
Efficiency of Bone Marrow-Derived Mesenchymal Stem Cells and Hesperetin in the Treatment of Streptozotocin-Induced Type 1 Diabetes in Wistar Rats

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Fasting and 2-h Post-Prandial Glucose Levels at Pre- and Post-Treatment Periods
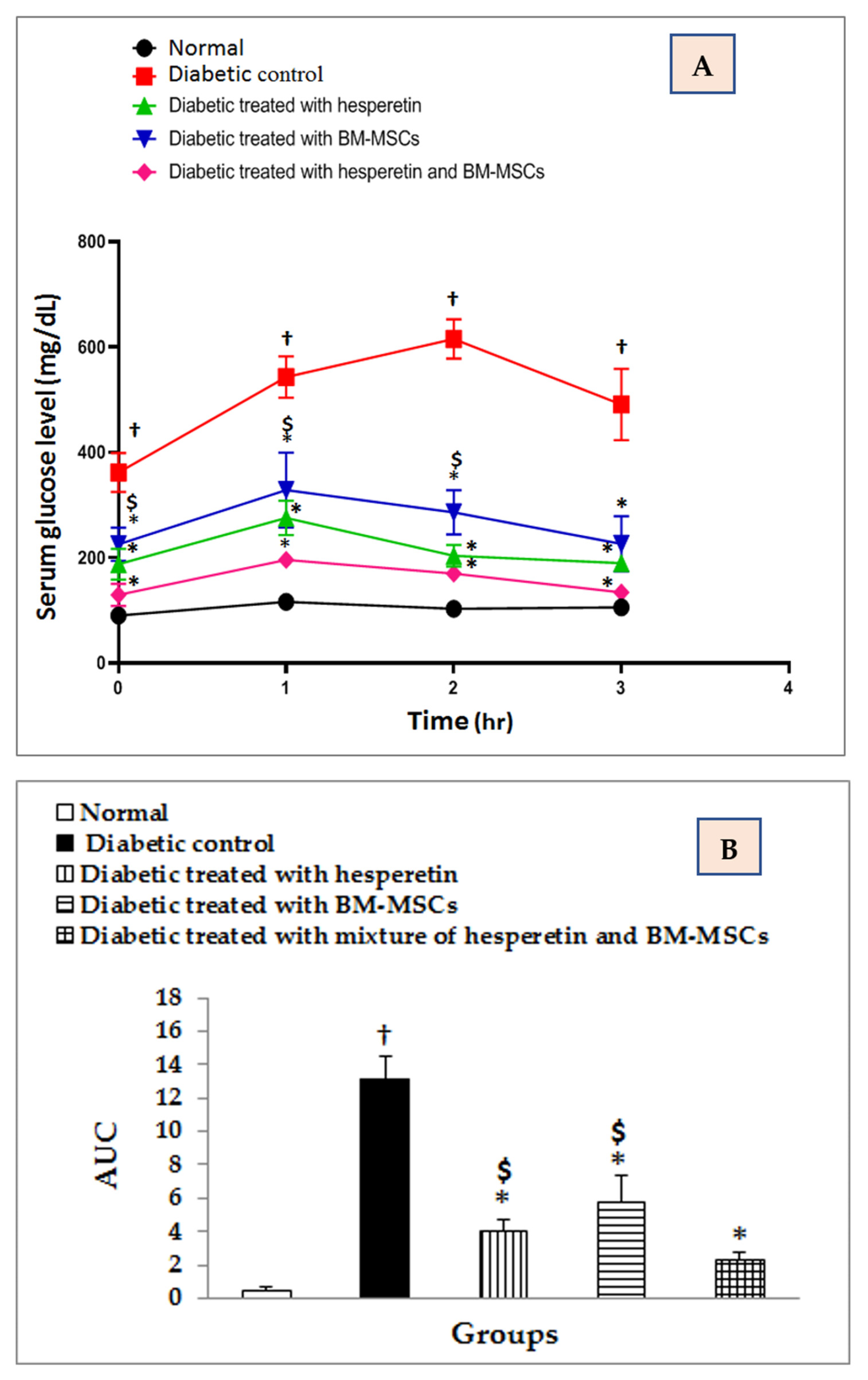
2.2. Evaluation of OGTT
2.3. Effects on Serum Fructosamine Level
2.4. Effects on Serum Insulin and C-Peptide Levels
2.5. Effects on Hepatic Glycogen Levels
2.6. Effects on Hepatic G6Pase and GP Levels
2.7. Effects on Hepatic LPO and GSH and GPx Activities
2.8. Effects on Hepatic GST and GR Activities
2.9. Effects on mRNA Levels of NF-κB, IL-1β and IL-10
2.10. Effects on Serum TNF-α, IL-17 and IL-4 Levels
2.11. Effects on mRNA Levels of P53 and Bcl-2
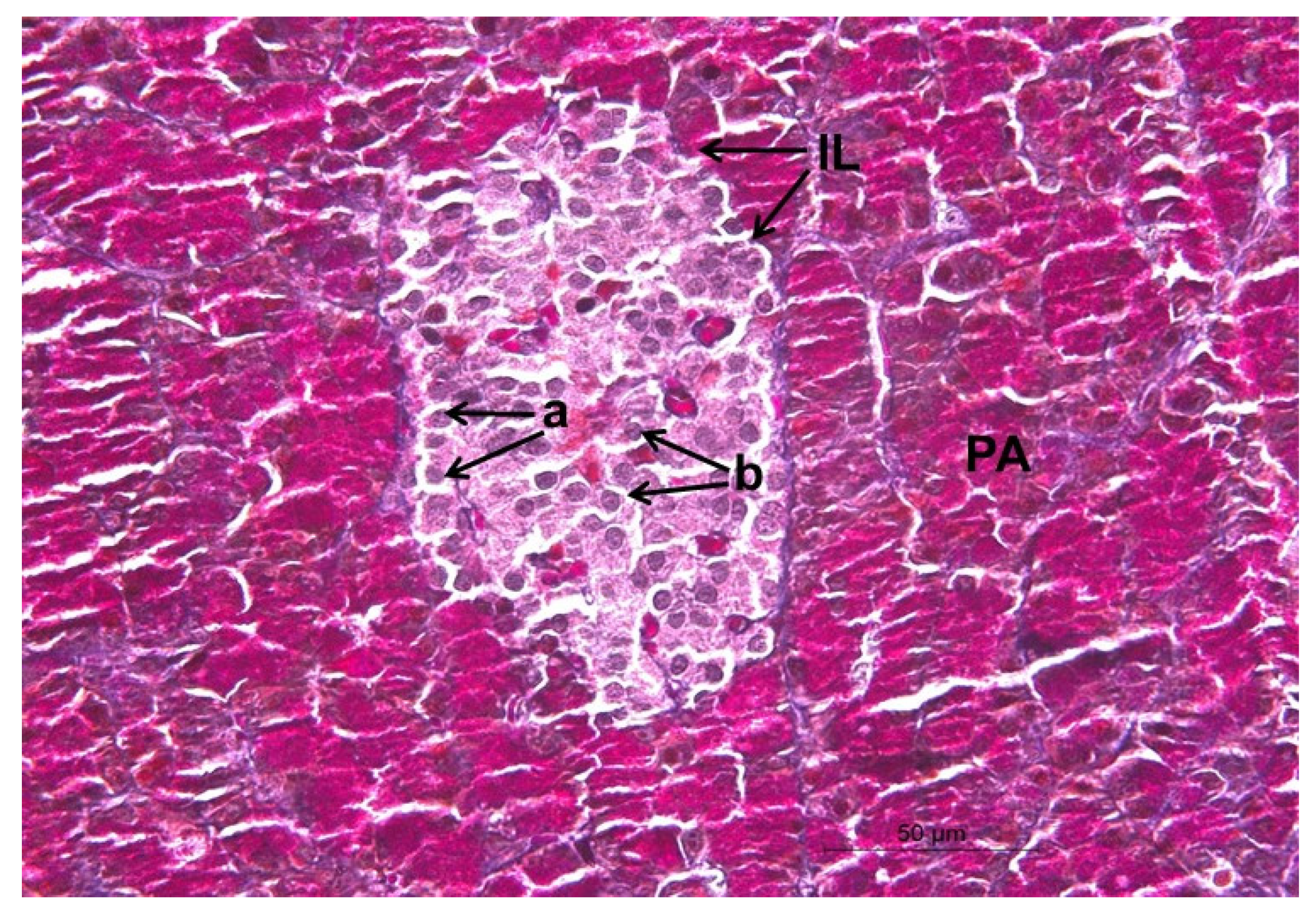
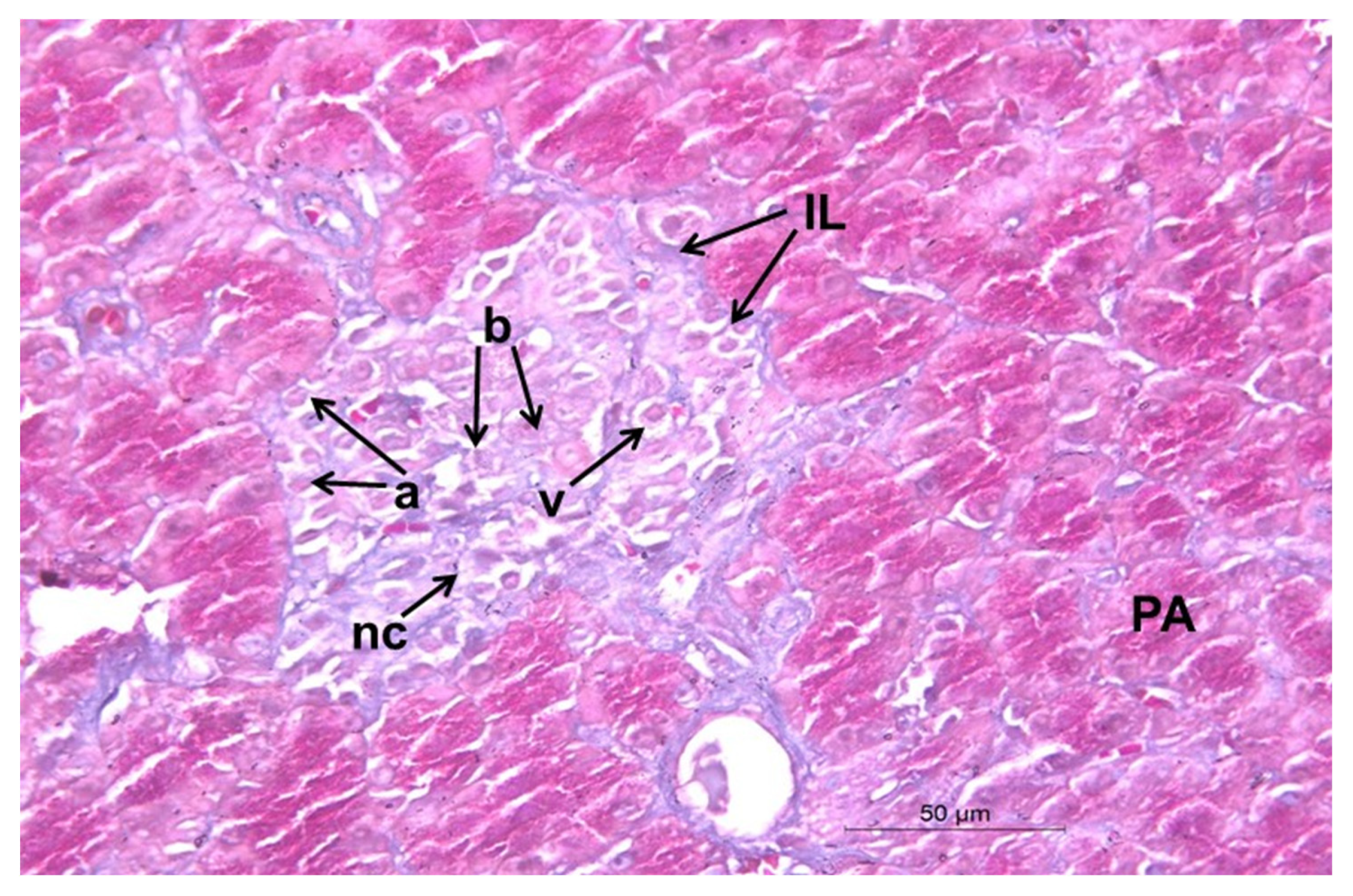
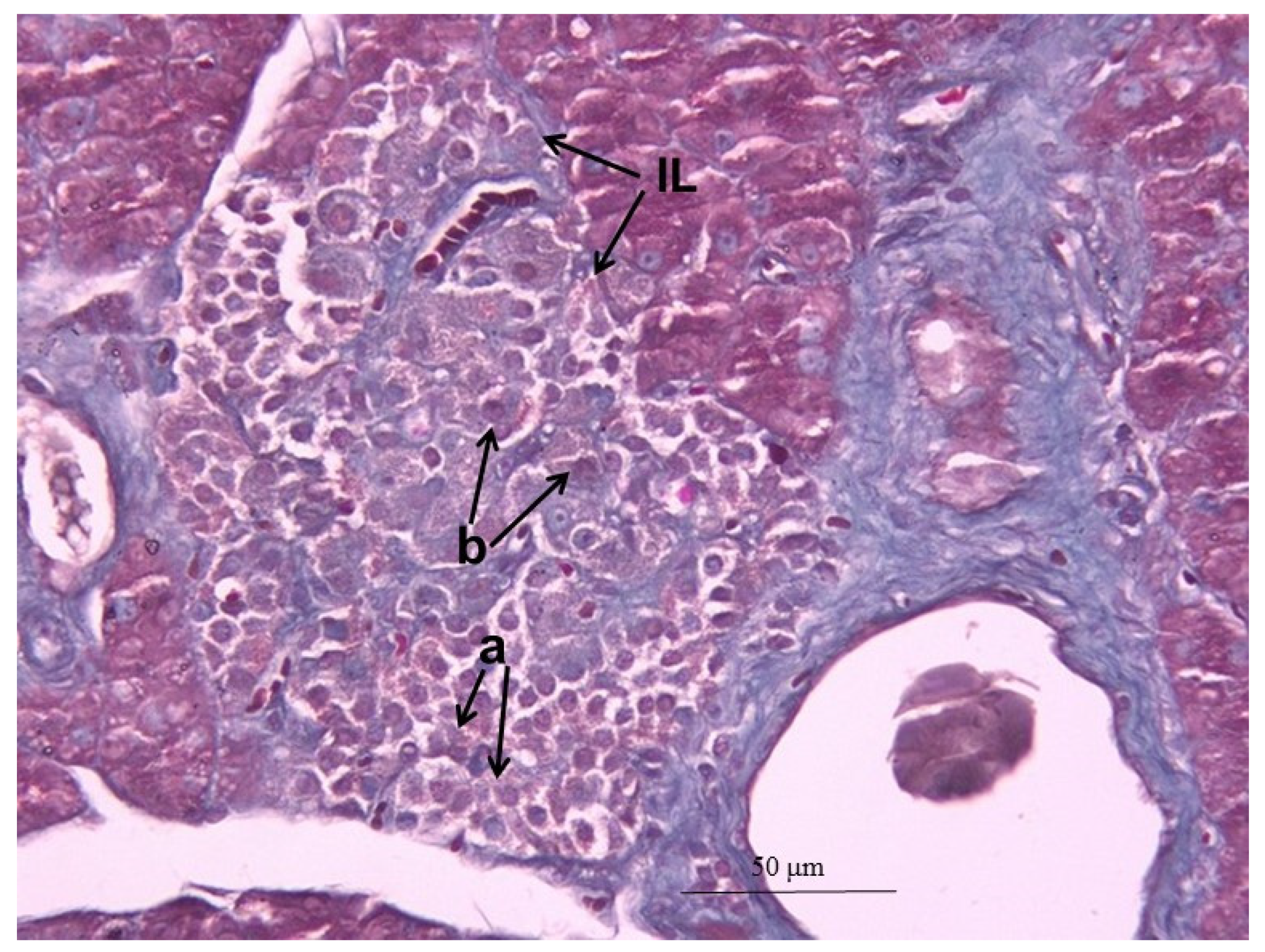
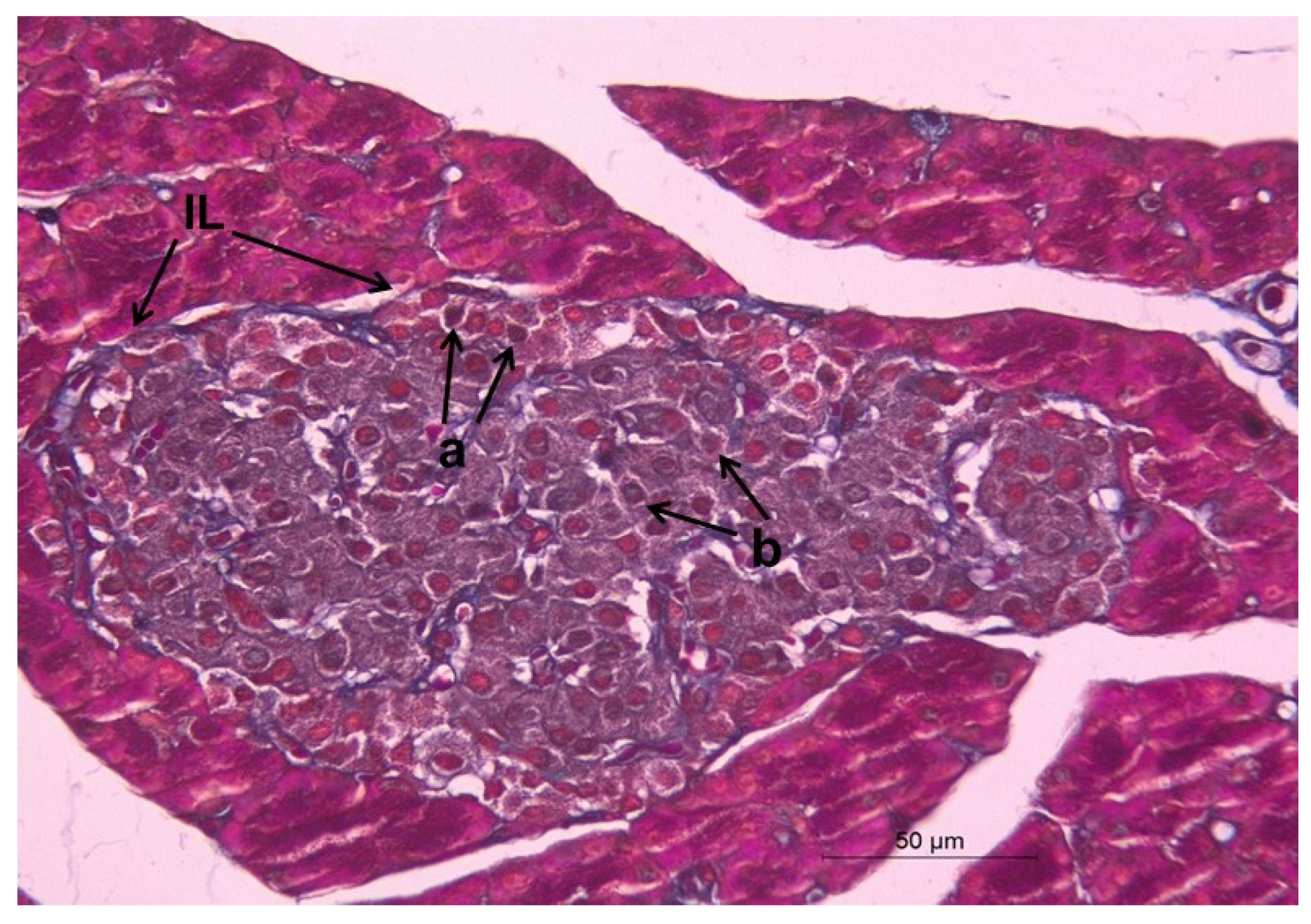
2.12. Histopathological Changes
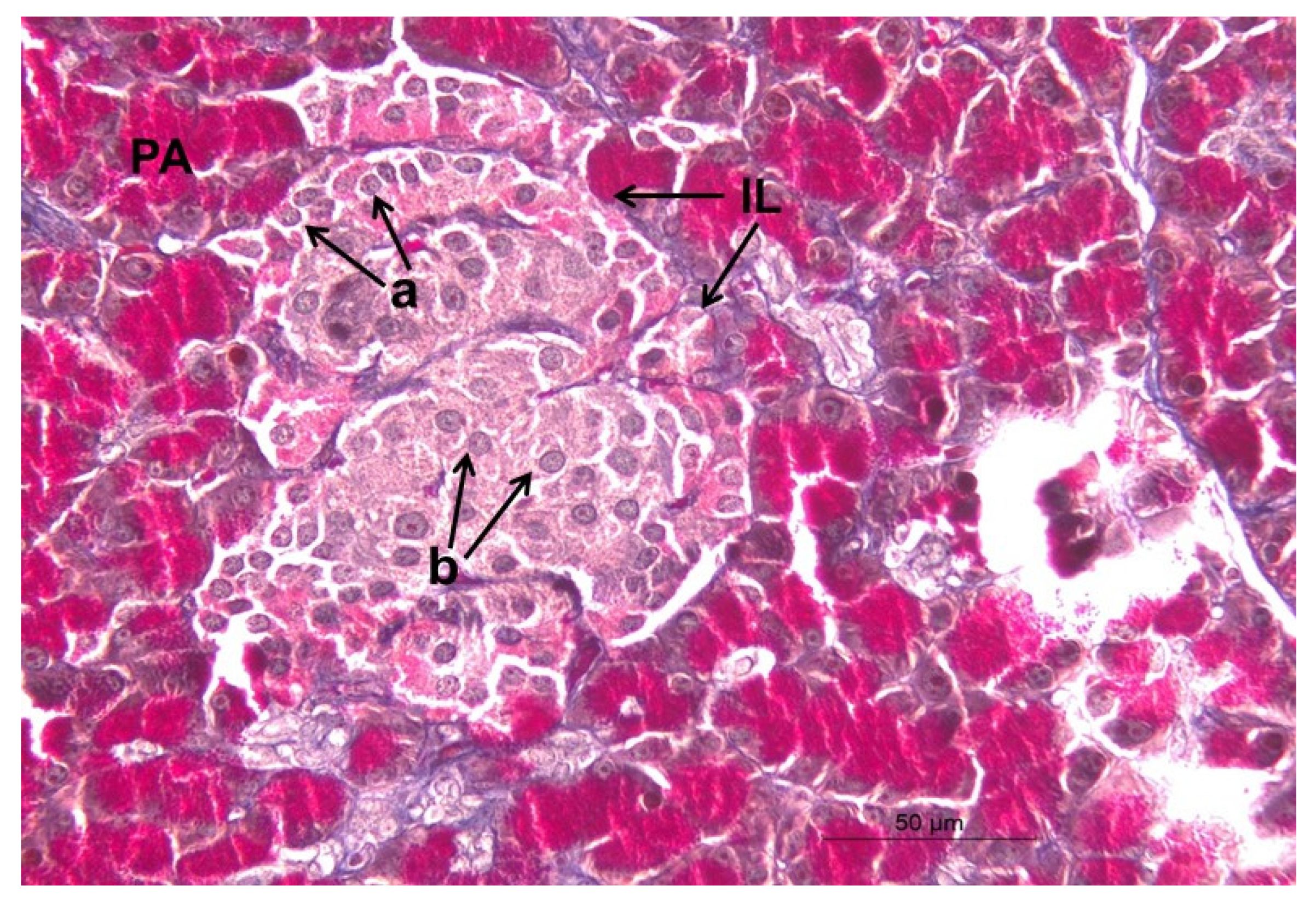
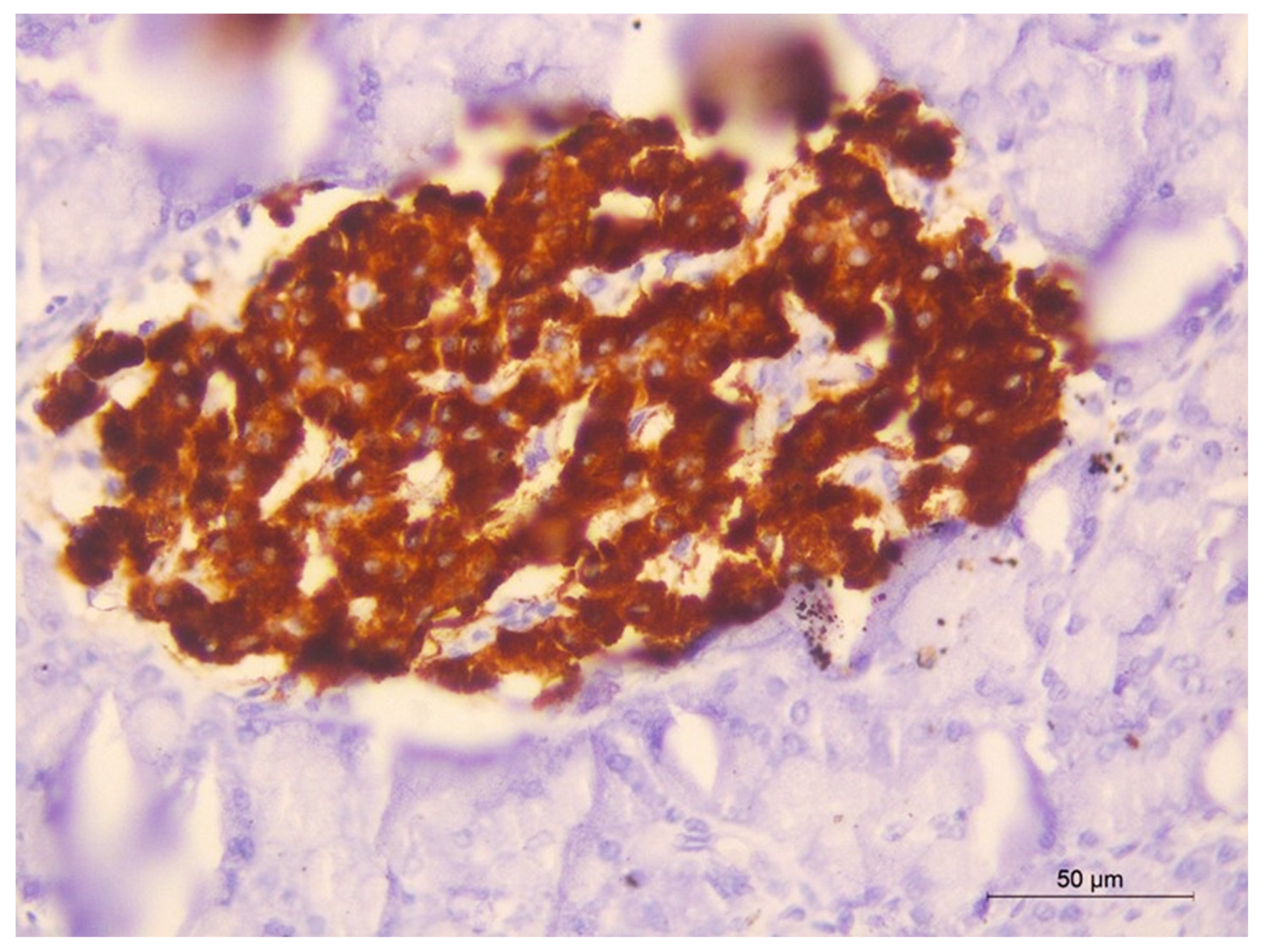
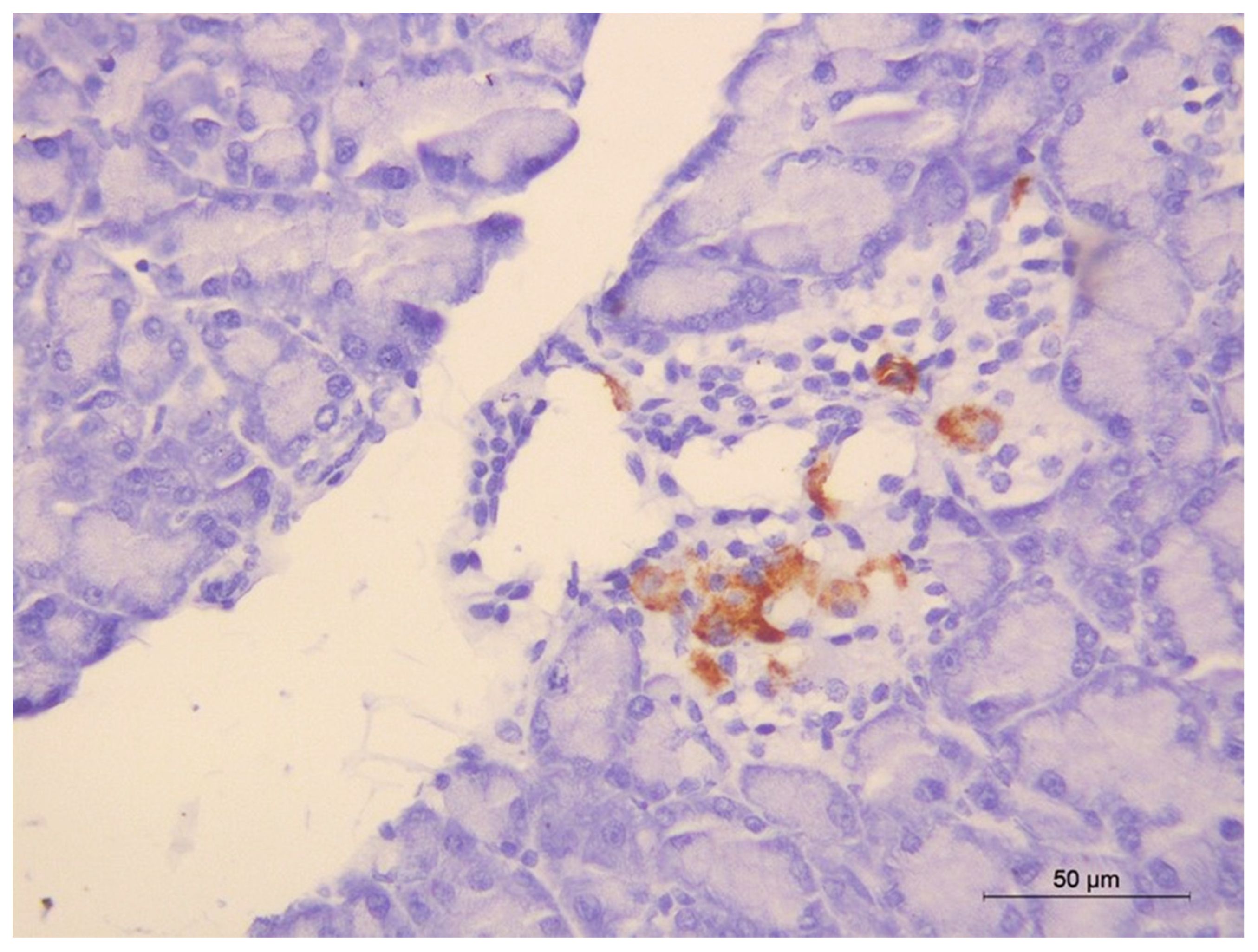
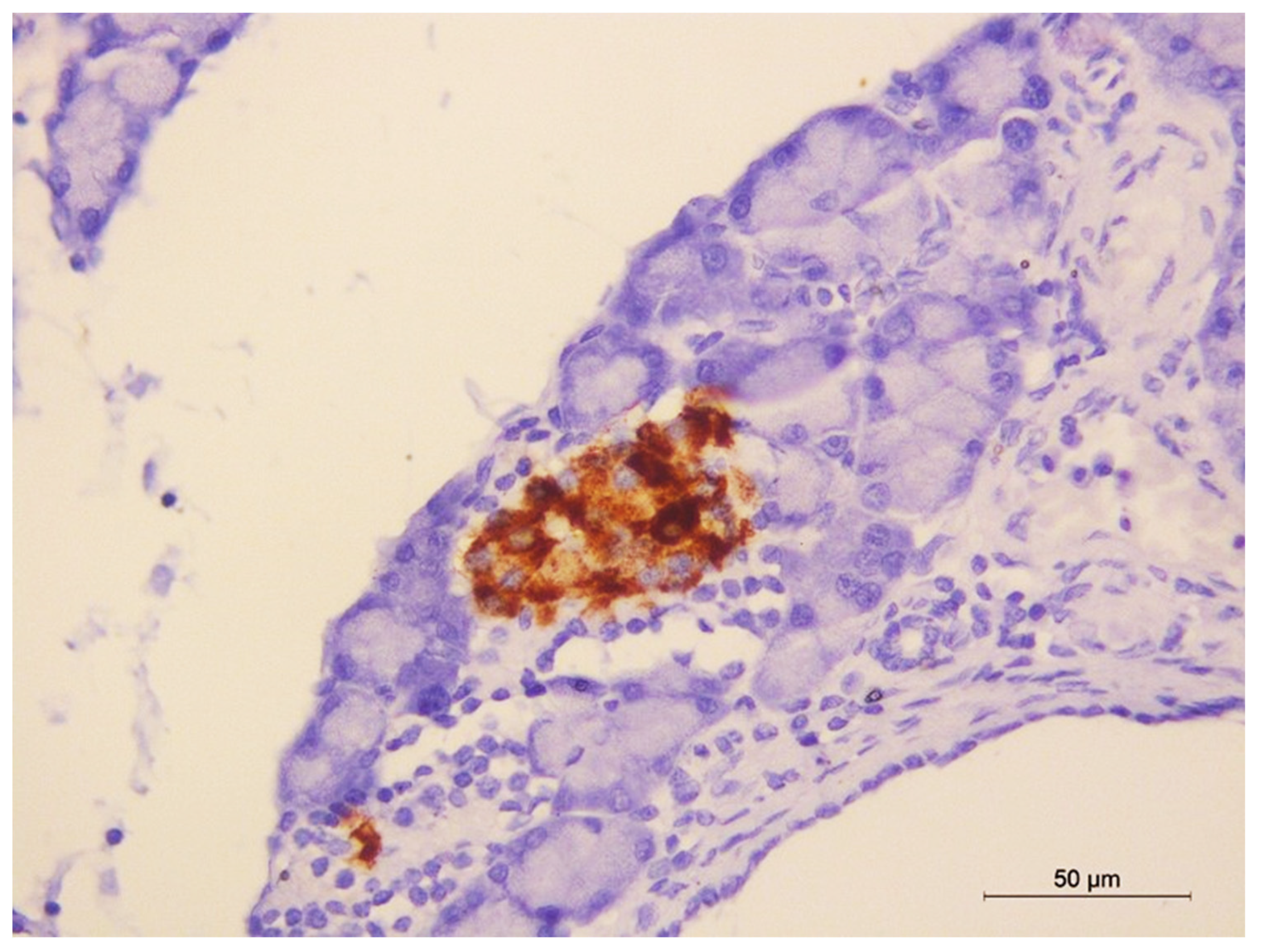
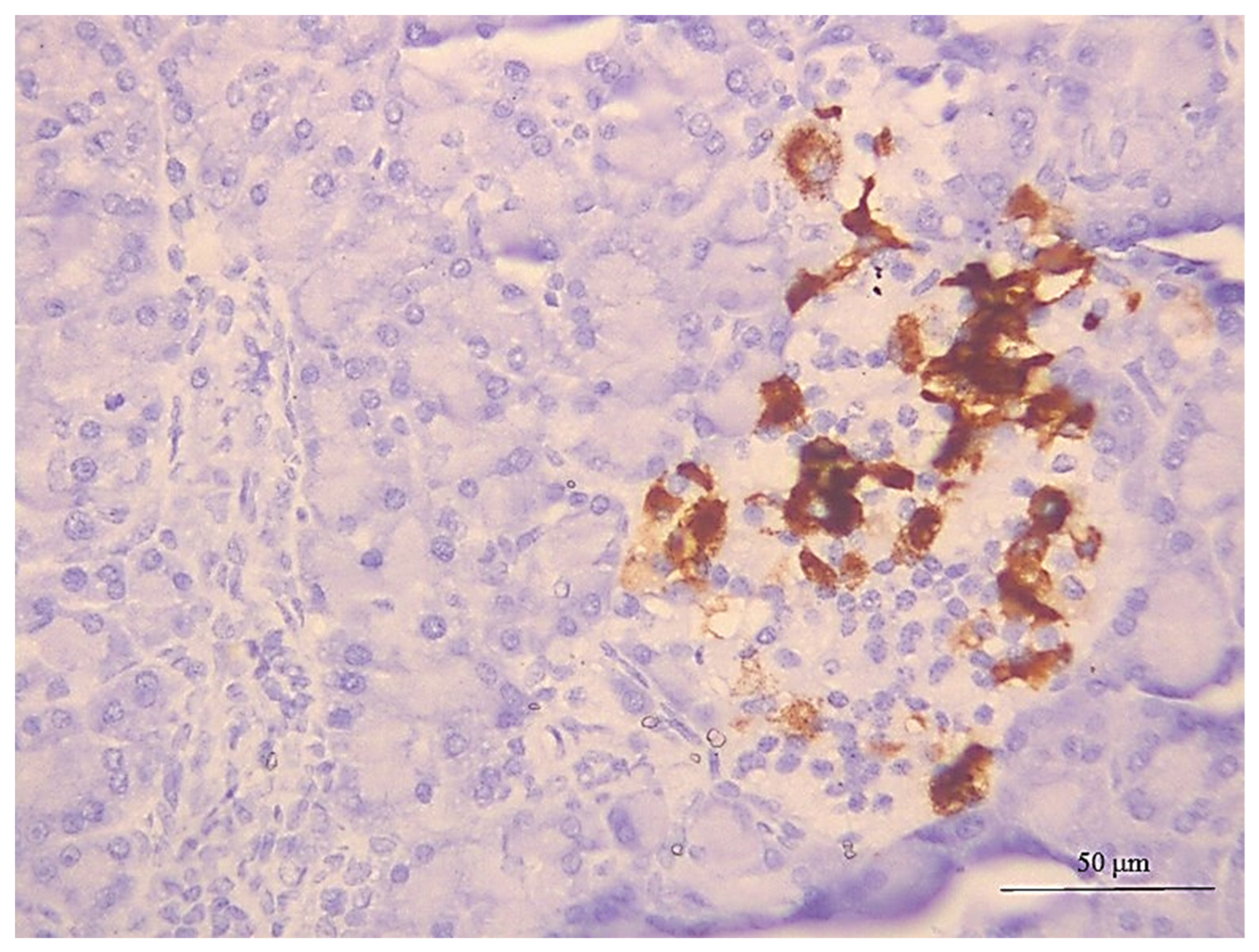
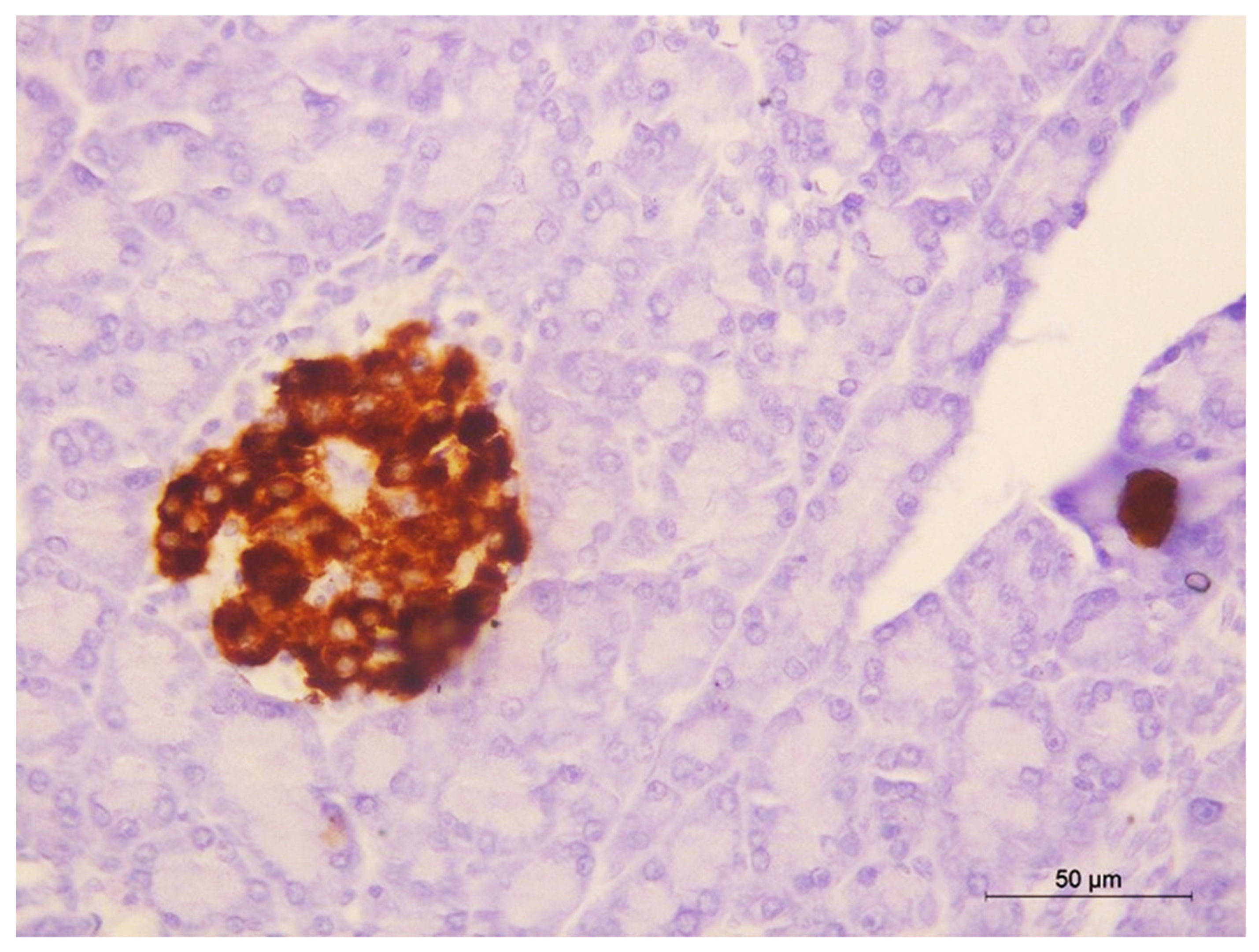
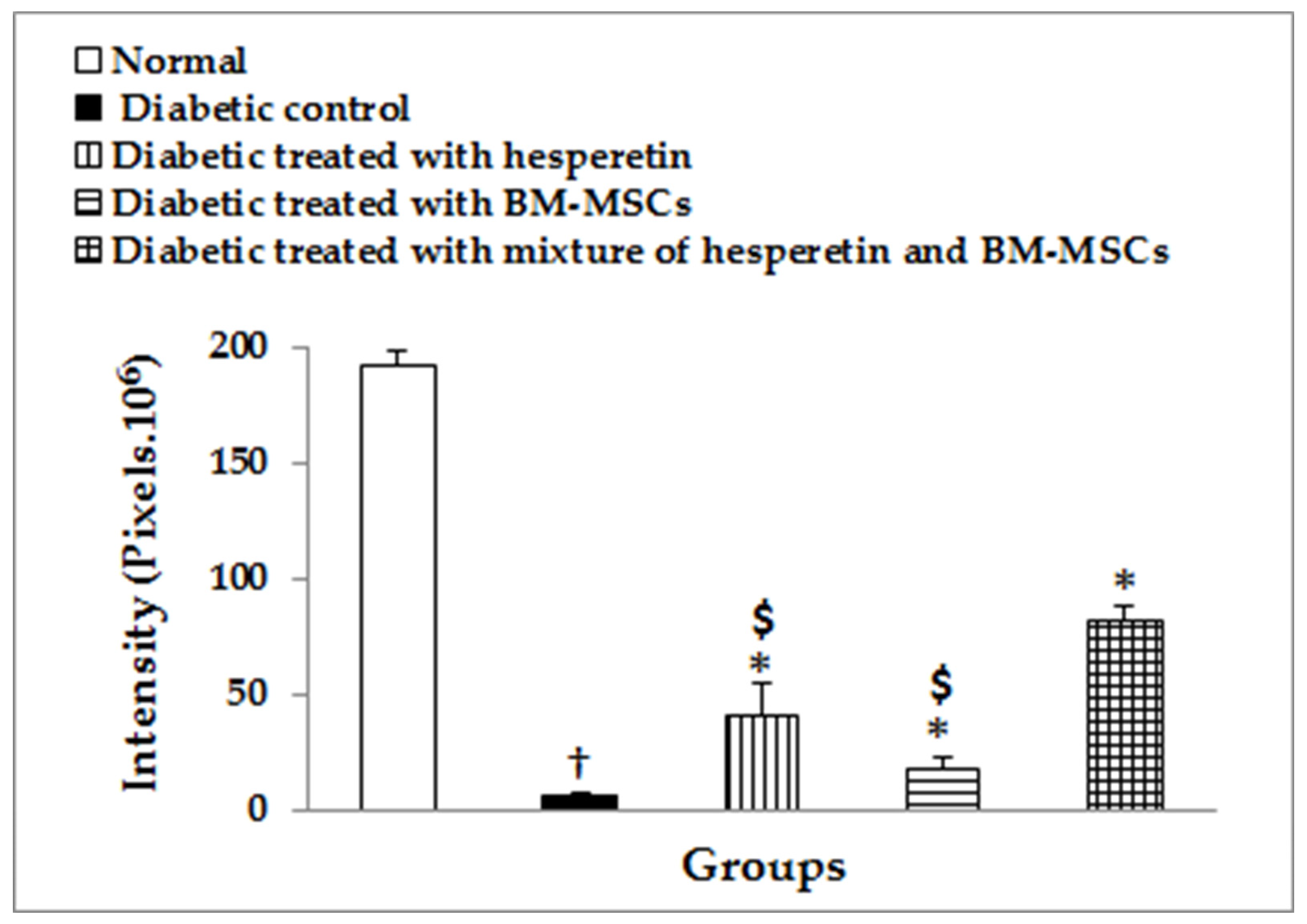
2.13. Immunohistochemical Staining of Insulin
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Preparation of Complete Culture Medium
4.3. Isolation, Culture and Proliferation of BM-MSCs
4.4. Experimental Animals
4.5. Induction of T1DM
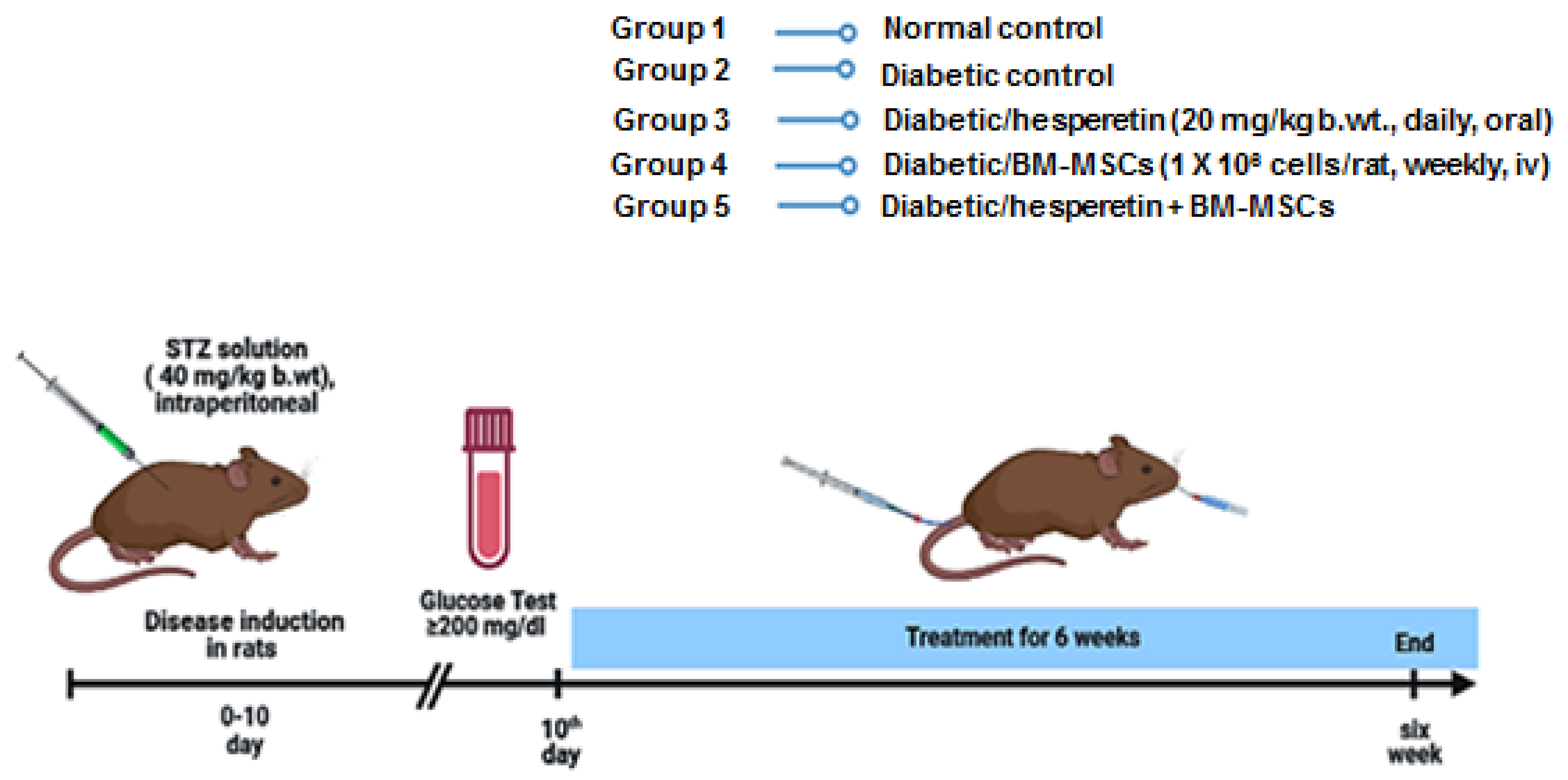
4.6. Animal Grouping and Experimental Design
4.7. Oral Glucose Tolerance Test (OGTT)
4.8. Blood and Tissue Sampling
4.9. Detection of Biochemical Parameters
4.10. Determination of Hepatic Oxidative Stress
4.11. Enzyme-Linked Immunosorbent Assay (ELISA)
4.12. Real-Time qPCR Assay for mRNA Level of NF-κB, IL-1β, IL-10, P53 and Bcl-2
4.13. Histological Preparations
4.14. Immunohistochemistry Assay of Insulin in Pancreatic Islets
4.15. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kodidela, S.; Shaik, F.B.; Chinta, V.; Mohammad, S.A.; Pasala, C.; Mittameedi, C.M.; Maddu, N.; Wudayagiri, R.; Nallanchakravarthula, V. Possible ameliorative role of green tea on chronic alcohol mediated renal toxicity of STZ -induced diabetic rats. Clin. Nutr. Exp. 2020, 34, 1–25. [Google Scholar] [CrossRef]
- Lima Júnior, J.P.; Franco, R.R.; Saraiva, A.L.; Moraes, I.B.; Espindola, F.S. Anacardium Humile St. Hil as a Novel Source of Antioxidant, Antiglycation and α-Amylase Inhibitors Molecules With Potential for Management of Oxidative Stress and Diabetes. J. Ethnopharmacol. 2021, 268, 13667. [Google Scholar] [CrossRef]
- Ozougwu, J.C.; Obimba, K.C.; Belonwu, C.D.; Unakalamba, C.B. The pathogenesis and pathophysiology of type 1 and type 2 diabetes mellitus. J. Physiol. Pathophysiol. 2013, 4, 46–57. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.M.; Nawaz, A.; Evans, M. Drug Therapy in Obesity: A Review of Current and Emerging Treatments. Diabetes Ther. Res. Treat. Educ. Diabetes Relat. Disord. 2020, 11, 1199–1216. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, H.; Sakata, N.; Yoshimatsu, G.; Hasegawa, S.; Kodama, S. Regenerative and Transplantation Medicine: Cellular Therapy Using Adipose Tissue-Derived Mesenchymal Stromal Cells for Type 1 Diabetes Mellitus. J. Clin. Med. 2019, 8, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balamurugan, A.; Bottino, R.; Giannoukakis, N.; Smetanka, C. Prospective and Challenges of Islet Transplantation for the Therapy of Autoimmune Diabetes. Pancreas 2006, 32, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Márquez-Aguirre, A.; Canales-Aguirre, A.; Padilla-Camberos, E.; Esquivel-Solis, H.; Díaz-Martínez, N. Development of the endocrine pancreas and novel strategies for β-cell mass restoration and diabetes therapy. Braz. J. Med. Biol. Res. 2015, 48, 765–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farge, D. Mesenchymal stem cell: Stem cell therapy for type 1diabetes. Proc. Rom. Acad. 2008, 1, 59–70. [Google Scholar]
- Abraham, E.J.; Leech, C.; Lin, J.C.; Zulewski, H.; Habener, J.F. Insulinotropic hormone glucagon-like peptide-1 differentiation of human pancreatic isletderived progenitor cells into insulin producing cells. Endocrinology 2002, 143, 3152–3161. [Google Scholar] [CrossRef] [PubMed]
- Fiorina, P.; Voltarelli, J.; Zavazava, N. Immunological Applications of Stem Cells in Type 1 Diabetes. Endocr. Rev. 2011, 32, 725–754. [Google Scholar] [CrossRef]
- Farooq, T.; Rehman, K.; Hameed, A.; Akash, M.S.H. Stem Cell Therapy and Type 1 Diabetes Mellitus: Treatment Strategies and Future Perspectives. Adv. Exp. Med. Biol. 2018, 1084, 95–107. [Google Scholar]
