1. Introduction
The treatment of cancer mainly includes surgery, chemotherapy, radiation therapy, and immunotherapy, and the choice of therapy has undergone evolutionary changes as the understanding of the underlying biological process has improved. Among these, cancer immunotherapy allows disease treatment via artificial stimulation of the immune system. However, despite these advancements, there are still some limitations in the field of immunotherapy [
1], which include the unpredictable treatment efficacy and patient response, the need for additional biomarkers, and the adverse immunological responses, especially the cytokine storm toxicity [
2]. Therefore, the search for high-efficacy and fewer-toxicity immunomodulators has become the focus of current research.
Codonopsis pilosula (Franch.) Nannf., which belongs to the family Campanulaceae, has been used in food and prescribed in traditional folk medicine across Asian countries for thousands of years [
3,
4]. The roots of
C. pilosula have long been prescribed for nourishing the spleen and tonifying Qi of the stomach [
5,
6]; importantly,
C. pilosula presents significant immunomodulatory activity with a high safety [
7,
8]. Various types of compounds have been extracted from the roots of
C. pilosula, including polysaccharides [
9], lignans [
10], flavonoids [
11], sesquiterpenes [
12], triterpenoids [
13], and alkaloids [
14]. Polysaccharides are one of the major biologically active substances responsible for their therapeutic functions. In recent years, several studies have reported that
C. pilosula polysaccharides (CPPS) can exert antitumor effects by activating T lymphocytes, dendritic cells, and macrophages both in vitro and in vivo [
4].
The activation of macrophages is necessary for immune system stimulation. Macrophages are an important part of the innate immune response and play a vital role in inhibiting the occurrence and metastasis of tumors [
15]. In addition, macrophages display plasticity and can differentiate into different types under the influence of environments. Several polysaccharides are known to affect the immunomodulatory activity of macrophages [
16], and they can induce tumor cell apoptosis through proinflammatory cytokines released by macrophages. Subsequently, the function of polysaccharides derived from
C. pilosula to activate and repolarize macrophages has gained attention in the past years. Several studies have suggested that CPPS can accelerate the generation of NO, TNF-α, and IL-6, as well as increase ERK1/2 and JNK by activating the MAPK signaling pathway [
17]. For instance, a study revealed that the crude polysaccharides from
C. pilosula could inhibit the proliferation of IL-4-induced M2-like TAMs and significantly increase the mRNA expression levels of IL-1, IL-6, iNOS, and TNF-α, thereby reducing the tumor volume in melanoma mice via repolarization of tumor macrophages [
18]. However, the crude CPPS also contains several neutral sub-polysaccharides, and each section may exhibit different characteristics, including their molecular weights, specific glycosidic linkages, monosaccharide compositions, the degree of branching, polymerization, and the spatial configuration of the chains [
19]. In addition, the relationship between the detailed structures of each CPPS and the pharmacological activities of tumor cells remains unclear, warranting further research.
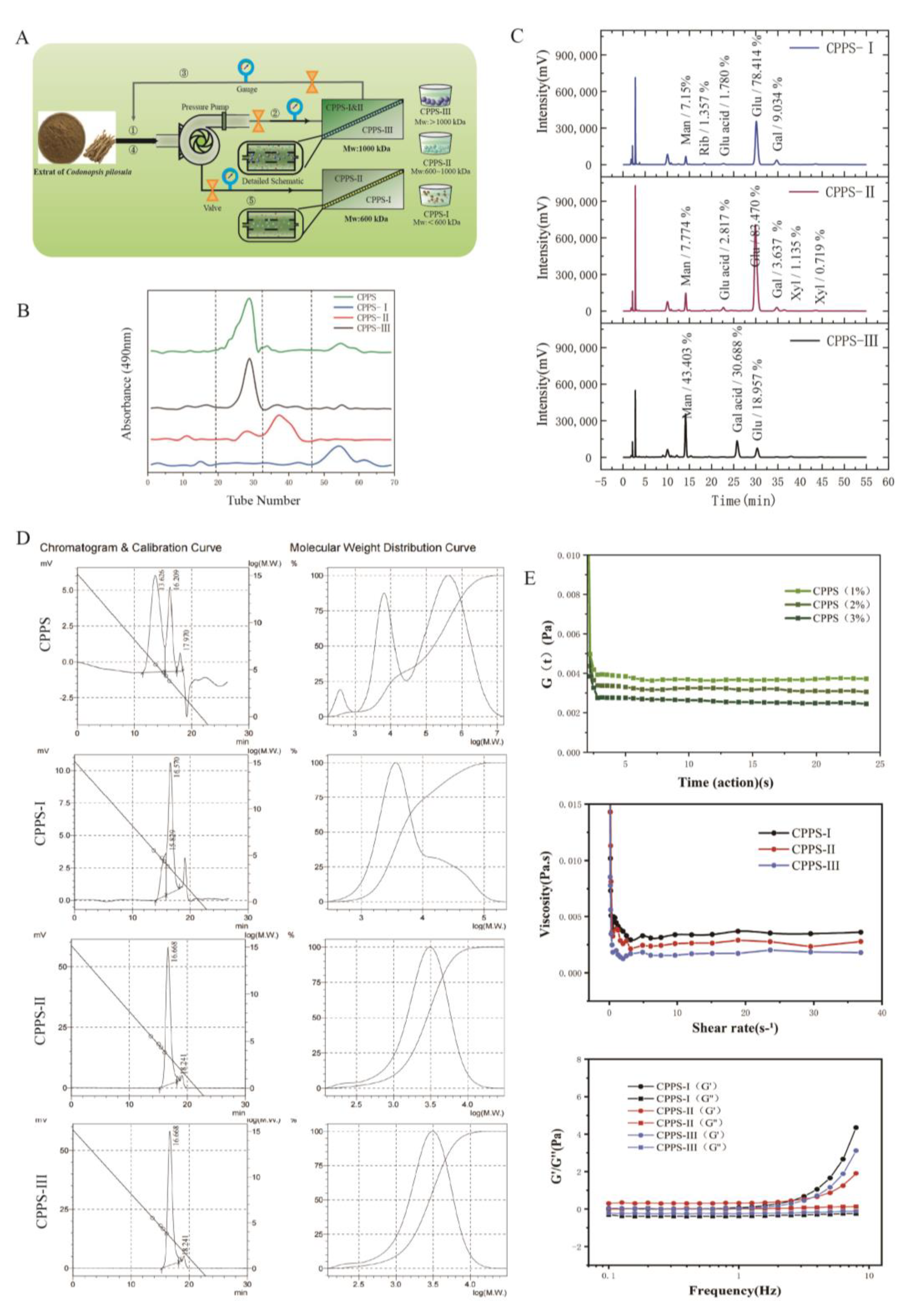
In the current study, we attempted to grade CPPS based on the average molecular weights of polysaccharides by using the membrane-separation method. The membrane-separation technology is a moderate and feasible approach to classify polysaccharides with different molecular weights [
20]. Ultrafiltration membranes have been effectively used to grade polysaccharides extracted from camellia seed cake and corn in other studies [
