
Annona muricata Leaf as an Anti-Cryptosporidial Agent: An In Silico Molecular Docking Analysis and In Vivo Studies
 , ,
, ,
Abstract
:1. Introduction
2. Results
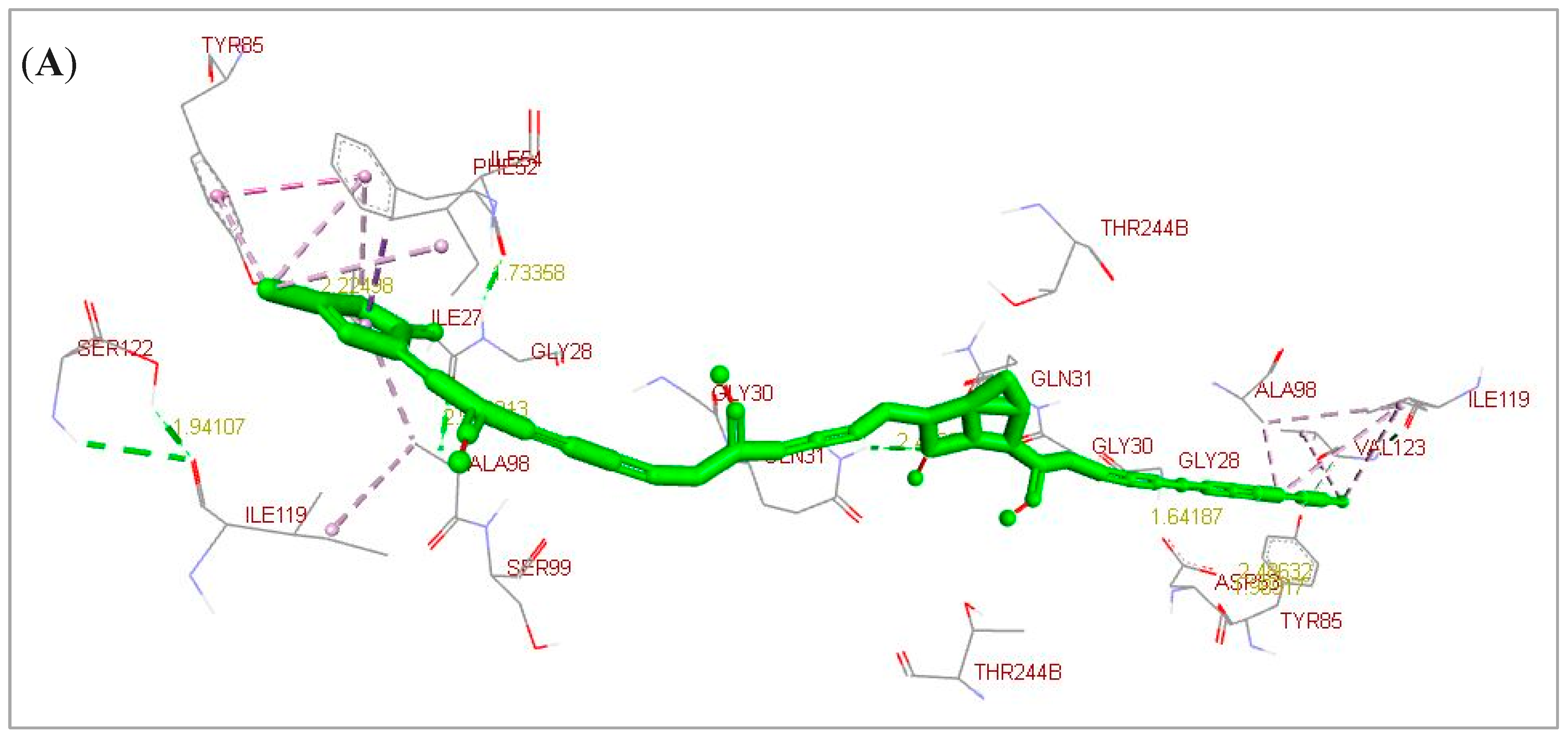
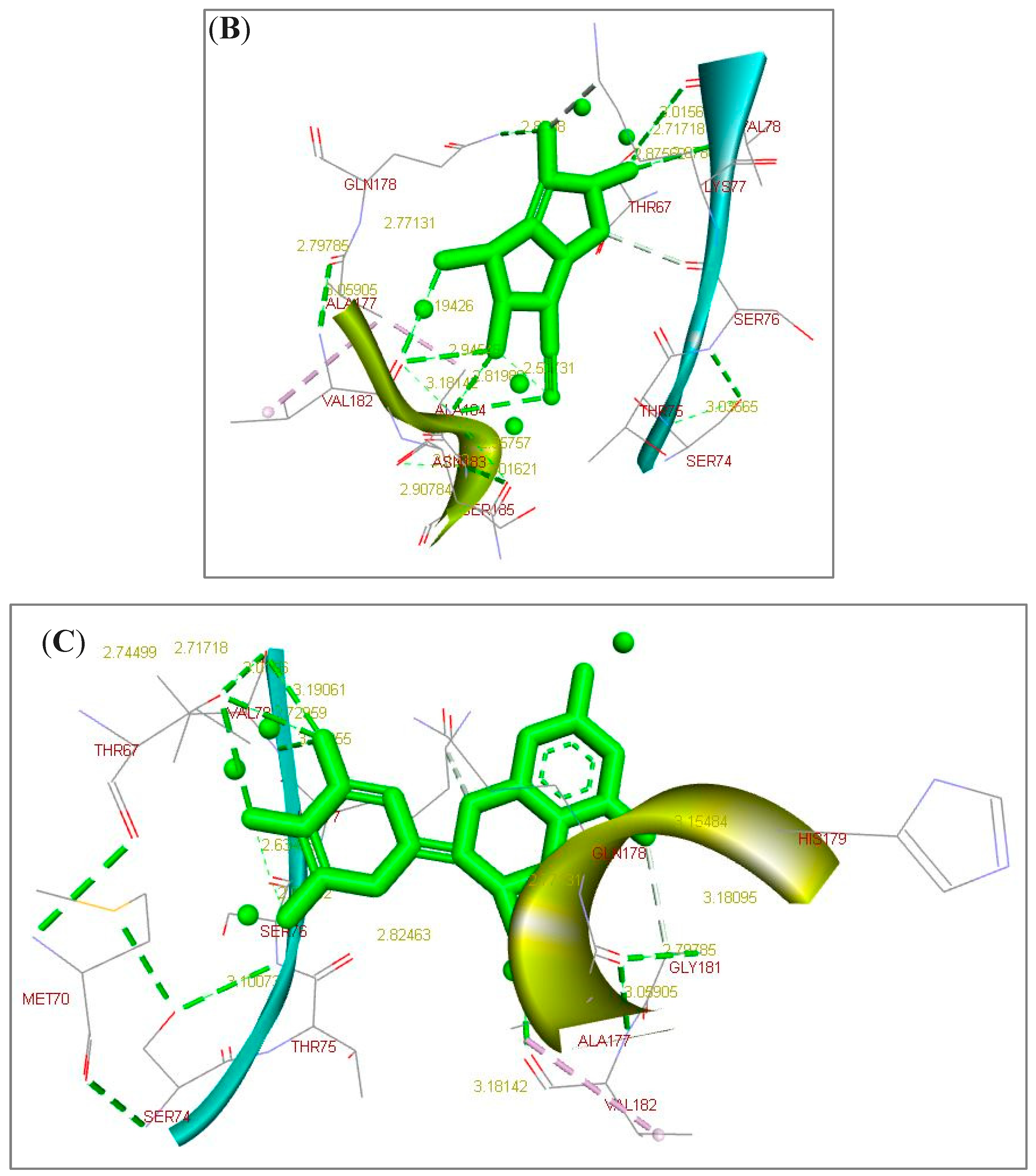
2.1. Docking Simulation Analysis Results
2.2. Oocyst Shedding
2.3. Histopathological Findings
2.4. Immunohistochemical Studies
3. Discussion
4. Materials and Methods
4.1. Molecular Docking Simulation Analysis
4.2. Experimental Animals
4.3. Animal Groups
4.4. Induction of Immunosuppression
4.5. Induction of Cryptosporidium Infection
4.6. Drug Preparation and Regimens
4.7. Drugs Assessment
4.7.1. Parasitological Examination
group)*100)/(mean value of infected untreated group).