- Dave, S.D.; Trivedi, H.L.; Gopal, S.C.; Chandra, T. Combined therapy of insulin-producing cells and haematopoietic stem cells offers better diabetic control than only haematopoietic stem cells’ infusion for patients with insulin-dependent diabetes. BMJ Case Rep. 2014, 2014, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drela, K.; Stanaszek, L.; Nowakowski, A.; Kuczynska, Z.; Lukomska, B. Experimental Strategies of Mesenchymal Stem Cell Propagation: Adverse Events and Potential Risk of Functional Changes. Stem Cells Int. 2019, 2019, 7012692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, O.M.; Sayed, H.M. Stem Cell Therapies in Regenerative Medicine and Diabetes Mellitus: Advances, Constraints and Future Prospects. J. Transpl. Stem Cell Biol. 2016, 3, 22. [Google Scholar]
- Birtwistle, L.; Chen, X.-M.; Pollock, C. Mesenchymal Stem Cell-Derived Extracellular Vesicles to the Rescue of Renal Injury. Int. J. Mol. Sci. 2021, 22, 6596. [Google Scholar] [CrossRef]
- Si, Y.; Zhao, Y.; Hao, H.; Liu, J.; Guo, Y.; Mu, Y.; Sen, J.; Cheng, C.; Fu, X.; Han, W. Infusion of Mesenchymal Stem Cells Ameliorates Hyperglycemia in Type 2 Diabetic Rats: Identification of a Novel Role in Improving Insulin Sensitivity. Diabetes 2012, 61, 1616–1625. [Google Scholar] [CrossRef] [Green Version]
- Sudkamp, N.P. Clinical applications of Mesenchymal Stem Cells. Eur. Cells Mater. 2007, 13, 3. [Google Scholar]
- Nadri, S.; Soleimani, M.; Hosseni, R.H.; Massumi, M.; Atashi, A.; Izadpanah, R. An efficient method for isolation of murine bone marrow mesenchymal stem cells. Int. J. Dev. Biol. 2007, 51, 723–729. [Google Scholar] [CrossRef]
- Sayed, H.M.; Awaad, A.S.; Abdel Rahman, F.E.-Z.S.; Al-Dossari, M.; Abd El-Gawaad, N.S.; Ahmed, O.M. Combinatory Effect and Modes of Action of Chrysin and Bone Marrow-Derived Mesenchymal Stem Cells on Streptozotocin/Nicotinamide-Induced Diabetic Rats. Pharmaceuticals 2023, 16, 34. [Google Scholar] [CrossRef]
- Morin, B.; Nichols, L.A.; Zalasky, K.M.; Davis, J.W.; Manthey, J.A.; Holland, L.J. The Citrus Flavonoids Hesperetin and Nobiletin Differentially Regulate Low Density Lipoprotein Receptor Gene Transcription in HepG2 Liver Cells 3. J. Nutr. 2008, 138, 1274–1281. [Google Scholar] [CrossRef] [Green Version]
- Aranganathan, S.; Nalini, N. Efficacy of the potential chemopreventive agent, hesperetin (citrus flavanone), on 1,2-dimethylhydrazine induced colon carcinogenesis. Food Chem. Toxicol. 2009, 47, 2594–2600. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, R.; Subramani, S.; Abdullah, S.H.S.; Udaiyar, M. Antihyperglycemic effect of hesperetin, a citrus flavonoid, extenuates hyperglycemia and exploring the potential role in antioxidant and antihyperlipidemic in streptozotocin-induced diabetic rats. Biomed. Pharmacother. 2018, 97, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, Y.; Xu, S.; Ren, J.; Tang, L.; Gong, J.; Lin, Y.; Fang, H.; Su, D. Hesperetin, a Promising Treatment Option for Diabetes and Related Complications: A Literature Review. J. Agric. Food Chem. 2022, 70, 8582–8592. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.J. Antioxidative effects of hesperetin against 7, 12-dimethylbenz (a) anthracene-induced oxidative stress in mice. Life Sci. 2008, 82, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.L.; Yen, G.C. Neuroprotective effects of the citrus flavanones against H2O2-induced cytotoxicity in PC12 cells. J. Agric. Food Chem. 2008, 56, 859–864. [Google Scholar] [CrossRef]
- Hirata, A.; Murakami, Y.; Shoji, M.; Kadoma, Y.; Fujisawa, S. Kinetics of radical-scavenging activity of hesperetin and hesperidin and their inhibitory activity on COX-2 expression. Anticancer Res. 2005, 25, 3367–3374. [Google Scholar]
- Yumol, J.L.; Ward, W.E. The Polyphenolic Compound Hesperidin and Bone Protection. In Polyphenols: Mechanisms of Action in Human Health and Disease; Academic Press: New York, NY, USA, 2018; pp. 431–440. [Google Scholar] [CrossRef]
- Ahmed, O.M.; Hassan, M.A.; Saleh, A.S. Combinatory effect of hesperetin and mesenchymal stem cells on the deteriorated lipid profile, heart and kidney functions and antioxidant activity in STZ-induced diabetic rats. Biocell 2020, 44, 27–29. [Google Scholar] [CrossRef]
- Revathy, J.; Abdullah, S.S. The Role of Hesperetin in the Management of Diabetes Mellitus and Its Complications. J. Cancer Treat. Res. 2017, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-J. Protective effects of Chrysanthemi Flos extract against streptozotocin-induced oxidative damage in diabetic mice. J. Med. Plants Res. 2012, 6, 622–630. [Google Scholar] [CrossRef]
- Eze, E.; Mohammed, A.; Musa, K.; Tanko, Y.; Isa, A. Effect of Ethanolic Leaf Extract of Mucunapruriens (fabaceae) on Lipid Profile inAlloxan-Induced Diabetic Wistar rats. Br. J. Pharmacol. Toxicol. 2012, 3, 102–109. [Google Scholar]
- WHO. Use of Glycated Haemoglobin (HbA1c) in the Diagnosis of Diabetes Mellitus: Abbreviated Report of a WHO Consultation. Approved by the Guidelines Review Committee; World Health Organization: Geneva, Switzerland, 2011; pp. 299–309. [Google Scholar]
- Laakso, M. Heart in Diabetes: A Microvascular Disease. Diabetes Care 2011, 34, S145–S149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Q.; Huang, Y.; Liu, J. Mesenchymal Stem Cells: An Excellent Candidate for the Treatment of Diabetes Mellitus. Int. J. Endocrinol. 2021, 2021, 9938658. [Google Scholar] [CrossRef] [PubMed]