21,
22]. The structural characteristics of different polysaccharide fractions were investigated by NMR spectroscopy, gel permeation chromatography, Sephadex G100 gel (GE, Uppsala, Sweden) permeation, and monosaccharide composition analysis, and the immune regulation activity in RAW264.7 macrophages of different purified polysaccharides was analyzed by in vitro experiments. Furthermore, the potential effect of polysaccharides on melanoma in C57 mice was explored, and the impact on tumor-associated macrophages was investigated. This is the first study to demonstrate the potential relationship between graded CPPS structure and TAM immunomodulatory activity and examine the possible molecular action mechanisms. Our results demonstrated that moderate molecular weight is the most critical immunomodulator in the anti-melanoma effect and that it may be helpful to develop specific polysaccharide-based pharmaceutical preparations.
3. Discussion
In this study, the crude polysaccharides of Codonopsis pilosula were extracted through water extraction and by using the alcohol precipitation method, and the proteins obtained in the crude polysaccharides were removed via the Sevage method to obtain the refined polysaccharides. In the ultrafiltration system (membrane-separation method), the polysaccharide fragments of different molecular weights (CPPS-I < 60 wDa, 60 wDa < CPPS-II < 100 wDa, and CPPS-III > 100 wDa) were obtained by using ultrafiltration membranes with molecular retention of 60 and 100 wDa, respectively, according to the molecular weight distribution. CPPS-I and CPPS-II are two glucose-rich heteropolysaccharides, mainly composed of glucose, mannose, and galactose. In contrast, CPPS-III mainly comprises mannose, galacturonic acid, and glucose. By UV, FT-IR, and NMR spectra, CPPS-I-III all belong to the α-type glucans containing pyranose moieties.
The molecular weight of polysaccharides isolated from the roots of
Codonopsis pilosula varies, and different molecules of polysaccharides may have different biological activities [
30]. Therefore, we investigated the antitumor activity and the immunomodulatory potential of different molecular weight polysaccharides from
Codonopsis pilosula. The results revealed that all polysaccharides from
Codonopsis pilosula demonstrated good antitumor activity, but only CPPS-II exhibited some immunomodulatory ability. From the study of the in vitro antitumor activity of
Codonopsis polysaccharides, CPPS-I–III showed a good tumor inhibition rate against tumor cells. However, the antitumor effect of CPPS-II was much greater than that of CPPS-I and CPPS-III. The tumor inhibition rate of CPPS-II at 30 μg/mL concentration was comparable to that of the positive-control group DOX. Second, both CPPS-I-III and LPS showed significant inhibitory effects on macrophages, albeit the toxicity to macrophages could not be relied on to reflect the immune functions. Therefore, co-culture experiments were performed to investigate macrophages and tumor cells. In the co-culture experiments, all macrophages treated with polysaccharides increased the tumor suppression rate. Still, CPPS-II increased the tumor suppression rate most significantly, followed by CPPS-II-led increase of the NO expression level of macrophages and improved tumor-killing ability. At the same time, CPPS-I and CPPS-III showed no significant change in the NO expression level.
Adriamycin is a broad-spectrum antitumor drug [
31] that can effectively treat mid- to late-stage tumors as well as tumors with a propensity to disseminate, albeit it induces damage to normal cells and leads to immune system destruction and organ toxicity, which limits its further application. Therefore, effectively overcoming the adverse effects of chemotherapeutic drugs could improve the efficiency of cancer treatment.
