
Blocking the CGRP Receptor: Differences across Human Vascular Beds
 , and
, and
Abstract
:1. Introduction
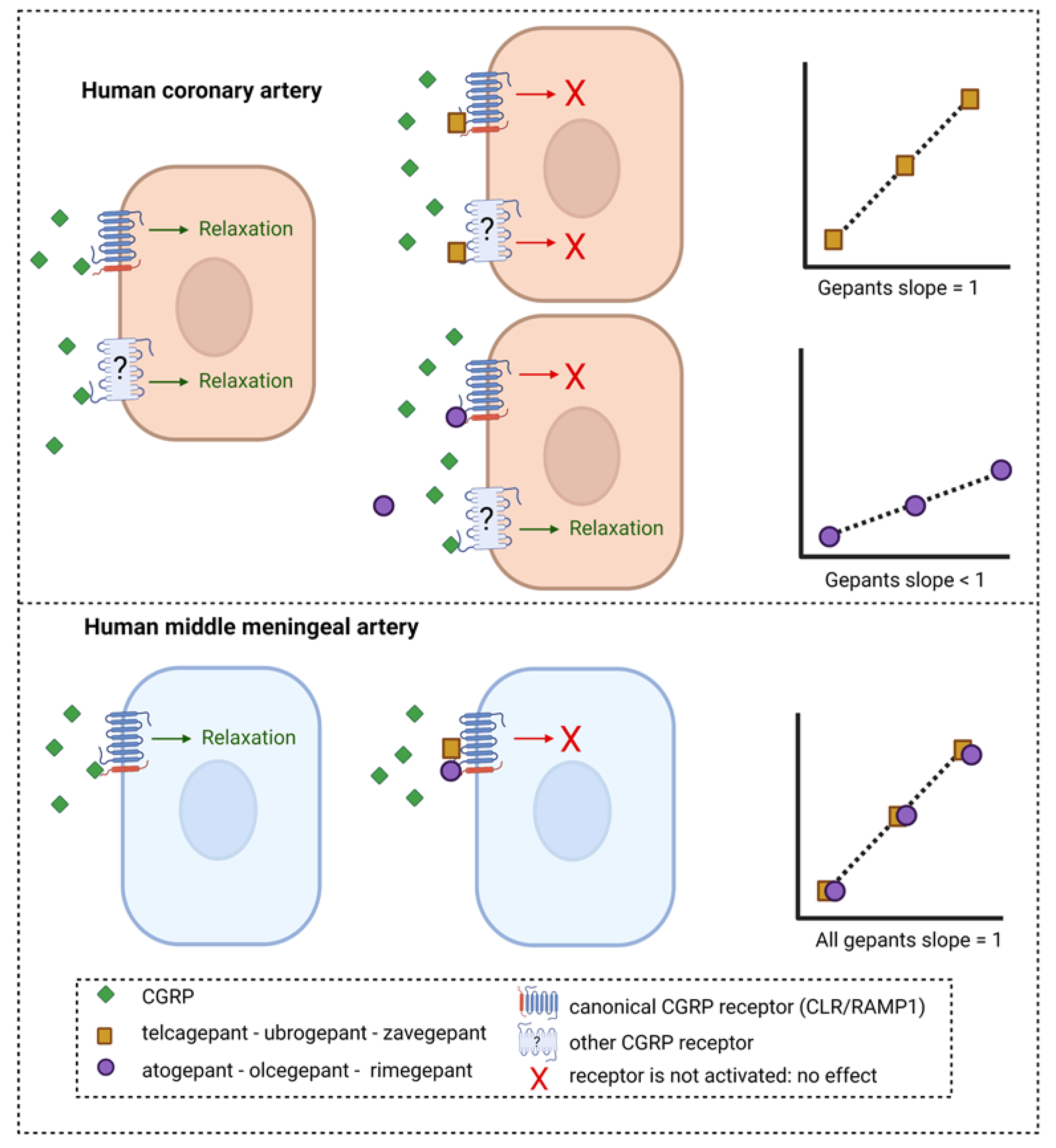
2. Results
2.1. Potency of Zavegepant in Human Coronary Arteries
2.2. Potency of All Gepants in Human Coronary Arteries and Human Middle Meningeal Arteries
3. Discussion
4. Materials and Methods
4.1. Wire Myography Experiments in Human Coronary Arteries
4.2. Comparison of Potency of Gepants in Human Coronary Arteries and Human Middle Meningeal Arteries
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smillie, S.J.; King, R.; Kodji, X.; Outzen, E.; Pozsgai, G.; Fernandes, E.; Marshall, N.; de Winter, P.; Heads, R.J.; Dessapt-Baradez, C.; et al. An ongoing role of alpha-calcitonin gene-related peptide as part of a protective network against hypertension, vascular hypertrophy, and oxidative stress. Hypertension 2014, 63, 1056–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MaassenVanDenBrink, A.; Meijer, J.; Villalon, C.M.; Ferrari, M.D. Wiping Out CGRP: Potential Cardiovascular Risks. Trends Pharmacol. Sci. 2016, 37, 779–788. [Google Scholar] [CrossRef] [PubMed]
- Noseda, R.; Burstein, R. Migraine pathophysiology: Anatomy of the trigeminovascular pathway and associated neurological symptoms, CSD, sensitization and modulation of pain. Pain 2013, 154, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Villalón, C.M.; Olesen, J. The role of CGRP in the pathophysiology of migraine and efficacy of CGRP receptor antagonists as acute antimigraine drugs. Pharmacol. Ther. 2009, 124, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Goadsby, P.J.; Edvinsson, L.; Ekman, R. Vasoactive peptide release in the extracerebral circulation of humans during migraine headache. Ann. Neurol. 1990, 28, 183–187. [Google Scholar] [CrossRef]
- Goadsby, P.J.; Edvinsson, L. The trigeminovascular system and migraine: Studies characterizing cerebrovascular and neuropeptide changes seen in humans and cats. Ann. Neurol. 1993, 33, 48–56. [Google Scholar] [CrossRef]
- Lassen, L.H.; Haderslev, P.A.; Jacobsen, V.B.; Iversen, H.K.; Sperling, B.; Olesen, J. CGRP may play a causative role in migraine. Cephalalgia Int. J. Headache 2002, 22, 54–61. [Google Scholar] [CrossRef]
- Hansen, J.M.; Hauge, A.W.; Olesen, J.; Ashina, M. Calcitonin gene-related peptide triggers migraine-like attacks in patients with migraine with aura. Cephalalgia Int. J. Headache 2010, 30, 1179–1186. [Google Scholar] [CrossRef]
- Hay, D.L.; Garelja, M.L.; Poyner, D.R.; Walker, C.S. Update on the pharmacology of calcitonin/CGRP family of peptides: IUPHAR Review 25. Br. J. Pharmacol. 2018, 175, 3–17. [Google Scholar] [CrossRef]
- Pak, K.; Kim, J.; Lee, G.H.; Cho, J.W.; Kim, H.W. Effectiveness of Calcitonin Gene-Related Peptide Receptor Antagonists for Migraine Treatment: A Meta-Analysis. Eur. Neurol. 2022, 85, 195–201. [Google Scholar] [CrossRef]