- Abdel Aziz, M.T.; Wassef, M.A.; Ahmed, H.H.; Rashed, L.; Mahfouz, S.; Aly, M.I.; Hussein, R.E. The role of bone marrow derived mesenchymal stem cells in attenuation of kidney function in rats with diabetic nephropathy. Diabetol. Metab. Syndr. 2014, 6, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira, A.; Kahlenberg, S.; Hornsby, P. Therapeutic potential of mesenchymal stem cells for diabetes. J. Mol. Endocrinol. 2017, 59, R109–R120. [Google Scholar] [CrossRef] [Green Version]
- Su, T.; Xiao, Y.; Xiao, Y.; Guo, Q.; Li, C.; Huang, Y.; Deng, Q.; Wen, J.; Zhou, F.; Luo, X.-H. Bone Marrow Mesenchymal Stem Cells-Derived Exosomal MiR-29b-3p Regulates Aging-Associated Insulin Resistance. ACS Nano 2019, 13, 2450–2462. [Google Scholar] [CrossRef]
- El Barky, A.R.; Ezz, A.A.; Abd-Elhameed, A.; Hussein, S.A.; Hafez, Y.A.; Mohamed, T.M. Can Stem Cells Ameliorate the Pancreatic Damage Induced by Streptozotocin in Rats? Can. J. Diabetes 2018, 42, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Lock, L.T.; Tzanakakis, E.S.; Duffy, C.; Prugue, C.; Glew, R.; Smith, T.; Howell, C.; Choi, G.; Cook, A.D.; Schmidt, J.J.; et al. Stem/Progenitor Cell Sources of Insulin-Producing Cells for the Treatment of Diabetes. Tissue Eng. 2007, 13, 1399–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberti, K.G.; Zimmet, P.Z. Definition, diagnosis and classification of diabetes mellitus and its complications. Part 1: Diagnosis and classification of diabetes mellitus. Provisional report of a WHO Consultation. Diabetes Med. 1998, 15, 539–553. [Google Scholar] [CrossRef]
- Ahmed, O.M.; Abdel-Hamid, H.; Bastawy, M.; Hasona, N. Antihyperglycemic effects of Plantago ispaghula seeds aqueous extracts in diabetic and hypercholesterolemic rats. J. Egypt. Ger. Soc. Zool. 2006, 51, 371–393. [Google Scholar]
- Gad, S.B.; El-Maddawy, Z.K. Silymarin improves pancreatic endocrine function in rats. Pak. Vet. J. 2013, 34, 214–218. [Google Scholar]
- Kamal, N.M. Role of Hormone Receptor in the Aetiology of Insulin Dependent and Non-Insulin Dependent Diabetes Mellitus. Master’s Thesis, Faculty of Medicine, Cairo University, Cairo, Egypt, 1991. [Google Scholar]
- Dutta, P.; Bhansali, S.; Kumar, V.; Saikia, U.N.; Medhi, B.; Jha, V.; Bhansali, A. Effect of mesenchymal stem cells transplantation on glycaemic profile & their localization in streptozotocin induced diabetic Wistar rats. Indian J. Med Res. 2015, 142, 63–71. [Google Scholar]
- Hess, D.; Li, L.; Martin, M.; Sakano, S.; Hill, D.; Strutt, B.; Thyssen, S.; A Gray, D.; Bhatia, M. Bone marrow–derived stem cells initiate pancreatic regeneration. Nat. Biotechnol. 2003, 21, 763–770. [Google Scholar] [CrossRef]
- Bell, G.I.; Broughton, H.C.; Levac, K.D.; Allan, D.A.; Xenocostas, A.; Hess, D.A. Transplanted Human Bone Marrow Progenitor Subtypes Stimulate Endogenous Islet Regeneration and Revascularization. Stem Cells Dev. 2012, 21, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.-Y.; Chen, L.; Gao, G.-Q.; Wang, L.; Song, J.; Chen, B.; Xu, Y.-X.; Sun, L. Allogeneic diabetic mesenchymal stem cells transplantation in streptozotocin-induced diabetic rat. Clin. Investig. Med. 2008, 31, E328–E337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hindle, E.J.; Rostron, G.M.; Clarck, S.A.; Gatt, J.A. Serum fructosamine and glycated hemoglobin measurements in diabetic control. Arch. Dis. Child 1986, 61, 113–117. [Google Scholar] [CrossRef] [Green Version]
- Donnelly, J.G. Carbohydrates and alterations in glucose metabolism. In Clinical Chemistry; Principles, Procedures, Correlations; Bishop, M.L., Fody, E.P., Eds.; Lippincott: Philadelphia, PA, USA, 1996; pp. 308–309. [Google Scholar]
- Ahmed, O.M.; Abdel-Moneim, A.; Yazid, I.A.; Mahmoud, A.M. Antihyperglycemic, antihyperlipidemic and antioxidant effects and the probable mechanisms of action of ruta graveolens infusion and rutin in nicotinamide streptozotocin-induced diabetic rats. Diabetol. Croat. 2010, 27, 39–41. [Google Scholar]
- Rajamani, U.; Joseph, D.; Roux, S.; Essop, F. The hexosamine biosynthetic pathway can mediate myocardial apoptosis in a rat model of diet-induced insulin resistance. Acta Physiol. 2011, 202, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Revathy, J.; Sheik Abdullah, S. Influence of hesperetin on glycoprotein components in diabetic rats. Int. J. Sci. Eng. Res. 2016, 7, 214–220. [Google Scholar]
- Farid, A.; Haridyy, H.; Ashraf, S.; Ahmed, S.; Safwat, G. Co-treatment with grape seed extract and mesenchymal stem cells in vivo regenerated beta cells of islets of Langerhans in pancreas of type I-induced diabetic rats. Stem Cell Res Ther. 2022, 13, 28. [Google Scholar] [CrossRef]
- Ahmed, O.M.; Ali, T.M.; Abdel Gaid, M.A.; Elberry, A.A. Effects of enalapril and paricalcitol treatment on diabetic nephropathy and renal expressions of TNF-α, p53, caspase-3 and Bcl-2 in STZ-induced diabetic rats. PLoS ONE 2019, 14, e0214349. [Google Scholar] [CrossRef] [Green Version]
- Akhani, S.P.; Vishwakarma, S.L.; Goyal, R.K. Antidiabetic activity of Zingiber officinal in streptozotocin-induced type 1 diabetic rats. J. Pharm. Pharmacol. 2004, 56, 101–105. [Google Scholar] [CrossRef]