Codonopsis polysaccharide, as an active component of traditional Chinese medicine, confers the effects of improving the immune function [
32] and anti-aging [
33], promoting metabolism [
34], and improving body functions [
35], which are beneficial to enhance the antitumor effect of chemotherapy drugs and improving the damage caused by chemotherapy drugs to patients. Therefore, we further explored the mechanism of CPPS to promote adriamycin tumor treatment, enhance immune function, and reduce its toxic side effects for antitumor treatment [
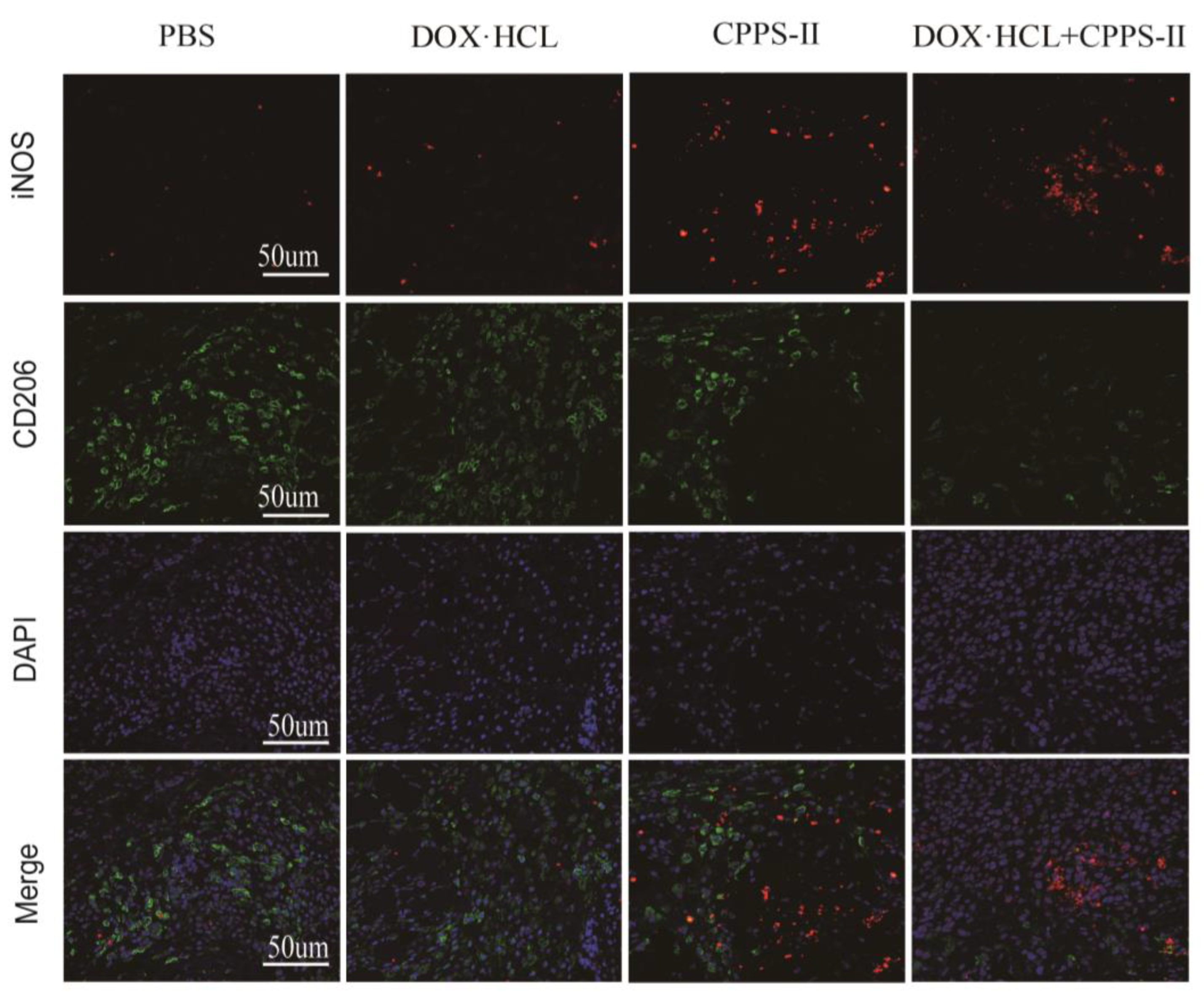
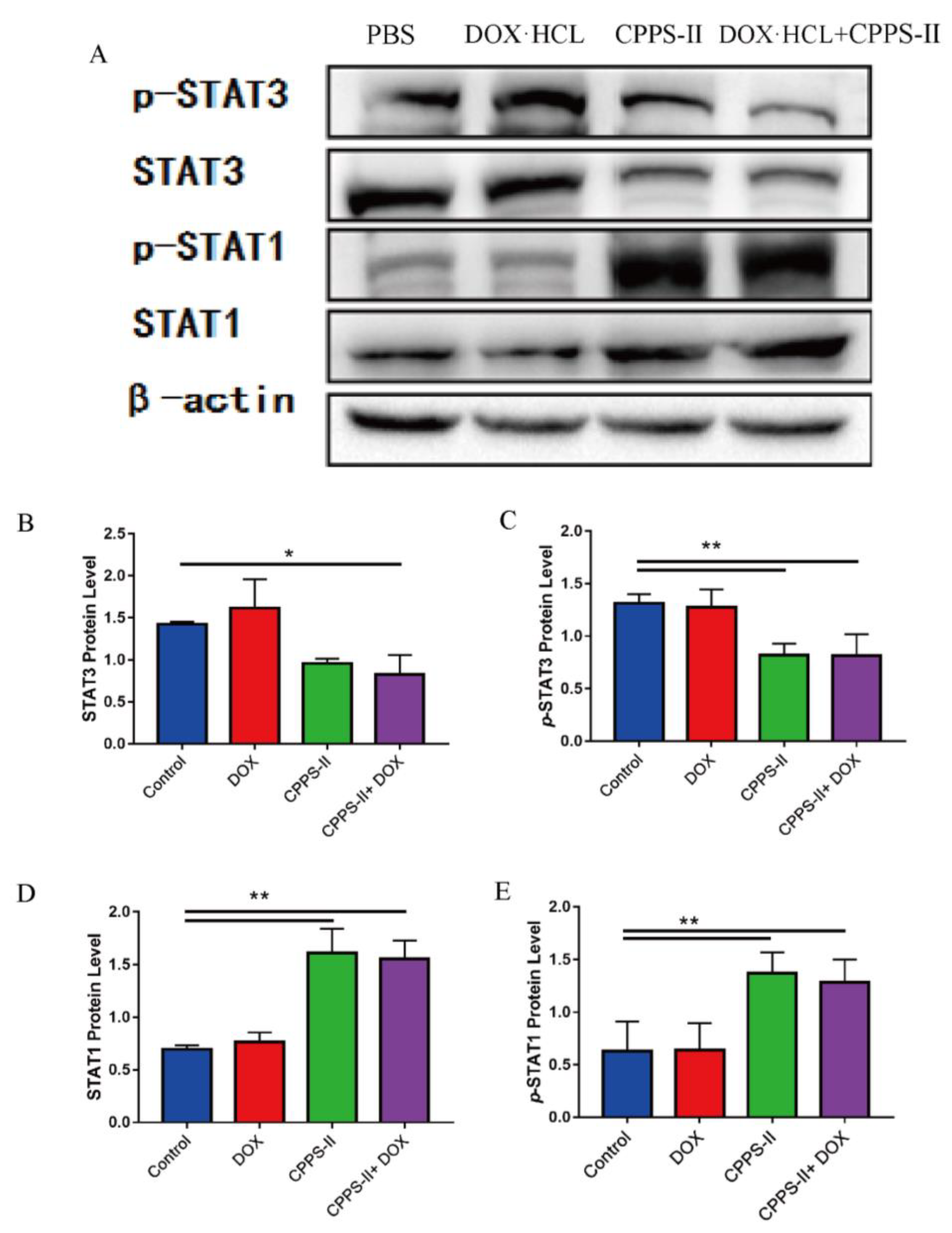
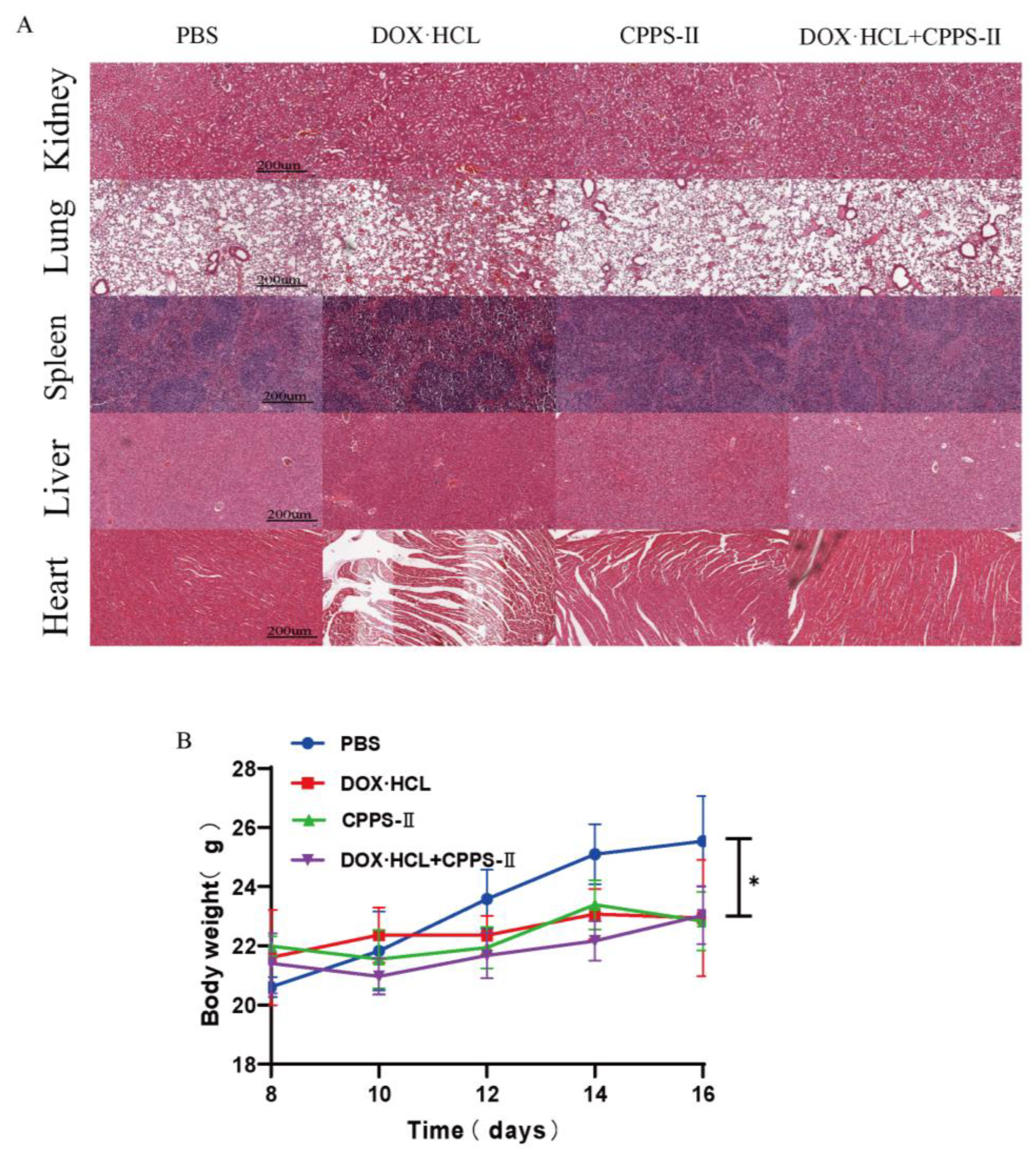
36]. First, CPPS-II, with the best efficacy, was selected from CPPS-I–III by in vitro experiments as the study of in vivo experiments. In the in vivo antitumor experiments, CPPS-II + DOX displayed better tumor suppression than polysaccharides or DOX administration alone, which facilitated the strategy of chemotherapy combined with polysaccharide immunotherapy. To investigate the regulation of immunity by polysaccharides, the phenotype of tumor tissue macrophages was determined by flowmetry, and the results revealed that the number of M1-TAMs was highest in the CPPS-II-containing administration group. In contrast, the number of M2-TAMs was significantly lower than that in the saline and DOX alone treatment groups. Second, CPPS-II monotherapy significantly decreased the p-Stat3 and Stat3 protein levels and increased the p-Stat1 and Stat1 protein expression. Considering the abovementioned experimental