
Local Delivery Strategies for Peptides and Proteins into the CNS: Status Quo, Challenges, and Future Perspectives

Abstract
:1. Introduction
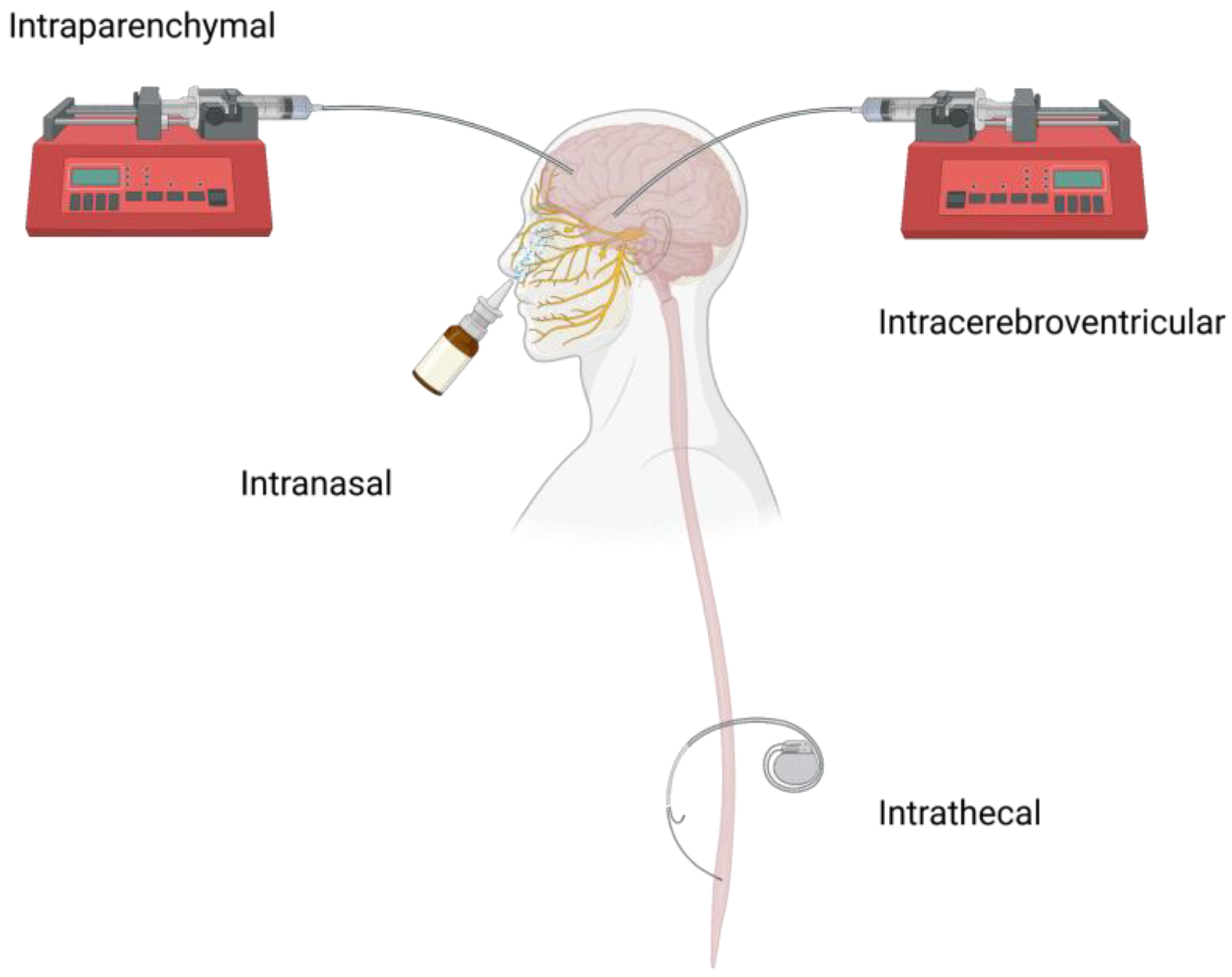
2. Local CNS Administration Routes and Challenges
2.1. ICV
2.2. CED
2.3. IT
2.4. Others
3. Formulation Strategies to Improve CNS Protein/Peptide Delivery
3.1. Peptide/Protein Functionalization Strategies
3.2. Long-Acting Formulations
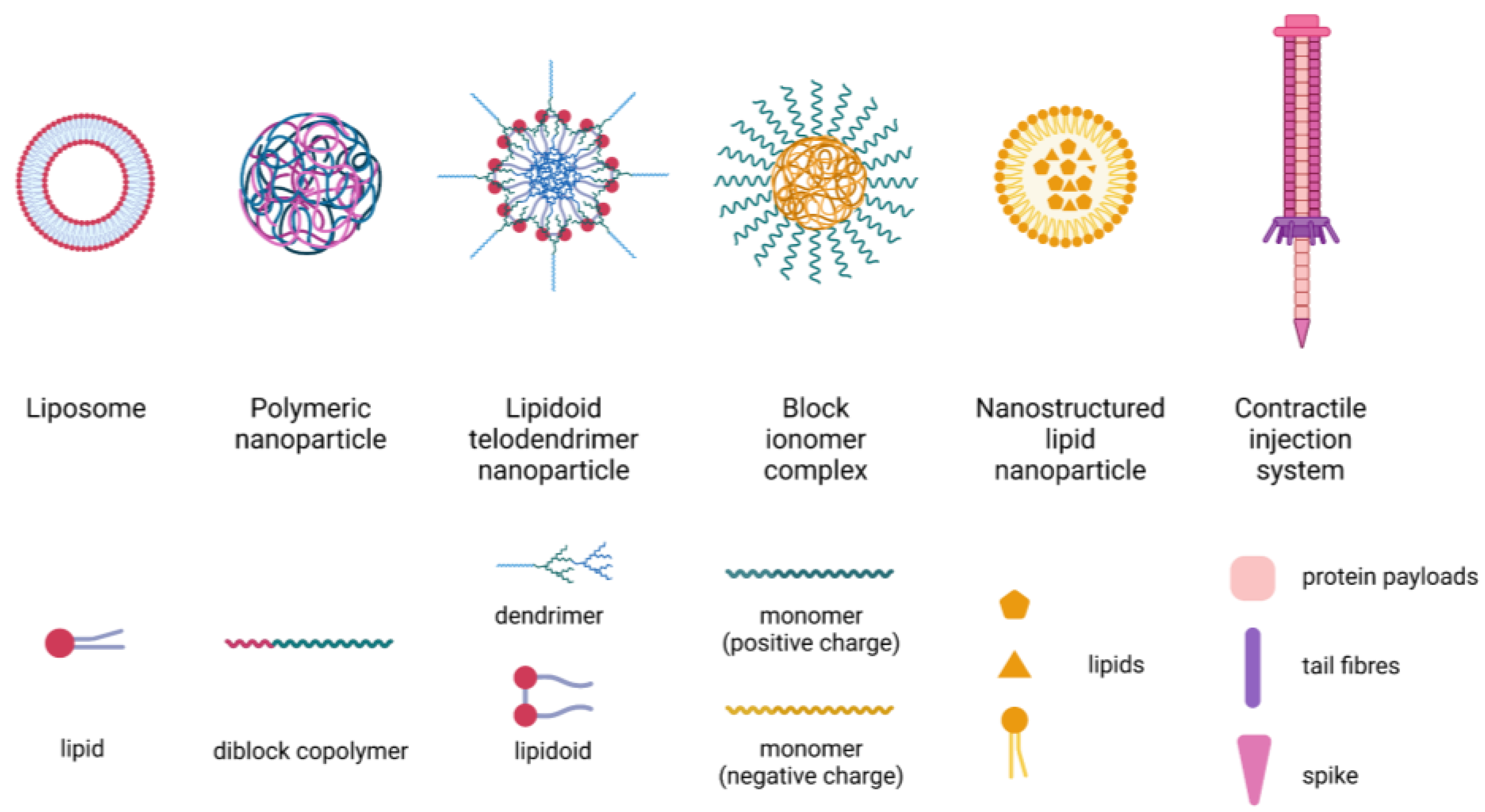
3.3. Nanotechnology-Based Delivery Systems
3.4. Extracellular Vesicles
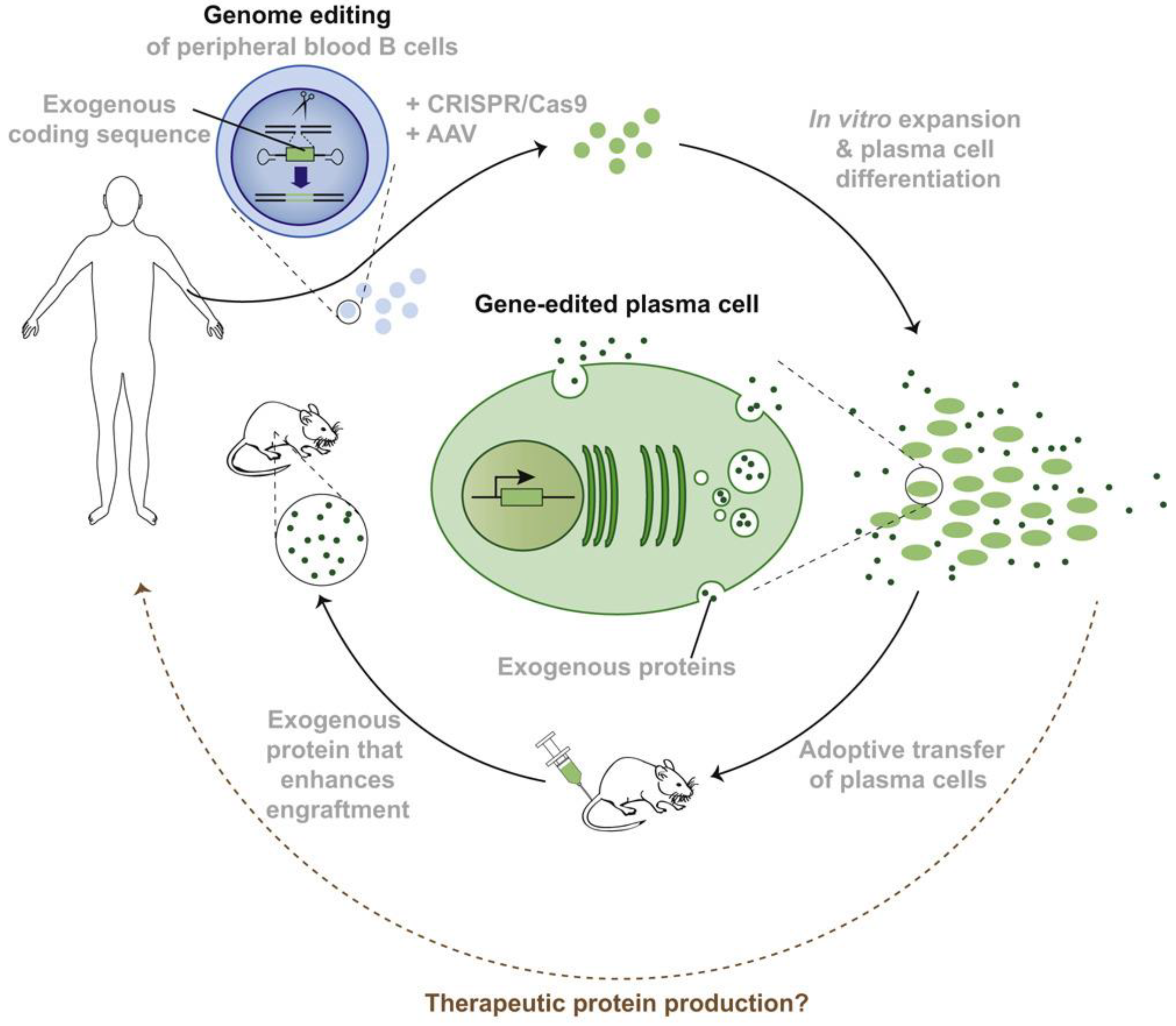
3.5. Live-Cell Therapy
4. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hartley, H. Origin of the Word ‘Protein’. Nature 1951, 168, 244. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.; Knight, G.; Wiles, P.; Keen, H.; Ward, J.; Cauldwell, J.; Adeniyi-Jones, R.; Leiper, J.; Jones, R.; Maccuish, A.; et al. Biosynthetic Human Insulin in the Treatment of Diabetes: A Double-blind Crossover Trial in Established Diabetic Patients. Lancet 1982, 320, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Brasnjevic, I.; Steinbusch, H.W.; Schmitz, C.; Martinez-Martinez, P. Delivery of peptide and protein drugs over the blood–brain barrier. Prog. Neurobiol. 