- Ahmed, O.M. Anti-hyperglycemic, immunomodulatory and anti-oxidant efficacy of vasoactive intestinal peptide in streptozotocin-induced diabetic mice. Int. J. Zool. Res. 2009, 2, 42–61. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, A.M.; Ahmed, O.M.; Ashour, M.B.; Abdel-Moneim, A. In vivo and in vitro antidiabetic effects of citrus flavonoids; a study on the mechanism of action. Int. J. Diabetes Dev. Ctries. 2015, 35, 250–263. [Google Scholar] [CrossRef]
- Kappel, V.D.; Frederico, M.J.; Postal, B.G.; Mendes, C.P.; Cazarolli, L.H.; Silva, F.R. The role of calcium in intracellular pathways of rutin in rat pancreatic islets: Potential insulin secretagogue effect. Eur. J. Pharmacol. 2013, 702, 264–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurewicz, M.; Yang, S.; Augello, A.; Godwin, J.G.; Moore, R.F.; Azzi, J.; Fiorina, P.; Atkinson, M.; Sayegh, M.H.; Abdi, R. Congenic Mesenchymal Stem Cell Therapy Reverses Hyperglycemia in Experimental Type 1 Diabetes. Diabetes 2010, 59, 3139–3147. [Google Scholar] [CrossRef] [Green Version]
- Ezquer, F.; Ezquer, M.; Simon, V.; Conget, P. The Antidiabetic Effect of MSCs Is Not Impaired by Insulin Prophylaxis and Is Not Improved by a Second Dose of Cells. PLoS ONE 2011, 6, e16566. [Google Scholar] [CrossRef] [Green Version]
- Grover, J.; Vats, V.; Rathi, S. Anti-hyperglycemic effect of Eugenia jambolana and Tinospora cordifolia in experimental diabetes and their effects on key metabolic enzymes involved in carbohydrate metabolism. J. Ethnopharmacol. 2000, 73, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, O.M. The hypoglycemic effect of curcumin and esculetin and their probable mechanisms of action in streptozotocin diabetic albino rats. J. Egypt Ger. Soc. Zool. 2005, 46, 351–375. [Google Scholar]
- Abdel-Moneim, A.; Ahmed, O.M.; Rawi, S.M.; Semmler, M. Studies on the hypoglycemic and hypolipidemic effects of glimepiride and some antidiabetic plants on streptozotocin diabetic rats. J. Egypt Ger. Soc. Zool. 2001, 34, 175–206. [Google Scholar]
- Raju, J.; Gupta, D.; Rao, A.R.; Yadav, P.K.; Baquer, N.Z. Trigonella foenum graecum (fenugreek) seed powder improves glucose homeostasis in alloxan diabetic rat tissues by reserving the altered glycolytic, gluconeogenic and lipogenic enzymes. Mol. Cell Biochem. 2001, 224, 45–51. [Google Scholar] [CrossRef]
- Ahmed, O.M. Evaluation of the antihyperglycemic, antihyperlipidemic and myocardial enhancing properties of pioglitazone in diabetic and hyperthyroid rats. J. Egypt. Ger. Soc. Zool. 2006, 51, 253–278. [Google Scholar]
- Bollen, M.; Keppens, S.; Stalmans, W. Specific features of glycogen metabolism in the liver. Biochem. J. 1998, 336, 19–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, T.; Amrein, H. Gluconeogenes is an ancient biochemical pathway with a new twist. Fly 2017, 25, 1–6. [Google Scholar]
- Jung, U.J.; Lee, M.K.; Jeong, K.S.; Choi, M.-S. The hypoglycemic effects of hesperidin and naringin are partly mediated by hepatic glucose-regulating enzymes in C57BL/KsJ-db/db mice. J. Nutr. 2004, 134, 2499–2503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constantin, R.P.; Constantin, R.P.; Bracht, A.; Yamamoto, N.S.; Ishii-Iwamoto, E.-L.; Constantin, J. Molecular mechanisms of citrus flavanones on hepatic gluconeogenesis. Fitoterapia 2014, 92, 148–162. [Google Scholar] [CrossRef]
- Villar-Palasí, C.; Guinovart, J.J. The role of glucose 6-phosphate in the control of glycogen synthase. FASEB J. 1997, 11, 544–548. [Google Scholar] [CrossRef]
- Shaban, N.Z.; El-Kersh, M.A.; El-Rashidy, F.H.; Habashy, N.H. Protective role of Punica granatum (pomegranate) peel and seed oil extracts on diethylnitrosamine and phenobarbital-induced hepatic injury in male rats. Food Chem. 2013, 141, 1587–1596. [Google Scholar] [CrossRef]
- Stadler, K.; Jenei, V.; von Bölcsházy, G.; Somogyi, A.; Jakus, J. Increased nitric oxide levels as an early sign of premature aging in diabetes. Free. Radic. Biol. Med. 2003, 35, 1240–1251. [Google Scholar] [CrossRef]
- Ling, P.-R.; Mueller, C.; Smith, R.J.; Bistrian, B.R. Hyperglycemia induced by glucose infusion causes hepatic oxidative stress and systemic inflammation, but not STAT3 or MAP kinase activation in liver in rats. Metabolism 2003, 52, 868–874. [Google Scholar] [CrossRef]
- Kume, E.; Fujimura, H.; Matsuki, N.; Ito, M.; Aruga, C.; Toriumi, W.; Kitamura, K.; Doi, K. Hepatic changes in the acute phase of streptozotocin (SZ)-induced diabetes in mice. Exp. Toxicol. Pathol. 2004, 55, 467–480. [Google Scholar] [CrossRef]
- Pasaoglu, H.; Sancak, B.; Bukan, N. Lipid Peroxidation and Resistance to Oxidation in Patients with Type 2 Diabetes Mellitus. Tohoku J. Exp. Med. 2004, 203, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef]
- Rajasekaran, S.; Sivagnanam, K.; Subramanian, S. Antioxidant effect of Aloe vera gel extract in streptozotocin-induced diabetes in rats. Pharmacol. Rep. 2005, 57, 90–96. [Google Scholar] [PubMed]