results, CPPS-II is likely to drive repolarization of M2 TAMs to M1 TAMs by promoting Stat1 phosphorylation and inhibiting Stat3 activation to modify tumor-associated macrophages in order to achieve tumor treatment effect through immunotherapy. H&E staining of the tumor tissues in the control group demonstrated typical pathological features of tumors. TUNEL and Ki67 results also revealed that the CPPS-II + DOX treatment group produced the most pronounced inhibition of tumor cell proliferation, indicating that the combination therapy produced a good therapeutic effect. Subsequently, the expression of markers such as CD31 and MMP9 were used to indicate the extent of tumor angiogenesis and metastasis, respectively, and the CPPS-II + DOX group significantly downregulated the expression of CD31 and MMP9 in the tumor tissues and reduced the migration ability of tumors. CPPS-II alleviated the weight loss and normal tissue damage induced by DOX drugs in the animals. These results demonstrated that CPPS-II could enhance the antitumor of DOX, regulate tumor-associated macrophages, and alleviate the damage of chemotherapeutic drugs to the organism, indicating that CPPS-II + DOX had good prospects.
4. Materials and Methods
4.1. Materials and Chemicals
The roots of Codonopsis pilosula were collected from Weiyuan, in the Shanxi province of China, and were identified by Dr. Jiushi Liu (The Institute of Medicinal Plant Development, affiliated with the Chinese Academy of Medical Sciences and Peking Union Medical College). Sephadex G-100 was purchased from GE Healthcare Life Science (Uppsala, Sweden). Monosaccharide standards, DMSO, Lipopolysaccharide (LPS), and ethylenediamine were obtained from Sigma (St. Louis, MO, USA).
