Lc) HT-29 (CRc) | Monkey kidney (Vero) and mice macrophages | Cytotoxic to HeLa (CD50 90.7 µg/mL), A549 (CD50 82.3 µg/mL), and HT-29 (CD50 113.6 µg/mL) and noncytotoxic to normal cells (CD50 > 600 µg/mL) | Cytotoxicity | [98] |
| Commiphora gileadensis | Sabinene, ß-caryophyllene, germacrene D, α-pinene | BS-241 (Mouse T-cell lymphoma) MoFir (Epstein Barr virus transformed human B lymphocytes) | Normal human skin fibroblasts (FB) | EO dilution of 1:5000 killed 87% of BS-24-1 cells and 40% of MoFir cells | Antiproliferative | [99] |
| Aniba rosaeodora | Rosewood essential oil (REO), linalool | A431 (Ec), HaCaT (pre-cancerous) | Epidermal keratinocytes (HEK001, NHEK) | Cytotoxicity to cancer cells A431 and HaCaT (<20% viability) and minor cytotoxicity to normal cells HEK001 and NHEK (>70% viability) | Cytotoxicity | [100] |
| Cell Lines | Chemotherapy Drug Used Alone and Concentration | EO Constituent Used Alone and Concentration | Combined EO and Chemotherapy Drug | Reference | |
|---|---|---|---|---|---|
| Prostate cancer cell (DU-145) | Docetaxel IC50 2.8 nM | d-limonene IC50 2.8 mM | IC50 docetaxel 1.9 mM and d-limonene 0.2 mM | [102] | |
| Human breast cancer (MCF-7) | Paclitaxel 0.025 µg/mL−1 resulted in 28% cell growth inhibition | β-caryophyllene resulted in no inhibition of cell growth | β-caryophyllene 2.5 µg/mL−1 and Paclitaxel 0.025 µg/mL−1 resulted in 50% cell growth inhibition | β-caryophyllene 10 µg/mL−1 and Paclitaxel 0.025 µg/mL−1 resulted in 68% cell growth inhibition | [103] |
| Human colorectal adenocarcinoma (DLD-1) | Paclitaxel 0.025 µg/mL−1 resulted in 17.3% cell growth inhibition | β-caryophyllene resulted in no inhibition of cell growth | β-caryophyllene 2.5 µg/mL−1 and Paclitaxel 0.025 µg/mL−1 resulted in 91% cell growth inhibition | β-caryophyllene 10 µg/mL−1 and Paclitaxel 0.025 µg/mL−1 resulted in 189% cell growth inhibition | [103] |
| Mouse fibroblast (L-929) | Paclitaxel 0.025 µg/mL−1 resulted in 18.4% cell growth inhibition | β-caryophyllene resulted in no inhibition of cell growth | β-caryophyllene 2.5 µg/mL−1 and Paclitaxel 0.025 µg/mL−1 resulted in 36% cell growth inhibition | β-caryophyllene 10 µg/mL−1 and Paclitaxel 0.025 µg/mL−1 resulted in 123% cell growth inhibition | [103] |
| Components | Antitumor Activity | Animal and/or Cell Line Tested | References |
|---|---|---|---|
| Carvacrol |
|
| [118,119,120,121,122] |
| Geraniol |
|
| [123,124,125,126] |
| Limonene |
|
| [127,128,129,130] |
| Linalool |
|
| [131,132] |
| Thymol |
|
| [133,134,135] |
| Thymoquinone |
|
| [136,137] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamed Abdoul-Latif, F.; Ainane, A.; Houmed Aboubaker, I.; Mohamed, J.; Ainane, T. Exploring the Potent Anticancer Activity of Essential Oils and Their Bioactive Compounds: Mechanisms and Prospects for Future Cancer Therapy. Pharmaceuticals 2023, 16, 1086. https://doi.org/10.3390/ph16081086
Mohamed Abdoul-Latif F, Ainane A, Houmed Aboubaker I, Mohamed J, Ainane T. Exploring the Potent Anticancer Activity of Essential Oils and Their Bioactive Compounds: Mechanisms and Prospects for Future Cancer Therapy. Pharmaceuticals. 2023; 16(8):1086. https://doi.org/10.3390/ph16081086
Chicago/Turabian StyleMohamed Abdoul-Latif, Fatouma, Ayoub Ainane, Ibrahim Houmed Aboubaker, Jalludin Mohamed, and Tarik Ainane. 2023. "Exploring the Potent Anticancer Activity of Essential Oils and Their Bioactive Compounds: Mechanisms and Prospects for Future Cancer Therapy" Pharmaceuticals 16, no. 8: 1086. https://doi.org/10.3390/ph16081086