4.7.2. Histopathological Examination
4.7.3. Immunohistochemical Examination
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Ghareeb, M.A.; Sobeh, M.; Aboushousha, T.; Esmat, M.; Mohammed, H.S.; El-Wakil, E.S. Polyphenolic profile of herniaria hemistemon aerial parts extract and assessment of its anti-cryptosporidiosis in a murine model: In silico supported in vivo study. Pharmaceutics 2023, 15, 415. [Google Scholar] [CrossRef] [PubMed]
- Desai, N.T.; Sarkar, R.; Kang, G. Cryptosporidiosis: An under-recognized public health problem. Trop. Parasitol. 2012, 2, 91–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdou, A.G.; Harba, N.M.; Afifi, A.F.; Elnaidany, N.F. Assessment of Cryptosporidium parvum infection in immunocompetent and immunocompromised mice and its role in triggering intestinal dysplasia. Int. J. Infect. Dis 2013, 17, e593–e600. [Google Scholar] [CrossRef] [Green Version]
- Jumani, R.S.; Bessoff, K.; Love, M.S.; Miller, P.; Stebbins, E.E.; Teixeira, J.E.; Huston, C.D. A novel piperazine-based drug lead for cryptosporidiosis from the medicines for malaria venture open-access malaria box. Antimicrob. Agents Chemother. 2018, 62, e01505-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Certad, G.; Ngouanesavanh, T.; Guyot, K.; Gantois, N.; Chassat, T.; Mouray, A.; Creusy, C. Cryptosporidium parvum, a potential cause of colic adenocarcinoma. Infect. Agents Cancer 2007, 2, 22. [Google Scholar] [CrossRef] [Green Version]
- Benamrouz, S.; Guyot, K.; Gazzola, S.; Mouray, A.; Chassat, T.; Delaire, B.; Certad, G. Cryptosporidium parvum infection in SCID mice infected with only one oocyst: qPCR assessment of parasite replication in tissues and development of digestive cancer. PLoS ONE 2012, 7, e51232. [Google Scholar] [CrossRef]
- Benamrouz, S.; Conseil, V.; Chabé, M.; Praet, M.; Audebert, C.; Blervaque, R.; Certad, G. Cryptosporidium parvum- Cryptosporidium parvum-induced ileo-caecal adenocarcinoma and Wnt signaling in a mouse model. Dis. Model. Mech. 2014, 7, 693–700. [Google Scholar] [CrossRef] [Green Version]
- Sparks, H.; Nair, G.; Castellanos-Gonzalez, A.; White, A. Treatment of Cryptosporidium: What we know, gaps, and the way forward. Curr. Trop. Med. Rep. 2015, 2, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Krstin, S.; Sobeh, M.; Braun, M.S.; Wink, M. Tulbaghia Violacea and Allium Ursinum extracts exhibit anti-parasitic and antimicrobial activities. Molecules 2018, 23, 313. [Google Scholar] [CrossRef] [Green Version]
- Abdelmaksoud, H.F.; El-Ashkar, A.M.; Elgohary, S.A.; El-Wakil, E.S. Potential therapeutic and prophylactic effects of asafoetida in murine cryptosporidiosis. J. Parasit. Dis. 2020, 44, 646–653. [Google Scholar] [CrossRef]
- Abd El-Hamed, W.F.; Yousef, N.S.; Mazrou, Y.S.; Elkholy, W.A.; El-Refaiy, A.I.; Elfeky, F.A.; Albadrani, M.; El-Tokhy, A.I.; Abdelaal, K. Anticryptosporidium efficacy of Olea Europaea and Ficus Carica leaves extract in immunocompromised mice associated with biochemical characters and antioxidative system. Cells 2021, 10, 2419. [Google Scholar] [CrossRef] [PubMed]