- Hong, P.; Tan, T.; Liu, Y.; Xiao, J. Gepants for abortive treatment of migraine: A network meta-analysis. Brain Behav. 2020, 10, e01701. [Google Scholar] [CrossRef]
- Lipton, R.B.; Croop, R.; Stock, D.A.; Madonia, J.; Forshaw, M.; Lovegren, M.; Mosher, L.; Coric, V.; Goadsby, P.J. Safety, tolerability, and efficacy of zavegepant 10 mg nasal spray for the acute treatment of migraine in the USA: A phase 3, double-blind, randomised, placebo-controlled multicentre trial. Lancet Neurol. 2023, 22, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Ashina, M.; Tepper, S.J.; Reuter, U.; Blumenfeld, A.M.; Hutchinson, S.; Xia, J.; Miceli, R.; Severt, L.; Finnegan, M.; Trugman, J.M. Once-daily oral atogepant for the long-term preventive treatment of migraine: Findings from a multicenter, randomized, open-label, phase 3 trial. Headache 2023, 63, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.W.; Connor, K.M.; Zhang, Y.; Pearlman, E.; Koppenhaver, J.; Fan, X.; Lines, C.; Edvinsson, L.; Goadsby, P.J.; Michelson, D. Randomized controlled trial of the CGRP receptor antagonist telcagepant for migraine prevention. Neurology 2014, 83, 958–966. [Google Scholar] [CrossRef] [PubMed]
- Woodhead, J.L.; Siler, S.Q.; Howell, B.A.; Watkins, P.B.; Conway, C. Comparing the Liver Safety Profiles of 4 Next-Generation CGRP Receptor Antagonists to the Hepatotoxic CGRP Inhibitor Telcagepant Using Quantitative Systems Toxicology Modeling. Toxicol. Sci. 2022, 188, 108–116. [Google Scholar] [CrossRef]
- Haghdoost, F.; Puledda, F.; Garcia-Azorin, D.; Huessler, E.M.; Messina, R.; Pozo-Rosich, P. Evaluating the efficacy of CGRP mAbs and gepants for the preventive treatment of migraine: A systematic review and network meta-analysis of phase 3 randomised controlled trials. Cephalalgia Int. J. Headache 2023, 43, 03331024231159366. [Google Scholar] [CrossRef]
- Gupta, S.; Mehrotra, S.; Villalón, C.M.; Garrelds, I.M.; de Vries, R.; van Kats, J.P.; Sharma, H.S.; Saxena, P.R.; Maassenvandenbrink, A. Characterisation of CGRP receptors in human and porcine isolated coronary arteries: Evidence for CGRP receptor heterogeneity. Eur. J. Pharmacol. 2006, 530, 107–116. [Google Scholar] [CrossRef]
- Gupta, S.; Mehrotra, S.; Avezaat, C.J.; Villalón, C.M.; Saxena, P.R.; Maassenvandenbrink, A. Characterisation of CGRP receptors in the human isolated middle meningeal artery. Life Sci. 2006, 79, 265–271. [Google Scholar] [CrossRef]