RPMI-1640 and fetal bovine serum were purchased from Gibco (Carlsbad, CA, USA). Cell counting kit-8 (CCK-8) and Annexin V-FITC Apoptosis Detection Kit were obtained from Beyotime (Shanghai, China). Assay kits for interleukin-6 (IL-6), interleukin-10 (IL-10) tumor necrosis factor-alpha (TNF-α), and BCA were all obtained from Nanjing Jiancheng Bioengineering Institute (Nanjing, China). All other chemicals and solvents used were of analytical reagent grade and obtained from Sinopharm Chemical Reagent Co., Ltd. (Ningbo, China).
4.2. Polysaccharide Extraction and Purification
Codonopsis pilosula polysaccharide was isolated from the dried body powder using hot-water extraction followed by ethanol extraction. The materials were extracted three times with boiling water (1:20, w/v, 2 h for each time), and the collected solution was evaporated to a certain volume to obtain the concentrate. An equal volume of ethyl acetate was added to the concentrate and mixed for at least 5 min. The top organic layer was carefully removed using a pipe and discarded. This step was repeated thrice, and the bottom aqueous was collected for further purification. Finally, the concentrate was precipitated by 95% ethanol (80% of eventual concentration) for 4 h at room temperature and collected the precipitate. The precipitate was redissolved in pure water and lyophilized for 48 h to yield Codonopsis pilosula crude polysaccharide(C-CPPS).