- El-Wakil, E.S.; El-Shazly, M.A.; El-Ashkar, A.M.; Aboushousha, T.; Ghareeb, M.A. Chemical profiling of Verbena officinalis and assessment of its anti-Cryptosporidial Activity in experimentally infected Immunocompromised Mice. Arab. J. Chem. 2022, 15, 103945. [Google Scholar] [CrossRef]
- Soufy, H.; Nadia, M.; Nasr, S.M.; Abd El-Aziz, T.H.; Khalil, F.A.; Ahmed, Y.F.; Abou Zeina, H.A. Effect of Egyptian Propolis on Cryptosporidiosis in Immunosuppressed Rats with Special Emphasis on Oocysts Shedding, Leukogram, Protein Profile and Ileum Histopathology. Asian Pac. J. Trop. Med. 2017, 10, 253–262. [Google Scholar] [CrossRef]
- Vila-Nova, N.S.; de Morais, S.M.; Falcão, M.J.; Negreiros Alcantara, T.T.; Travassos Ferreira, P.A.; Barreira Cavalcanti, E.S.; Pinto Vieira, I.G.; Campello, C.C.; Wilson, M. Different susceptibilities of Leishmania spp. promastigotes to the Annona muricata acetogenins annonacinone and corossolone, and the Platymiscium floribundum coumarin scoparone. Exp. Parasitol. 2013, 133, 334–338. [Google Scholar] [CrossRef] [Green Version]
- Rady, I.; Bloch, M.B.; Chamcheu, R.C.N.; Banang Mbeumi, S.; Anwar, M.R.; Mohamed, H.; Chamcheu, J.C. Anticancer properties of graviola (Annona muricata): A comprehensive mechanistic review. Oxid. Med. Cell. Longev. 2018, 2018, 1826170. [Google Scholar] [CrossRef] [Green Version]
- Mutakin, M.; Fauziati, R.; Fadhilah, F.N.; Zuhrotun, A.; Amalia, R.; Hadisaputri, Y.E. Pharmacological activities of soursop (Annona muricata Lin.). Molecules 2022, 27, 1201. [Google Scholar] [CrossRef] [PubMed]
- Nolasco-González, Y.; Chacón-López, M.A.; Ortiz-Basurto, R.I.; Aguilera-Aguirre, S.; González-Aguilar, G.A.; Rodríguez-Aguayo, C.; Montalvo-González, E. Annona muricata leaves as a source of bioactive compounds: Extraction and quantification using ultrasound. Horticulturae 2022, 8, 560. [Google Scholar] [CrossRef]
- El-Wakil, E.S.; Abdelmaksoud, H.F.; AbouShousha, T.S.; Ghallab, M.M.I. Evaluation of Annona muricata (Graviola) leaves activity against experimental trichinellosis: In vitro and in vivo studies. J. Helminthol. 2021, 95, e53. [Google Scholar] [CrossRef]
- Bartkova, J.; Thullberg, M.; Slezak, P.; Jaramillo, E.; Rubio, C.; Thomassen, L.H.; Bartek, J. Aberrant expression of G1-phase cell cycle regulators in flat and exophytic adenomas of the human colon. Gastroenterology 2001, 120, 1680–1688. [Google Scholar] [CrossRef]
- Albasri, A.M.; Elkablawy, M.A.; Ansari, I.A.; Alhujaily, A.S. Prognostic significance of Cyclin D1 over-expression in colorectal cancer: An experience from Madinah, Saudi Arabia. APJCP 2019, 20, 2471. [Google Scholar] [CrossRef]
- Fan, J.; Fu, A.; Zhang, L. Progress in molecular docking. Quant. Biol. 2019, 7, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Ghallab, M.; Khater, M.; Tyler, K.; Bouzid, M.; Kamal, A.; El-Badry, A. High-resolution melting curve (hrm) analysis in genotypic discrimination of Cryptosporidium isolates from stool of Egyptian children. J. Egypt. Soc. Parasitol. 2018, 48, 557–562. [Google Scholar]