- Chan, K.Y.; Edvinsson, L.; Eftekhari, S.; Kimblad, P.O.; Kane, S.A.; Lynch, J.; Hargreaves, R.J.; de Vries, R.; Garrelds, I.M.; van den Bogaerdt, A.J.; et al. Characterization of the calcitonin gene-related peptide receptor antagonist telcagepant (MK-0974) in human isolated coronary arteries. J. Pharmacol. Exp. Ther. 2010, 334, 746–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edvinsson, L.; Chan, K.Y.; Eftekhari, S.; Nilsson, E.; de Vries, R.; Saveland, H.; Dirven, C.M.; Danser, A.H.; MaassenVanDenBrink, A. Effect of the calcitonin gene-related peptide (CGRP) receptor antagonist telcagepant in human cranial arteries. Cephalalgia Int. J. Headache 2010, 30, 1233–1240. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Beltrán, E.; Chan, K.Y.; Danser, A.H.J.; MaassenVanDenBrink, A.; Edvinsson, L. Characterisation of the calcitonin gene-related peptide receptor antagonists ubrogepant and atogepant in human isolated coronary, cerebral and middle meningeal arteries. Cephalalgia Int. J. Headache 2019, 40, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Mulder, I.A.; Li, M.; de Vries, T.; Qin, T.; Yanagisawa, T.; Sugimoto, K.; van den Bogaerdt, A.; Danser, A.H.J.; Wermer, M.J.H.; van den Maagdenberg, A.; et al. Anti-migraine Calcitonin Gene-Related Peptide Receptor Antagonists Worsen Cerebral Ischemic Outcome in Mice. Ann. Neurol. 2020, 88, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Boucherie, D.M.; Dammers, R.; Vincent, A.; Danser, A.H.J.; MaassenVanDenBrink, A. Comparison of gepant effects at therapeutic plasma concentrations: Connecting pharmacodynamics and pharmacokinetics. Cephalalgia, in Press.
- Kenakin, T.P. The Schild regression in the process of receptor classification. Can. J. Physiol. Pharmacol. 1982, 60, 249–265. [Google Scholar] [CrossRef]
- Vauquelin, G.; Van Liefde, I.; Swinney, D.C. On the different experimental manifestations of two-state ‘induced-fit’ binding of drugs to their cellular targets. Br. J. Pharmacol. 2016, 173, 1268–1285. [Google Scholar] [CrossRef] [Green Version]
- Pan, K.S.; Siow, A.; Hay, D.L.; Walker, C.S. Antagonism of CGRP Signaling by Rimegepant at Two Receptors. Front. Pharmacol. 2020, 11, 1240. [Google Scholar] [CrossRef]
- Walker, C.S.; Eftekhari, S.; Bower, R.L.; Wilderman, A.; Insel, P.A.; Edvinsson, L.; Waldvogel, H.J.; Jamaluddin, M.A.; Russo, A.F.; Hay, D.L. A second trigeminal CGRP receptor: Function and expression of the AMY1 receptor. Ann. Clin. Transl. Neurol. 2015, 2, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Haanes, K.A.; Chan, K.Y.; MaassenVanDenBrink, A. Comment on “A second trigeminal CGRP receptor: Function and expression of the AMY1 receptor”. Ann. Clin. Transl. Neurol. 2016, 3, 307–308. [Google Scholar] [CrossRef] [Green Version]
- Ashina, M.; Hansen, J.M.; Do, T.P.; Melo-Carrillo, A.; Burstein, R.; Moskowitz, M.A. Migraine and the trigeminovascular system-40 years and counting. Lancet Neurol. 2019, 18, 795–804. [Google Scholar] [CrossRef]
- Nowaczewska, M.; Straburzyński, M.; Waliszewska-Prosół, M.; Meder, G.; Janiak-Kiszka, J.; Kaźmierczak, W. Cerebral Blood Flow and Other Predictors of Responsiveness to Erenumab and Fremanezumab in Migraine-A Real-Life Study. Front. Neurol. 2022, 13, 895476. [Google Scholar] [CrossRef] [PubMed]