- Ugwu, M.N.; Umar, I.A.; Utu-Baku, A.B.; Dasofunjo, K.; Ukpanukpong, R.U.; Yakubu, O.E.; Okafor, A. Antioxidant Status and Organ Function in Streptozotocin-Induced Diabetic Rats Treated with Aqueous, Methanolic and Petroleum Ether Extracts of Ocimum. J. Appl. Pharm. Sci. 2013, 3, 75–79. [Google Scholar]
- Narenjkar, J.; Roghani, M.; Shekouhi, M. The effect of hesperetin on serum level of aspartate and alanine amoinotransferase and hepatic and cardiac level of malondialdehyde in diabetic rats. Daneshvar Med. 2010, 18, 33–42. [Google Scholar]
- Fang, Y.; Shuling, B.X.T.; Fan, J.; Tian, X.; Hou, W.; Tong, H.; Li, D. Autologous transplantation of adipose derived mesenchymal stem cells ameliorates streptozotocin-induced diabetic nephropathy in rats by inhibiting oxidative stress, pro-inflammatory cytokines and the p38 MAPK signaling pathway. Int. J. Mol. Med. 2012, 30, 85–92. [Google Scholar] [PubMed]
- Sheweita, S.A.; Mashaly, S.; Newairy, A.A.; Abdou, H.M.; Eweda, S.M. Changes in Oxidative Stress and Antioxidant Enzyme Activities in Streptozotocin-Induced Diabetes Mellitus in Rats: Role of Alhagi maurorum Extracts. Oxidative Med. Cell. Longev. 2016, 2016, 5264064. [Google Scholar] [CrossRef] [Green Version]
- Gokce, G.; Haznedaroglu, M.Z. Evaluation of antidiabetic, antioxidant and vasoprotective effects of Posidonia oceanica extract. J. Ethnopharmacol. 2008, 115, 122–130. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, J.; Wang, J.; Pang, X.; Zhuang, X.; Zhu, X.; Qu, W. Hypoglycemic effect of aqueous extract of sea buckthorn (Hippophae rhamnoides L.) seed residues in streptozotocin induced diabetic rats. Phytother. Res. 2010, 24, 228–232. [Google Scholar] [CrossRef]
- Zahran, F.M.; Al-Hussaini, A.S.; Abou-Mesalam, A.I. Hepatoprotective effect of cinnamaldehyde and hesperedin mixture on induced type-1- diabetic rats. Int. J. Biol. Pharm. Res. 2016, 7, 74–82. [Google Scholar]
- Coskun, O.; Kanter, M.; Korkmaz, A.; Oter, S. Quercetin, a flavonoid antioxidant, prevents and protects streptozotocininduced oxidative stress and β -cell damage in rat pancreas. Pharmacol. Res. 2005, 51, 117–123. [Google Scholar] [CrossRef]
- Bruce, A.; Freeman, D.; James, C. Biology of disease-free radicals and tissue injury. Lab Investig. 1982, 47, 412–426. [Google Scholar]
- Ahmed, N. Advanced glycation endproducts—Role in pathology of diabetic complications. Diabetes Res. Clin. Pract. 2005, 67, 3–21. [Google Scholar] [CrossRef] [PubMed]
- Yagmur, E.; Tacke, F.; Weiss, C.; Lahme, B.; Manns, M.P.; Kiefer, P.; Trautwein, C.; Gressner, A. Elevation of Nepsilon-(carboxymethyl)lysine-modified advanced glycation end products in chronic liver disease is an indicator of liver cirrhosis. Clin. Biochem. 2006, 39, 39–45. [Google Scholar] [CrossRef]
- Illing, E.K.B.; Gray, C.H.; Lawrence, R.D. Blood glutathione and non-glucose reducing substances in diabetes. Biochem. J. 1951, 48, 637–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andallu, B. Varadacharyulu NCh. Antioxidant role of mulberry leaves in streptozotocin-diabetic rats. Clin. Chim. Acta 2003, 338, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Yamamoto, K.; Tsujihara, N.; Osawa, T. Protective effects of lemon flavonoids on oxidative stress in diabetic rats. Lipids 1998, 33, 689–695. [Google Scholar] [CrossRef]
- Donato, F.; de Gomes, M.G.; Goes, A.T.; Filho, C.B.; Antunes, M.; Souza, L.C.; Boeira, S.; Jesse, C. Hesperidin exerts antidepressant-like effects in acute and chronic treatments in mice: Possible role of l-arginine-NO-cGMP pathway and BDNF levels. Brain Res. Bull. 2014, 104, 19–26. [Google Scholar] [CrossRef]
- Volz, H.C.; Seidel, C.; Laohachewin, D.; Kaya, Z.; Müller, O.J.; Pleger, S.T.; Lasitschka, F.; Bianchi, M.E.; Remppis, A.; Bierhaus, A.; et al. HMGB1: The missing link between diabetes mellitus and heart failure. Basic Res. Cardiol. 2010, 105, 805–820. [Google Scholar] [CrossRef]
- Rahman, A.; Fazal, F. Blocking NF-kappaB: An inflammatory issue. Proc. Am. Thorac. Soc. 2011, 8, 497–503. [Google Scholar] [CrossRef]
- Cipollone, F.; Chiarelli, F.; Iezzi, A.; Fazia, M.; Cuccurullo, C.; Pini, B.; De Cesare, D.; Torello, M.; Tumini, S.; Mezzetti, A. Relationship between Reduced BCL-2 Expression in Circulating Mononuclear Cells and Early Nephropathy in Type 1 Diabetes. Int. J. Immunopathol. Pharmacol. 2005, 18, 625–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cnop, M.; Welsh, N.; Jonas, J.; Jorns, A.; Lenzen, S.; Eizirik, D.L. Mechanisms of pancreatic {beta}-cell death in Type 1 and Type 2 diabetes: Many differences, few similarities. Diabetes 2005, 54, S97–S107. [Google Scholar] [CrossRef] [Green Version]