The Sevage method was used to remove the protein in the crude polysaccharide to yield Codonopsis pilosula refined polysaccharides (CPPS), the CPPS was intercepted according to the Mw distribution by using ultrafiltration membrane with molecular retention of 60 and 100 wDa in the ultrafiltration system. Each fraction was collected and lyophilized to give a white powder designated CPPS-I (<60 wDa), CPPS-II (60–100 wDa), and CPPS-III (>100 wDa). The purity of the preparation was measured by a Sephadax G-100 column (2.6 cm × 100 cm, GE Tech., Uppsala, Sweden) equilibrated with distilled water. Each polysaccharide was prepared for further immunological analysis.
4.3. Polysaccharide Characterization
4.3.1. Molecular Weight and Homogeneity
Weight average molecular weight (Mw) of CPPS and sub-fractions were performed according to the report described by Li [
37]. PEO standards were used to establish a calibration curve. The analysis was conducted on a gel permeation chromatography system (Shimadzu LC20, Shimadzu Co., Kyoto, Japan) equipped with a refractive index detector (Shimadzu RID-20, Shimadzu Co., Kyoto, Japan). The samples were loaded into a TSK Ultrahydrogel™ linear column (8 × 300 mm) coupled with a TSK Ultrahydrogel™ guard column (6 × 200 mm), and the column temperature was set at 35 °C. The 0.1 N nano3 solution containing 0.06% (
w/
v) nan3 was used as the mobile phase at the flow rate of 0.6 mL/min and the injection volume was 20 μL. Data was collected by Shimadzu labsolutions HW200 workstation.
4.3.2. Rheological Properties of CPPS
Three kinds of CPPS-I, CPPS-II and CPPS-III polysaccharides were prepared, and their apparent viscosity was determined by Kinexus Lab + rheometer at 25 °C, the storage modulus G′ and loss modulus G′ of three kinds of Codonopsis pilosula polysaccharides solutions were determined by frequency scanning at 25 °C under 1% strain. The relaxation modulus g(t) PA of CPPS in 1%, 2%, and 3% solutions were calculated at 25 °C.
4.3.3. Qualitative Analysis of Monosaccharide Composition
The monosaccharide composition of the polysaccharide was analyzed by reverse-phase HPLC according to PMP derivatization procedures [
38]. Briefly, the standard monosaccharides were dissolved in deionized water and saved for PMP-label. The polysaccharides solution (2 mg/mL) and trifluoroacetic acid (4 M) were mixed for at least 5 s, then the mixture was sealed and incubated at 110 °C for 4 h in a heating block. Then, the reaction mixture was evaporated to dryness to remove residual TFA. The residue samples were reconstituted in pure water, and the standard samples were prepared as described above. Then, solution samples were dissolved in 0.6 M sodium hydroxide (50 uL), and 0.5 M PMP (100 uL), and the mixture was incubated at 70 °C for 1 h in a heating block. The mixture was neutralized by 0.3 M HCl solution and filtered through a 0.22 μm membrane. Analysis of the resulting solution was carried out using Agilent Technologies 1260 series apparatus equipped with DAD detectors and an Agilent Eclipse XDB C18 (150 mm × 4.6 mm). the flow rate was 1 mL/min, and the UV absorbance of the effluent was monitored at 250 nm. Mobile phases A and B were 0.1 M ammonium acetate, pH5.5, containing 15% acetonitrile, respectively.
4.3.4. UV and IR Spectrum Analysis
The UV-vis absorption spectra of polysaccharide samples from 200 nm to 400 nm were recorded with a UV spectrophotometer (Agilent, Santa Clara, CA, USA). The FT-IR spectrum for the polysaccharide in the 4000–400 cm−1 wavelength was obtained with a Bruker vertex spectrometer (Bruker, Karlsruhe, Germany) at room temperature.
4.3.5. NMR Analysis
The hydroxyl protons of polysaccharides were exchanged with D2O three times. Samples were lyophilized and dissolved in D2O at a 20–30 g/L concentration. all NMR spectra were recorded using an NMR spectrometer (Jeol, Kyoto, Japan).
4.3.6. SEM Analysis
Scanning electron microscopy (SEM), images were observed with field emission scanning electron microscopy (Jeol, Kyoto, Japan). The samples were sputtered with gold and placed onto double-sided adhesive tape attached to a circular specimen stub for analysis.
4.3.7. AFM Analysis
The molecular morphology of polysaccharides was recorded using a ScanAsyst AFM (Bruker, Karlsruhe, Germany).
4.4. In Vitro Evaluation
4.4.1. Cell Culture
RAW264.7 and B16 cells were purchased from the Cell Resource Center of the Chinese Academy of Medical Sciences & Peking Union Medical College (Beijing, China). Both were maintained in DMEM high-glucose medium supplemented with 100 μg/mL streptomycin, 100 U/mL penicillin, and 10% heat-inactivated fetal bovine serum. The cells were cultured at 37 °C in a humidified atmosphere containing a 5% CO2 incubator.