2009, 87, 212–251. [Google Scholar] [CrossRef] [PubMed]
- Hökfelt, T.; Bartfai, T.; Bloom, F. Neuropeptides: Opportunities for drug discovery. Lancet Neurol. 2003, 2, 463–472. [Google Scholar] [CrossRef]
- Oosthuyse, B.; Moons, L.; Storkebaum, E.; Beck, H.; Nuyens, D.; Brusselmans, K.; Van Dorpe, J.; Hellings, P.; Gorselink, M.; Heymans, S.; et al. Deletion of the hypoxia-response element in the vascular endothelial growth factor promoter causes motor neuron degeneration. Nat. Genet. 2001, 28, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Craft, S.; Claxton, A.; Baker, L.D.; Hanson, A.J.; Cholerton, B.; Trittschuh, E.H.; Dahl, D.; Caulder, E.; Neth, B.; Montine, T.J.; et al. Effects of Regular and Long-Acting Insulin on Cognition and Alzheimer’s Disease Biomarkers: A Pilot Clinical Trial. J. Alzheimers Dis. 2017, 57, 1325–1334. [Google Scholar] [CrossRef]
- Sayed, S.; Van Dam, N.; Horn, S.R.; Kautz, M.M.; Parides, M.; Costi, S.; Collins, K.A.; Iacoviello, B.; Iosifescu, D.V.; Mathé, A.A.; et al. A Randomized Dose-Ranging Study of Neuropeptide Y in Patients with Posttraumatic Stress Disorder. Int. J. Neuropsychopharmacol. 2018, 21, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Proulx, S.T. Cerebrospinal fluid outflow: A review of the historical and contemporary evidence for arachnoid villi, perineural routes, and dural lymphatics. Cell. Mol. Life Sci. 2021, 78, 2429–2457. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M. The blood-brain barrier: Bottleneck in brain drug development. Neurorx 2005, 2, 3–14. [Google Scholar] [CrossRef]
- Lin, J.H. Pharmacokinetics of Biotech Drugs: Peptides, Proteins and Monoclonal Antibodies. Curr. Drug Metab. 2009, 10, 661–691. [Google Scholar] [CrossRef]
- Bumbaca, B.; Li, Z.; Shah, D.K. Pharmacokinetics of protein and peptide conjugates. Drug Metab. Pharmacokinet. 2019, 34, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Filippi, C.G.; Wong, T.; Ray, A.; Fralin, S.; Tsiouris, A.J.; Praminick, B.; Demopoulos, A.; McCrea, H.J.; Bodhinayake, I.; et al. Superselective intraarterial cerebral infusion of cetuximab after osmotic blood/brain barrier disruption for recurrent malignant glioma: Phase I study. J. Neuro-Oncol. 2016, 128, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Chaichana, K.L.; Pinheiro, L.; Brem, H. Delivery of local therapeutics to the brain: Working toward advancing treatment for malignant gliomas. Ther. Deliv. 2015, 6, 353–369. [Google Scholar] [CrossRef] [PubMed]
- Wolinsky, J.B.; Colson, Y.L.; Grinstaff, M.W. Local drug delivery strategies for cancer treatment: Gels, nanoparticles, polymeric films, rods, and wafers. J. Control. Release 2012, 159, 14–26. [Google Scholar] [CrossRef]
- Yi, X.; Manickam, D.S.; Brynskikh, A.; Kabanov, A.V. Agile delivery of protein therapeutics to CNS. J. Control. Release 2014, 190, 637–663. [Google Scholar] [CrossRef] [PubMed]
- Press Releases from BioMarin. Available online: https://investors.biomarin.com/2017-04-27-FDA-Approves-Brineura-TM-cerliponase-alfa-for-the-Treatment-of-CLN2-Disease-a-Form-of-Batten-Disease-and-Ultra-Rare-Pediatric-Brain-Disorder-in-Children (accessed on 26 April 2023).