- Audebert, C.; Bonardi, F.; Caboche, S.; Guyot, K.; Touzet, H.; Merlin, S.; Gantois, N.; Creusy, C.; Meloni, D.; Mouray, A.; et al. Genetic basis for virulence differences of various Cryptosporidium parvum carcinogenic isolates. Sci. Rep. 2020, 10, 7316. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.E.; Castro, P.M.; Chagas, A.C.; França, S.C.; Beleboni, R.O. In vitro anthelmintic activity of aqueous leaf extract of Annona muricata L. (Annonaceae) against Haemonchus contortus from sheep. Exp. Parasitol. 2013, 134, 327–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Tawiil, G.A.; Noaman, E.A.; Askar, M.A.; El Fatih, N.M.; Mohamed, H.E. Anticancer and Apoptogenic Effect of A. muricata and Low-Dose Radiation in Tumor Xenograft in Mice. Integr. Cancer Ther. 2020, 19, 1534735419900930. [Google Scholar] [CrossRef] [PubMed]
- Souza, M.M.; Bevilaqua, C.M.; Morais, S.M.; Costa, C.T.; Silva, A.R.; Braz-Filho, R. Anthelmintic acetogenin from Annona squamosa L. Seeds. An. Acad. Bras. Ciências 2008, 80, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Vieira, L.S.; Cavalcante, A.C.; Pereira, R.; Dantas, M.; Ximenes, L.B. Evaluation of anthelmintic efficacy of plants available in Ceará state, northeast Brazil, for the control of goat gastrointestinal nematodes. Rev. Med. Vet. 1999, 150, 447–452. [Google Scholar]
- Nguyen-Pouplin, J.; Tran, H.; Tran, H.; Phan, T.A.; Dolecek, C.; Farrar, J.; Tran, T.H.; Caron, P.; Bodo, B.; Grellier, P. Antimalarial and cytotoxic activities of ethnopharmacologically selected medicinal plants from South Vietnam. J. Ethnopharmacol. 2007, 109, 417–427. [Google Scholar] [CrossRef]
- Somsak, V.; Polwiang, N.; Chachiyo, S. In vivo antimalarial activity of Annona muricata leaf extract in mice infected with Plasmodium berghei. J. Pathog. 2016, 2016, 3264070. [Google Scholar] [CrossRef] [Green Version]
- Yamthe, L.R.T.; Fokou, P.V.T.; Mbouna, C.D.J.; Keumoe, R.; Ndjakou, B.L.; Djouonzo Mfopa, A.N.; Legac, J.; Tsabang, N.; Gut, J.; Rosenthal, P.J.; et al. Extracts from Annona Muricata L. and Annona Reticulata L. (Annonaceae) Potently and Selectively Inhibit Plasmodium Falciparum. Medicines 2015, 2, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Coria-Téllez, A.V.; Montalvo-Gónzalez, E.; Yahia, E.; Obledo-Vázquez, E. Annona muricata: A comprehensive review on its traditional medicinal uses, phytochemicals, pharmacological activities, mechanisms of action and toxicity. Arab. J. Chem. 2018, 11, 662–691. [Google Scholar] [CrossRef] [Green Version]
- Enemark, H.; Ahrens, P.; Bille-hansen, V.; Heegaard, P.; Vigre, H.; Thamsborg, S.; LIND, P. Cryptosporidium parvum: Infectivity and pathogenicity of the ‘porcine’ genotype. Parasitol 2003, 126, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Verdon, R.; Polianski, J.; Grodet, A.; Garry, L.; Carbon, C. Cryptosporidium parvum biliary tract infection in adult immunocompetent and immunosuppressed mice. J. Med. Microbiol. 1998, 47, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Machicado, C.; Marcos, L.A. Carcinogenesis associated with parasites other than Schistosoma, Opisthorchis and Clonorchis: A systematic review. Int. J. Cancer 2016, 138, 2915–2921. [Google Scholar] [CrossRef]
- Torres, P.H.M.; Sodero, A.C.R.; Jofily, P.; Silva, F.P., Jr. Key Topics in Molecular Docking for Drug Design. Int. J. Mol. Sci. 2019, 20, 4574. [Google Scholar] [CrossRef] [Green Version]