- de Vries Lentsch, S.; Al-Hassany, L.; Ferrari, M.D.; Terwindt, G.M.; MaassenVanDenBrink, A. CGRP-mediated trigeminovascular reactivity in migraine patients treated with erenumab. J. Neurol. Neurosurg. Psychiatry. 2022, 93, 911–912. [Google Scholar] [CrossRef] [PubMed]
- Scuteri, D.; Corasaniti, M.T.; Tonin, P.; Nicotera, P.; Bagetta, G. Role of CGRP pathway polymorphisms in migraine: A systematic review and impact on CGRP mAbs migraine therapy. J. Headache Pain 2021, 22, 87. [Google Scholar] [CrossRef]
- Mulvany, M.J.; Halpern, W. Contractile properties of small arterial resistance vessels in spontaneously hypertensive and normotensive rats. Circ. Res. 1977, 41, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Neubig, R.R.; Spedding, M.; Kenakin, T.; Christopoulos, A. International Union of Pharmacology Committee on Receptor Nomenclature and Drug Classification. XXXVIII. Update on terms and symbols in quantitative pharmacology. Pharmacol. Rev. 2003, 55, 597–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| HCA | HMMA | Potency Difference | |
|---|---|---|---|
| Olcegepant [17,18] | pKb 1 nM *: 9.56 ± 0.22 pKb 10 nM: 9.33 ± 0.25 pKb 100 nM: 9.13 ± 0.17 | pA2: 10.59 ± 0.54 | 10.7× more potent in HMMA 18.2× more potent in HMMA 28.8× more potent in HMMA |
| Telcagepant [19,20] | pA2: 8.43 ± 0.24 | pKb 1 µM: 8.03 ± 0.16 | 2.5× more potent in HCA |
| Atogepant [21] | pKb 10 nM: 9.42 ± 0.22 pKb 100 nM: 9.11 ± 0.34 pKb 1 µM: 8.64 ± 0.21 | pKb 10 nM: 10.96 ** | 34.7× more potent in HMMA 70.8× more potent in HMMA 208.9× more potent in HMMA |
| Ubrogepant [21] | pA2: 8.86 ± 0.39 | pKb 10 nM: 9.70 ** | 6.9× more potent in HMMA |
| Rimegepant [22] | pKb 1 nM *: 9.74 ± 0.05 pKb 10 nM: 9.25 ± 0.15 pKb 100 nM: 8.71 ± 0.16 pKb 1 µM: 8.43 ± 0.25 | pA2: 10.02 ± 0.33 | 1.9× more potent in HMMA 5.9× more potent in HMMA 20.4× more potent in HMMA 38.9× more potent in HMMA |
| Zavegepant Figure 1 of [23] | pA2: 9.92 ± 0.24 | pKb 10 nM: 10.02 ± 0.07 | 1.3× more potent in HMMA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Vries, T.; Boucherie, D.M.; van den Bogaerdt, A.; Danser, A.H.J.; MaassenVanDenBrink, A. Blocking the CGRP Receptor: Differences across Human Vascular Beds. Pharmaceuticals 2023, 16, 1075. https://doi.org/10.3390/ph16081075
de Vries T, Boucherie DM, van den Bogaerdt A, Danser AHJ, MaassenVanDenBrink A. Blocking the CGRP Receptor: Differences across Human Vascular Beds. Pharmaceuticals. 2023; 16(8):1075. https://doi.org/10.3390/ph16081075
Chicago/Turabian Stylede Vries, Tessa, Deirdre M. Boucherie, Antoon van den Bogaerdt, A. H. Jan Danser, and Antoinette MaassenVanDenBrink. 2023. "Blocking the CGRP Receptor: Differences across Human Vascular Beds" Pharmaceuticals 16, no. 8: 1075. https://doi.org/10.3390/ph16081075