4.4.2. Cell Viability Assay
The effect of the polysaccharides on the viability of RAW264.7 and B16 cells was determined by the CCK-8 method. Briefly, RAW264.7 and B16 cells were seeded 100 μL at a density of 1 × 105 cells/mL in 96-well plates, respectively, and cultured overnight. CPPS-I, CPPS-II, and CPPS-III were added at final concentrations of 0, 62.5, 125, 250, 500, and 1000 μg/mL to B16. After 24 h of incubation at 37 °C in a humidified, 20 μL of the cck-8 reagent were added to each well. After 1 h of incubation, the absorbance was recorded at 560 nm by using an ELISA plate reader and then translated into macrophage viability ratio for comparison. All determinations were performed for three replicates for every sample, and three independent assays were conducted.
4.4.3. Measurement of NO
Logarithmically grown RAW264.7 cells were inoculated in 96-well culture plates at 1.0 × 104 cells per well, incubated for 24 h at 37 °C with 5% CO2 saturation, and then treated with CPPS-I, CPPS-II, and CPPS-III (0, 62.5, 125, 250, 500 and 1000 μg/mL) or LPS (1 μg/mL), followed by incubation for another 12 h. The supernatant was collected from each well for measurement of NO. The NO level was measured according to manufacturer’s instructions of NO detection kit (Beyotime Biotechnology Institute, Shanghai, China).
4.4.4. Macrophage Polarization
RAW 264.7 cells were inoculated in 6-well culture plates at 1.0 × 105 cells per well and treated with three CPPS (50 μg/mL) for 24 h. At the end of the culture, the cells were collected in centrifuge tubes, washed three times with PBS, and incubated with APC-CD11c antibody for 30 min at 4 °C in the dark. Then, the expression levels of surface receptors were determined by flow cytometry (LE-SH800SA).
4.4.5. Cell Cycle
Cell cycle was performed as previously described [
39,
40]. Briefly, B16 cells were seeded in 6-well plates at 1.0 × 10
6 cells per well and mediated by various concentrations of CPPSs. After 48 h of incubation, cells were routinely digested with trypsin and collected followed by wash with cold PBS. Then, cells were fixed with 70% ethanol at 4 °C overnight and treated with 100 mg/mL RNasesolution and PI staining at 37 °C for 30 min. The samples were detected at the wavelength of 488 nm using a flow cytometer (FACS Canto, BD Biosciences, Franklin Lakes, NJ, USA), and the relative proportions of B16 cells in each phase were analyzed with Modfit software (Verity Software House Inc., Topsham, ME, USA).
4.4.6. Annexin V-FITC/PI Detection
The extent of cell apoptosis was assayed using Annexin V FITC PI apoptosis kit. B16 cells (1 × 106 cells/mL) were seeded in 6-well plates and treated with three CPPS (50 μg/mL) for 48 h. Cells were collected following digestion with EDTA-free trypsin, wash with PBS and centrifugation for 5 min at 1000 rpm. Subsequently, cells were subjected to 5 μL of Annexin V-FITC and 5 μL of PI were added and incubated at room temperature in the dark for 10 min. Cells were then filtered through a 40 μm cell filter and tested by flow cytometry.
4.5. In Vivo Anti-Tumor Activity Evalution
4.5.1. Tumor Xenograft Models
Sub-confluent B16-F10 cells were harvested and resuspended in PBS at a concentration of 5.0 × 106 cells/mL. the subcutaneous melanoma model was established by injecting 0.1 mL of the cell suspension into the right axilla of the male C57 BL/6N mice. Once the tumor volume was approximately 50 mm3 (5–7 days after implantation), the mice were randomly divided into four groups (n = 6 each) and treated accordingly: (ⅰ) the control group (PBS); CPPS-II (10 mg/kg); Dox (1 mg/kg); and DOX (1 mg/kg) + CPPS-II (10 mg/kg). DOX.HCl was administered via the tail vein and CPPS-II was administered via the intraperitoneal injection, respectively every 2 days for 8 days. All animals were handled in compliance with internal guidelines, and the above animal protocols and operations were approved by the ethics committee of the Chinese Academy of Medical Science, Beijing, China.
4.5.2. Antitumor Efficiency
The antitumor effect was performed by the previously published work [
39]. Briefly, the tumor volume and body weight were recorded at 2-day intervals. Tumor volume was monitored by measuring the perpendicular diameter with a caliper. The estimated tumor volume was calculated as 0.5 × length × width
2. Tumor suppression rates (TSR) and tumor growth rates were calculated at the end of the study. Tumors and vital organs (heart, liver, spleen, lung, and kidney) were excised from the B16F10 tumor-bearing mice, and conduct the standard hematoxylin and eosin (H&E) staining test to evaluate the anti-tumor efficacy and systemic toxicity.
4.5.3. Immunohistochemical Staining Analysis
In addition, immunohistochemical staining was performed on paraffin-embedded tumor sections using primary antibodies for Ki67 (dilution rate as 1:500, Abcam, Cambridge, UK), a cluster of differentiation 31 (CD31, dilution rate as 1:800, Abcam, Cambridge, UK), matrix metallopeptidase 9 (MMP-9, dilution rate as 1:500, Abcam, Cambridge, UK) with biotinylated goat anti-rabbit IgG (dilution rate as 1:200, Zhongshan Jinqiao Biotechnology, Beijing, China) as the secondary antibody. Representative images were obtained using an inverted microscope (Leica, Weztlar, Germany). The expression levels of antigens were semi-quantitatively analyzed at 20× magnification using Image-Pro Plus software (Media Cybernetics, Rockville, MD, USA).