- Pantsar, T.; Poso, A. Binding Affinity via Docking: Fact and Fiction. Molecules 2018, 23, 1899. [Google Scholar] [CrossRef] [Green Version]
- Cook, W.J.; Senkovich, O.; Hernandez, A.; Speed, H.; Chattopadhyay, D. Biochemical and structural characterization of Cryptosporidium parvum Lactate dehydrogenase. Int. J. Biol. Macromol. 2015, 74, 608–619. [Google Scholar] [CrossRef] [Green Version]
- Gaillard, T. Evaluation of AutoDock and AutoDock Vina on the CASF-2013 Benchmark. J. Chem. Inf. Model. 2018, 58, 1697–1706. [Google Scholar] [CrossRef]
- Gurung, A.B.; Bhattacharjee, A.; Ali, M.A. Exploring the physicochemical profile and the binding patterns of selected novel anticancer Himalayan plant derived active compounds with macromolecular targets. Inform. Med. Unlocked 2016, 5, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Komori, A.; Li, M.; Chen, X.; Yang, A.W.H.; Sun, X.; Liu, Y.; Hung, A.; Zhao, X.; Zhou, L. Multi-ligand molecular docking, simulation, free energy calculations and wavelet analysis of the synergistic effects between natural compounds baicalein and cubebin for the inhibition of the main protease of SARS-CoV-2. J. Mol. Liq. 2023, 374, 121253. [Google Scholar] [CrossRef]
- Li, H.; Hung, A.; Yang, A.W.H. Herb-target virtual screening and network pharmacology for prediction of molecular mechanism of Danggui Beimu Kushen Wan for prostate cancer. Sci. Rep. 2021, 11, 6656. [Google Scholar] [CrossRef] [PubMed]
- Rehg, J.E.; Hancock, M.L.; Woodmansee, D.B. Characterization of a dexamethasone-treated rat model of cryptosporidial infection. J. Infect. Dis. 1988, 158, 1406–1407. [Google Scholar] [CrossRef]
- El-Wakil, E.S.; Mohamed, E.A.; El-Wakil, E.A.; AbouShousha, T.S.; Amer, N.M. Anti-cryptosporidial activity of Camellia sinensis (green tea extract) in experimentally infected immunocompromised mice. Acta Protozool. 2022, 61, 23–34. [Google Scholar] [CrossRef]
- Arrowood, M.J. Improved purification methods for calf-derived Cryptosporidium parvum oocysts using discontinuous sucrose and cesium chloride gradients. J. Eukaryot. Microbiol. 1996, 43, 89S. [Google Scholar] [CrossRef] [PubMed]
- Love, M.S.; Beasley, F.C.; Jumani, R.S.; Wright, T.M.; Chatterjee, A.K.; Huston, C.D.; McNamara, C.W. A high throughput phenotypic screen identifies clofazimine as a potential treatment for cryptosporidiosis. PLoS Negl. Trop. Dis. 2017, 11, e0005373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Brasseur, P.; Agnamey, P.; Leméteil, D.; Favennec, L.; Ballet, J.J.; Rossignol, J.F. Long-lasting anticryptosporidial activity of nitazoxanide in an immunosuppressed rat model. Folia Parasitol. 2003, 50, 19–22. [Google Scholar] [CrossRef] [Green Version]
- Ghallab, M.M.I.; Abdel Aziz, I.Z.; Shoeib, E.Y.; El-Badry, A.A. Laboratory utility of coproscopy, copro immunoassays and copro nPCR assay targeting Hsp90 gene for detection of Cryptosporidium in children, Cairo, Egypt. JOPD 2016, 40, 901–905. [Google Scholar] [CrossRef] [Green Version]
- Garcia, L.S. (Ed.) Clinically important human parasites: Intestinal protozoa: Cryptosporidium spp. In Diagnostic Medical Parasitology, 5th ed.; ASM Press: Washington, DC, USA, 2007; pp. 761–818. [Google Scholar]