- Maggini, J.; Mirkin, G.; Geffner, J.R.; Bognanni, I.; Holmberg, J.; Piazzón, I.M.; Nepomnaschy, I.; Costa, H.; Cañones, C.; Raiden, S.; et al. Mouse Bone Marrow-Derived Mesenchymal Stromal Cells Turn Activated Macrophages into a Regulatory-Like Profile. PLoS ONE 2010, 5, e9252. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, H.; Takamura, N.; Shuto, T.; Ogata, K.; Tokunaga, J.; Kawai, K.; Kai, H. The citrus flavonoids hesperetin and naringenin block the lipolytic actions of TNF-α in mouse adipocytes. Biochem. Biophys. Res. Commun. 2010, 394, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Mandrup-Poulsen, T.; Bendtzen, K.; Nielsen, J.H.; Bendixen, G.; Nerup, J. Cytokines Cause Functional and Structural Damage to Isolated Islets of Langerhans. Allergy 1985, 40, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Bendtzen, K.; Barington, T.; Mandrup-Poulsen, T.; Pedersen, J.G.; Svenson, M. Measurement of human IL-1 by LIF induction, pancreatic islet-cell cytotoxicity, and bone resorption. Lymphokine Res. 1986, 5, S93–S98. [Google Scholar] [PubMed]
- Corbett, J.A.; Wang, J.L.; Sweetland, M.A.; Lancaster, J.R.; McDaniel, M.L. Interleukin 1 beta induces the formation of nitric oxide by beta-cells purified from rodent islets of Langerhans. Evidence for the beta-cell as a source and site of action of nitric oxide. J. Clin. Investig. 1992, 90, 2384–2391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Zhang, C.; Graves, D.T. Abnormal Cell Responses and Role of TNF-α in Impaired Diabetic Wound Healing. Biomed. Res. Int. 2013, 2013, 754802. [Google Scholar] [CrossRef] [Green Version]
- Benavente-Garcia, O.; Castillo, J. Update on uses and properties of Citrus flavonolds: New findings in anticancer, cardiovascular, and anti-inflammatory activity. J. Agric. Food Chem. 2008, 56, 6185–6205. [Google Scholar] [CrossRef]
- Sridharan, B.; Mehra, Y.; Ganesh, R.N.; Viswanathan, P. Regulation of urinary crystal inhibiting proteins and inflammatory genes by lemon peel extract and formulated citrus bioflavonoids on ethylene glycol induced urolithic rats. Food Chem. Toxicol. 2006, 94, 75–84. [Google Scholar] [CrossRef]
- Zhao, H.; He, Z.; Huang, D.; Gao, J.; Gong, Y.; Wu, H.; Xu, A.; Meng, X.; Li, Z. Infusion of Bone Marrow Mesenchymal Stem Cells Attenuates Experimental Severe Acute Pancreatitis in Rats. Stem Cells Int. 2016, 2016, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.K.; Huma, Z.E.; Dangles, O. A comprehensive review on flavanones, the major citrus polyphenols. J. Food Compos. Anal. 2014, 33, 85–104. [Google Scholar] [CrossRef]
- Chandravanshi, B.; Bhonde, R.R. Shielding Engineered Islets With Mesenchymal Stem Cells Enhance Survival Under Hypoxia. J. Cell. Biochem. 2017, 118, 2672–2683. [Google Scholar] [CrossRef] [PubMed]
- Yoshimatsu, G.; Sakata, N.; Tsuchiya, H.; Minowa, T.; Takemura, T.; Morita, H.; Hata, T.; Fukase, M.; Aoki, T.; Ishida, M.; et al. The Co-Transplantation of Bone Marrow Derived Mesenchymal Stem Cells Reduced Inflammation in Intramuscular Islet Transplantation. PLoS ONE 2015, 10, e0117561. [Google Scholar] [CrossRef] [Green Version]
- Sabat, R.; Grütz, G.; Warszawska, K.; Kirsch, S.; Witte, E.; Wolk, K.; Geginat, J. Biology of interleukin-10. Cytokine Growth Factor Rev. 2010, 21, 331–344. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liu, J.; Liao, G.; Zhang, J.; Chen, Y.; Li, L.; Li, L.; Liu, F.; Chen, B.; Guo, G.; et al. Early intervention with mesenchymal stem cells prevents nephropathy in diabetic rats by ameliorating the inflammatory microenvironment. Int. J. Mol. Med. 2018, 41, 2629–2639. [Google Scholar] [CrossRef] [Green Version]
- Comalada, M.; Ballester, I.; Bailón, E.; Sierra, S.; Xaus, J.; Gálvez, J.; de Medina, F.S.; Zarzuelo, A. Inhibition of pro-inflammatory markers in primary bone marrow-derived mouse macrophages by naturally occurring flavonoids: Analysis of the structure–activity relationship. Biochem. Pharmacol. 2006, 72, 1010–1021. [Google Scholar] [CrossRef]
- Crouvezier, S.; Powell, B.; Keir, D.; Yaqoob, P. The effects of phenolic components of tea on the production of pro- and anti-inflammatory cytokines by human leukocytes in vitro. Cytokine 2001, 13, 280–286. [Google Scholar] [CrossRef]
- Abdi, R.; Fiorina, P.; Adra, C.N.; Atkinson, M.; Sayegh, M.H. Immunomodulation by mesenchymal stem cells: A potential therapeutic strategy for type 1 diabetes. Diabetes 2008, 57, 1759–1767. [Google Scholar] [CrossRef] [Green Version]
- Alabsi, A.M.; Ali, R.; Ali, A.M.; Harun, H.; Al-Dubai, S.A.R.; Ganasegeran, K.; Alshagga, M.A.; Salem, S.D.; Abu Kasim, N.H.B. Induction of Caspase-9, Biochemical Assessment and Morphological Changes Caused by Apoptosis in Cancer Cells Treated with Goniothalamin Extracted from Goniothalamus macrophyllus. Asian Pac. J. Cancer Prev. 2013, 14, 6273–6280. [Google Scholar] [CrossRef] [Green Version]
- Anzola, M.; Burgos, J.J. Hepatocellular carcinoma: Molecular interactions between hepatitis C virus and p53 in hepatocarcinogenesis. Expert Rev. Mol. Med. 2003, 5, 926. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.T.; Holloway, M.P.; A Altura, R. The CRM1 nuclear export protein in normal development and disease. Int. J. Biochem. Mol. Biol. 2012, 3, 137–151. [Google Scholar] [PubMed]