4.5.4. TUNEL Assay
To further characterize apoptosis levels of the tumor section, TUNEL assays were performed following the steps recommended by the manufacturer (Beyotime, Shanghai, China). Biotin-antibody diluent was dropped on the sections for 3′-hydroxyl termini of DNA double-strand breaks staining and incubated for 60 min at 37 °C. Next, streptavidin-HRP was used as the secondary antibody for 40 min, followed by DAB staining and hematoxylin counterstaining. Positive staining was analyzed under an inverted microscope (Leica, Germany) at 20× magnification and analyzed on Image J.
4.5.5. Analysis of Macrophage Polarization by Flow Cytometry
Macrophages infiltrating the tumor section were evaluated by flow cytometry. tumor tissues were harvested, minced, and incubated with serum-free RPMI-1640 medium. After digested by motor grinding (GentleMACS™, Miltenyi Biotec, Bergisch Gladbach, Germany), The dissociated cells were passed through the 40-μm nylon mesh and lysis of the red blood cells (RBCs). To stain macrophage cluster, anti-CD45 FITC (isotype control: Mouse IgG1, κ), an-ti-F4/80 APC (isotype control: Rat IgG2b, κ), anti-CD11b PE (isotype control: Mouse IgG1, κ), anti-CD206 PerCP (isotype control: Rat IgG2a, κ), anti-IAIE Brilliant Violet 421 (isotype control: Rat IgG2b, κ) were added to approximately 2 × 106 cells suspended in PBS with fluorescent probes and incubated for 30 min. All staining reactions were performed in a final volume of 100 μL at 4 °C. Data was acquired using an 18-color flow cytometer (LSRII, BD, USA) and analyzed using FlowJo v10.0 software (Tree Star Inc., Ashland, OR, USA).
4.5.6. Immunofluorescence Examination of Macrophages
Paraffin-embedded tumor sections were dewaxed with a gradient ethanol solution, then boiled in citrate buffer for antigen repair. Next, the primary antibodies were added to the surface of the sample slide and incubated in a wet box at 4 °C overnight, followed by the secondary antibody incubation at room temperature for about 50 min in the dark. All the antibodies are diluted 200 times for use. DAPI was used to counterstain nuclei for 10 min. The primary antibodies included: rat polyclonal CD206 antibody (Abcam, Cambridge, UK) and rat polyclonal iNOS antibody (Abcam, Cambridge, UK). CD206 and iNOS were used to label anti-inflammatory (M2) and pro-inflammatory (M1) macrophages, respectively. Fluorescence images were obtained by using a Nikon confocal microscope (SONY, Tokyo, Japan), and the average fluorescence intensity was quantitated by ImageJ (NIH, Bethesda, MD, USA). All staining experiments were performed three times with different sections.
4.5.7. ELISA Experiments
Two commercial ELISA kits (IL-10&TGF-β, Invitrogen, Waltham, MA, USA) were used to evaluate IL-10&TGF-β levels in tumor tissues. Assays were performed according to our previously reported work [
41]. 100 mg tumor tissues were homogenized in 1 mL PBS and stored overnight at −20 °C, Next, the samples were subjected to two freeze–thaw cycles and centrifuged at 4 °C for 10 min to separate supernatants. the concentration of total protein in supernatants was analyzed by the BCA method (Beyotime, Shanghai, China). The sandwich ELISA method was performed according to the manufacturer’s instructions, and OD values were measured at 450 nm using a microplate reader. The standard curve was generated using a known concentration of recombinant cytokines and used to calculate the cytokine concentration of the samples.
4.5.8. Western Blot Assay
The tumor tissue homogenate solution was lysed by lysis buffer containing protease inhibitors to extract the total proteins. Then, the samples were loaded onto 10% SDS-PAGE gels of separation and transferred to the PVDF membrane (Millipore Sigma, St. Louis, MO, USA). Bovine serum albumin (5%) in Tris-buffer saline containing 0.1% Tween-20 (TBST) was used to block the unspecific proteins for 1 h at room temperature. The primary antibodies were used by diluting 1000 times with TBST buffer. After incubating with the following primary antibodies: phosphorylated Stat1 (Abcam, Cambridge, UK), Stat1 (Abcam, Cambridge, UK), phosphorylated Stat3 (Abcam, Cambridge, UK), or Stat3 (Abcam, Cambridge, UK). Rabbit anti-β-actin (Perprotech, China) was employed as a loading control, the membranes were incubated with HRP-labeled secondary antibodies for 1 h at room temperature. The secondary antibody was used by diluting 2000 times with TBST buffer. Finally, protein expression was quantitatively measured and visualized by SuperLumia ECL HRP Substrate Kit (Servicebio) and Image J.
4.6. Statistical Analyses
Data were presented as mean ± standard deviation (SD) of a minimum of three replicates unless otherwise indicated. Comparisons among multiple groups were conducted by one-way ANOVA followed by analysis using GraphPad Prism 8.0 (La Jolla, CA, USA). p values < 0.05 and <0.01 were considered to indicate statistical significance.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