- Hosking, B.C.; Watson, T.G.; Leathwick, D.M. Multigeneric resistance to oxfendazole by nematodes in cattle. Vet. Rec. 1996, 138, 67–68. [Google Scholar] [CrossRef]
- Drury, R.A.; Wallington, E.A. Carleton’s Histological Technique, 5th ed.; Oxford University Press: New York, NY, USA, 1980; p. 546. [Google Scholar]
- Hsu, S.D.; Schwartze, G.M.; Maxwell, V.L. Prolonged survival of a patient with advanced colonic cancer. Dis. Colon Rectum 1981, 24, 636–638. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lead/Ligand @ | Binding Affinity # | Ki | Lead-Target Protein Binding and Interactions (Non-Covalent Intermolecular Interactions) |
|---|---|---|---|
| Annonacin | −6.11 | 33.48 * | H-bonds (GLY28.B and GLN31.B). |
| Casuarine | −6.32 | 23.25 * | Electrostatic and hydrophobic interactions (GLY28, ILE54, TYR85, ALA98, ILE119, and VAL123). |
| L-epigallocatechin | −7.51 | 3.12 * | H-bonds (THR67.B, SER74.B, VAL78.B, GLN178.A, VAL182.A, ASN183.A, and ALA184.A). |
| P-coumaric acid | −7.81 | 1.88 * | H-bonds (THR67.B, SER76.B, VAL78.B, GLN178.A, and VAL182.A). |
| Ellagic acid | −9.64 | 85.42 Δ | Electrostatic and hydrophobic interactions (GLY181). |
| NTZ | −7.03 | 7.06 * | H-bonds (GLY30.A, GLN31.B, LYS62.A, and LYS62.B). |
| Groups | 10th Day PI | 12th Day PI | 15th Day PI | 20th Day PI | ||||
|---|---|---|---|---|---|---|---|---|
| M ± SE × 103 | PR% | M ± SE × 103 | PR% | M ± SE × 103 | PR% | M ± SE × 103 | PR% | |
| GI | 83 ± 1.4 aA | 1% | 16.3 ±. 8 bC | 81% | 14 ± 1.6 cC | 84% | 12.3 ±1.1 dC | 86% |
| GII | 82.9 ± 1.3 aA | 1% | 30.8 ± 1.3 bB | 64% | 29.1 ± 1.1 cB | 67% | 27 ± 1.9 dB | 70% |
| GIII | 83.9 ±. 9 aA | - | 84.4 ± 1.9 aA | - | 87.5 ± 1 aA | 0 | 90 ± 2.2 aA | - |
| Groups | 90th Day PI | 93rd Day PI | 95th Day PI | 100th Day PI | ||||
|---|---|---|---|---|---|---|---|---|
| M ± SE × 103 | PR% | M ± SE × 103 | PR% | M ± SE × 103 | PR% | M ± SE × 103 | PR% | |
| GI | 114.6 ± 3.4 aA | 0% | 26.1 ± 1.8 bC | 78% | 23.7 ± 1 cC | 80% | 21.1 ± 1 dC | 83% |
| GII | 115.1 ± 3.2 aA | 0% | 50 ± 1.9 bB | 57% | 48.9 ± 3.2 cB | 59% | 46.1 ± 1.5 dB | 62% |
| GIII | 115.3 ± 3.1 aA | - | 117.1 ± 2.5 aA | - | 118.6 ± 4.3 aA | - | 121.9 ± 4.2 aA | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Wakil, E.S.; Abdelmaksoud, H.F.; Wakid, M.H.; Alsulami, M.N.; Hammam, O.; Albohiri, H.H.; Ghallab, M.M.I. Annona muricata Leaf as an Anti-Cryptosporidial Agent: An In Silico Molecular Docking Analysis and In Vivo Studies. Pharmaceuticals 2023, 16, 878. https://doi.org/10.3390/ph16060878
El-Wakil ES, Abdelmaksoud HF, Wakid MH, Alsulami MN, Hammam O, Albohiri HH, Ghallab MMI. Annona muricata Leaf as an Anti-Cryptosporidial Agent: An In Silico Molecular Docking Analysis and In Vivo Studies. Pharmaceuticals. 2023; 16(6):878. https://doi.org/10.3390/ph16060878
Chicago/Turabian StyleEl-Wakil, Eman S., Hagar F. Abdelmaksoud, Majed H. Wakid, Muslimah N. Alsulami, Olfat Hammam, Haleema H. Albohiri, and Marwa M. I. Ghallab. 2023. "Annona muricata Leaf as an Anti-Cryptosporidial Agent: An In Silico Molecular Docking Analysis and In Vivo Studies" Pharmaceuticals 16, no. 6: 878. https://doi.org/10.3390/ph16060878