- Tse, C.; Shoemaker, A.R.; Adickes, J.; Anderson, M.G.; Chen, J.; Jin, S.; Johnson, E.F.; Marsh, K.C.; Mitten, M.J.; Nimmer, P.; et al. ABT-263: A Potent and Orally Bioavailable Bcl-2 Family Inhibitor. Cancer Res. 2008, 68, 3421–3428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Kholy, W.M.; Hussein, R.H.; Khalil, D.Y. Antioxidant and Anti-Inflammatory Therapeutic Roles of BM-Mscs in Enhancing Pancreatic Auto-Immunity and Apoptotic Status in TIDM. J. Am. Sci. 2018, 14, 12. [Google Scholar]
- Haidara, M.A.; Assiri, A.S.; Youssef, A.; Mahmoud, M.M.; Eajaz, A.M.S.; Al-Hakami, A.; Chandramoorthy, H.C. Differentiated mesenchymal stem cells ameliorate cardiovascular complications in diabetic rats. Cell Tissue Res. 2014, 358, 325. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Oksbjerg, N.; Young, J.; Jeppesen, P.B. Caffeic acid, naringenin and quercetin enhance glucose-stimulated insulin secretion and glucose sensitivity in INS-1E cells. Diabetes Obes. Metab. 2014, 16, 602–612. [Google Scholar] [CrossRef]
- Lee, D.; Kim, K.H.; Lee, J.; Hwang, G.S.; Lee, H.L.; Hahm, D.-H.; Huh, C.K.; Lee, S.C.; Lee, S.; Kang, K.S. Protective effect of cirsimaritin against streptozotocin-induced apoptosis in pancreatic beta cells. J. Pharm. Pharmacol. 2017, 69, 875–883. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, D. Flavonol kaempferol improves chronic hyperglycemia-impaired pancreatic beta-cell viability and insulin secretory function. Eur. J. Pharmacol. 2011, 670, 325–332. [Google Scholar] [CrossRef]
- Carrasco-Pozo, C.; Tan, K.N.; Reyes-Farias, M.; de la Jara, N.; Ngo, S.T.; Garcia-Diaz, D.F.; Llanos, P.; Cires, M.J.; Borges, K. The deleterious effect of cholesterol and protection by quercetin on mitochondrial bioenergetics of pancreatic β-cells, glycemic control and inflammation: In vitro and in vivo studies. Redox Biol. 2016, 9, 229–243. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Jiang, H.; Yang, H. In vitro culture of bone marrow mesenchymal stem cells in rats and differentiation into retinal neural-like cells. J. Huazhong Univ. Sci. Technol. 2007, 27, 598–600. [Google Scholar] [CrossRef]
- Chaudhary, J.K.; Rath, P.C. A Simple Method for Isolation, Propagation, Characterization, and Differentiation of Adult Mouse Bone Marrow-Derived Multipotent Mesenchymal Stem Cells. J. Cell Sci. Ther. 2016, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Dobbin, Z.C.; Landen, C.N. Isolation and Characterization of Potential Cancer Stem Cells from Solid Human Tumors—Potential Applications. Curr. Protoc. Pharmacol. 2013, 63, 8s63. [Google Scholar] [CrossRef] [Green Version]
- Tariq, M.; Masoud, M.S.; Mehmood, A.; Khan, S.N.; Riazuddin, S. Stromal cell derived factor-1alpha protects stem cell derived insulin-producing cells from glucotoxicity under high glucose conditions in-vitro and ameliorates drug induced diabetes in rats. J. Transl. Med. 2013, 11, 115–210. [Google Scholar] [CrossRef] [Green Version]
- Revathy, J.; Sheik Abdullah, S. Effect of hesperetin on hyperglycemia-mediated oxidative damage in the hepatic and renal tissues of experimental diabetic rats. J. Glob. Biosci. 2016, 5, 3980–3985. [Google Scholar]
- Baker, J.R.; Metcalf, P.A.; Holdway, I.; Johnson, R.N. Serum fructosamine concentration as a measure of blood glucose control in type 1 (insulin dependent) diabetes. Br. Med. J. 1985, 290, 352–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seifter, S.; Dayton, S.; Novic, B.; Muntwyler, E. The estimation of glycogen with anthrone reagent. Arch. Biochem. 1950, 25, 191–200. [Google Scholar] [PubMed]
- Begum, N.; Sam, G.P.; Shanmugasundaram, K.R. Serum enzymes in human and experimental diabetes mellitus. Indian J. Med. Res. 1978, 68, 774–784. [Google Scholar] [PubMed]
- Stallman, W.; Hers, H.G. The stimulation of liver phosphorylase b by AMP, fluoride and sulfate. A technical note of the specific determination of the a and b forms of liver glycogen phosphorylase. Eur. J. Biochem. 1975, 54, 341–350. [Google Scholar] [CrossRef] [PubMed]
- EL-Merzabani, M.M.; EL-Aaser, A.A.; Zakhary, N.I. Determination of inorganic phosphorus in serum. J. Clin. Chem. Clin. Biochem. 1977, 15, 715–718. [Google Scholar]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Beutler, E.; Duron, O.; Kelly, B.M. Improved method for the determination of blood glutathione. J. Lab. Clin. Med. 1963, 61, 882–888. [Google Scholar] [PubMed]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar] [PubMed]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, D.M.; Spooner, R.J. Methods of Enzymatic Analysis, 3rd ed.; Bergmeyen, H.V., Ed.; Verlag Chemie: Weinheim, Germany, 1983; Volume 3, pp. 258–265. [Google Scholar]
- Bancroft, J.D.; Stevens, A. Theory and Practice of Histological Techniques, 2nd ed.; Churchill-Livingstone: New York, NY, USA, 1982; pp. 374–375. [Google Scholar]
- Cemek, M.; Kaga, S.; Simsek, N.; Buyukokuroglu, M.E.; Konuk, M. Antihyperglycemic and antioxidative potential of Matricaria chamomilla L. in streptozotocin-induced diabetic rats. J. Nat. Med. 2008, 62, 284–293. [Google Scholar] [CrossRef]
- Duncan, B.D. Multiple ranges tests for correlated and heteroscedastic means. Biometrics 1957, 13, 359–364. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Glucose Level 10 Days Post-STZ Injection (Pre-Treatment) | Glucose Level at the End of the Experiment (Post-Treatment) | ||
|---|---|---|---|---|
| Fasting (mg/dL) | 2 h (mg/dL) | Fasting (mg/dL) | 2 h (mg/dL) | |
| Diabetic control | 208.91 ± 21.74 | 459.17 ± 13.16 | 361.83 ± 36.97 # | 490.67 ± 67.55 |
| Diabetic + hesperetin | 212.60 ± 15.23 | 489.605 ± 66.05 | 187.63 ± 29.30 * | 189.50 ± 13.52 *,# |
| Diabetic + BM-MSCs | 225.85 ± 16.41 | 457.20 ± 74.04 | 225.38 ± 31.63 *,$ | 226.00 ± 52.38 *,# |
| Diabetic + hesperetin and BM-MSCs | 221.60 ± 18.99 | 473.33 ± 78.91 | 129.3 ± 21.07 * | 133.83 ± 9.45 *,# |
| Groups | Fructosamine (mmole/L) | Insulin (ng/mL) | C-Peptide (ng/mL) |
|---|---|---|---|
| Normal | 0.462 ± 0.037 | 2.50 ± 0.15 | 6.64 ± 0.27 |
| Diabetic control | 1.167 ± 0.062 † | 1.21 ± 0.07 † | 2.72 ± 0.21 † |
| Diabetic + hesperetin | 0.562 ± 0.032 * | 2.25 ± 0.09 * | 5.08 ± 0.27 * |
| Diabetic + BM-MSCs | 0.547 ± 0.014 * | 2.13 ± 0.09 * | 4.75 ± 0.02 *,$ |
| Diabetic + hesperetin and BM-MSCs | 0.565 ± 0.029 * | 2.01 ± 0.03 * | 5.46 ± 0.15 * |
| Groups | Liver Glycogen (mg Glycogen/g Liver) | Liver G6Pase (mg Pi Liberated/g Tissue/h) | Liver GP (mg Piliberated/g Tissue/h) |
|---|---|---|---|
| Normal | 11.44 ± 1.44 | 0.37 ± 0.01 | 0.36 ± 0.02 |
| Diabetic control | 0.50 ± 0.07 † | 0.76 ± 0.06 † | 0.97 ± 0.04 † |
| Diabetic + hesperetin | 2.10 ± 0.33 *,$ | 0.43 ± 0.01 * | 0.57 ± 0.06 *,$ |
| Diabetic + BM-MSCs | 9.00 ± 0.64 *,$ | 0.43 ± 0.01 * | 0.67 ± 0.07 * |
| Diabetic + hesperetin and BM-MSCs | 4.25 ± 0.29 * | 0.46 ± 0.01 * | 0.76 ± 0.05 * |
| Groups | LPO (nmole MDA/100 mg Tissue) | GSH (nmole/100 mg Tissue) | GPx (U/g Tissue) |
|---|---|---|---|
| Normal | 26.73 ± 0.59 | 57.27 ± 1.14 | 48.20 ± 4.40 |
| Diabetic control | 142.77 ± 14.65 † | 24.83 ± 1.01 † | 17.72 ± 0.96 † |
| Diabetic + hesperetin | 90.42 ± 3.92 * | 34.77 ± 1.79 * | 20.09 ± 0.71 $ |
| Diabetic + BM-MSCs | 64.78 ± 4.44 *,$ | 50.20 ± 0.99 *,$ | 28.95 ± 1.21 * |
| Diabetic + hesperetin and BM-MSCs | 76.95 ± 6.95 * | 34.37 ± 1.96 * | 26.46 ± 0.75 * |
| Groups | GST (U/100 mg Tissue) | GR (mU/100 mg Tissue) |
|---|---|---|
| Normal | 116.83 ± 3.60 | 14 ± 0.73 |
| Diabetic control | 62.20 ± 1.93 † | 5.5 ± 0.55 † |
| Diabetic + hesperetin | 92.43 ± 4.77 *,$ | 20.17 ± 0.83 * |
| Diabetic + BM-MSCs | 82.88 ± 2.99 *,$ | 19.67 ± 1.76 * |
| Diabetic + hesperetin and BM-MSCs | 69.42 ± 2.66 † | 22.17 ± 0.95 * |
| Groups | NF-κB (Fold Change, Relative to Normal) | IL-1β (Fold Change, Relative to Normal) | IL-10 (Fold Change, Relative to Normal) |
|---|---|---|---|
| Normal | 1.01 ± 0.002 | 1.01 ± 0.002 | 1.01 ± 0.004 |
| Diabetic control | 5.00 ± 0.1 1 † | 6.65 ± 0.09 † | 0.25 ± 0.03 † |
| Diabetic + hesperetin | 1.63 ± 0.16 *,$ | 2.45 ± 0.05 *,$ | 0.65 ± 0.03 *,$ |
| Diabetic + BM-MSCs | 1.45 ± 0.05 *,$ | 2.24 ± 0.06 * | 0.76 ± 0.01 * |
| Diabetic + hesperetin and BM-MSCs | 2.05 ± 0.01 * | 2.10 ± 0.22 * | 0.72 ± 0.01* |
| Groups | TNF-α (pg/mL) | IL-17 (pg/mL) | IL-4 (pg/mL) |
|---|---|---|---|
| Normal | 14.73 ± 0.47 | 42.57 ± 1.42 | 120.50 ± 2.80 |
| Diabetic control | 106.12 ± 3.50 † | 122.35 ± 2.10 † | 40.03 ± 3.66 † |
| Diabetic + hesperetin | 49.27 ± 2.56 *,$ | 68.13 ± 1.86 *,$ | 90.33 ± 3.65 * |
| Diabetic + BM-MSCs | 43.40 ± 2.35 * | 66.00 ± 6.68 *,$ | 95.10 ± 4.83 * |
| Diabetic + hesperetin and BM-MSCs | 39.33 ± 5.12 * | 52.20 ± 4.11 * | 96.77 ± 5.93 * |
| Groups | P53 (Fold Change, Relative to Normal Control Rats) | Bcl-2 (Fold Change, Relative to Normal Control Rats) |
|---|---|---|
| Normal | 1.02 ± 0.01 | 1.01 ± 0.002 |
| Diabetic control | 6.77 ± 0.1 † | 0.23 ± 0.004 † |
| Diabetic + hesperetin | 3.21 ± 0.07 *,$ | 0.56 ± 0.03 *,$ |
| Diabetic + BM-MSCs | 2.95 ± 0.09 * | 0.57 ± 0.01 *,$ |
| Diabetic + hesperetin and BM-MSCs | 2.65 ± 0.31 * | 0.65 ± 0.01 * |
| Histopathological Lesion Score | ||
|---|---|---|
| Groups | Vacuolations | Necrosis |
| Normal | − | − |
| Diabetic control | +++ | +++ |
| Diabetic + hesperetin | + | − |
| Diabetic + BM-MSCs | ++ | + |
| Diabetic + hesperetin and BM-MSCs | − | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, O.M.; Saleh, A.S.; Ahmed, E.A.; Ghoneim, M.M.; Ebrahim, H.A.; Abdelgawad, M.A.; Abdel-Gabbar, M. Efficiency of Bone Marrow-Derived Mesenchymal Stem Cells and Hesperetin in the Treatment of Streptozotocin-Induced Type 1 Diabetes in Wistar Rats. Pharmaceuticals 2023, 16, 859. https://doi.org/10.3390/ph16060859
Ahmed OM, Saleh AS, Ahmed EA, Ghoneim MM, Ebrahim HA, Abdelgawad MA, Abdel-Gabbar M. Efficiency of Bone Marrow-Derived Mesenchymal Stem Cells and Hesperetin in the Treatment of Streptozotocin-Induced Type 1 Diabetes in Wistar Rats. Pharmaceuticals. 2023; 16(6):859. https://doi.org/10.3390/ph16060859
Chicago/Turabian StyleAhmed, Osama M., Ablaa S. Saleh, Eman A. Ahmed, Mohammed M. Ghoneim, Hasnaa Ali Ebrahim, Mohamed A. Abdelgawad, and Mohammed Abdel-Gabbar. 2023. "Efficiency of Bone Marrow-Derived Mesenchymal Stem Cells and Hesperetin in the Treatment of Streptozotocin-Induced Type 1 Diabetes in Wistar Rats" Pharmaceuticals 16, no. 6: 859. https://doi.org/10.3390/ph16060859