- Schulz, A.; Ajayi, T.; Specchio, N.; Reyes, E.D.L.; Gissen, P.; Ballon, D.; Dyke, J.P.; Cahan, H.; Slasor, P.; Jacoby, D.; et al. Study of Intraventricular Cerliponase Alfa for CLN2 Disease. N. Engl. J. Med. 2018, 378, 1898–1907. [Google Scholar] [CrossRef]
- Gissen, P.; Specchio, N.; Olaye, A.; Jain, M.; Butt, T.; Ghosh, W.; Ruban-Fell, B.; Griffiths, A.; Camp, C.; Sisic, Z.; et al. Investigating health-related quality of life in rare diseases: A case study in utility value determination for patients with CLN2 disease (neuronal ceroid lipofuscinosis type 2). Orphanet J. Rare Dis. 2021, 16, 217. [Google Scholar] [CrossRef]
- Seo, J.-H.; Kosuga, M.; Hamazaki, T.; Shintaku, H.; Okuyama, T. Impact of intracerebroventricular enzyme replacement therapy in patients with neuronopathic mucopolysaccharidosis type II. Mol. Ther.-Methods Clin. Dev. 2021, 21, 67–75. [Google Scholar] [CrossRef]
- Van Damme, P.; Tilkin, P.; Mercer, K.J.; Terryn, J.; D’hondt, A.; Herne, N.; Tousseyn, T.; Claeys, K.G.; Thal, D.R.; Zachrisson, O.; et al. Intracerebroventricular delivery of vascular endothelial growth factor in patients with amyotrophic lateral sclerosis, a phase I study. Brain Commun. 2020, 2, fcaa160. [Google Scholar] [CrossRef]
- Bander, E.D.; Ramos, A.D.; Wembacher-Schroeder, E.; Ivasyk, I.; Thomson, R.; Morgenstern, P.F.; Souweidane, M.M. Repeat convection-enhanced delivery for diffuse intrinsic pontine glioma. J. Neurosurg. Pediatr. 2020, 26, 661–666. [Google Scholar] [CrossRef]
- Souweidane, M.M.; Kramer, K.; Pandit-Taskar, N.; Zhou, Z.; Haque, S.; Zanzonico, P.; Carrasquillo, J.A.; Lyashchenko, S.K.; Thakur, S.B.; Donzelli, M.; et al. Convection-enhanced delivery for diffuse intrinsic pontine glioma: A single-centre, dose-escalation, phase 1 trial. Lancet Oncol. 2018, 19, 1040–1050. [Google Scholar] [CrossRef] [PubMed]
- Dali, C.; Sevin, C.; Krägeloh-Mann, I.; Giugliani, R.; Sakai, N.; Wu, J.; Wasilewski, M. Safety of intrathecal delivery of recombinant human arylsulfatase A in children with metachromatic leukodystrophy: Results from a phase 1/2 clinical trial. Mol. Genet. Metab. 2020, 131, 235–244. [Google Scholar] [CrossRef]
- Luger, T.J.; Kathrein, A.; Rieger, M.; Lorenz, I.H. Intracerebroventricular and intrathecal injectate spread in rats. Eur. J. Anaesthesiol. 2005, 22, 236–239. [Google Scholar] [CrossRef]
- Schultz, M.L.; Tecedor, L.; Chang, M.; Davidson, B.L. Clarifying lysosomal storage diseases. Trends Neurosci. 2011, 34, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Bellettato, C.M.; Scarpa, M. Pathophysiology of neuropathic lysosomal storage disorders. J. Inherit. Metab. Dis. 2010, 33, 347–362. [Google Scholar] [CrossRef]
- Brown, R.H.; Al-Chalabi, A. Amyotrophic Lateral Sclerosis. N. Engl. J. Med. 2017, 377, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.E.; Boumaza, I.; Lacomis, D.; Bowser, R. Cystatin C: A Candidate Biomarker for Amyotrophic Lateral Sclerosis. PLoS ONE 2010, 5, e15133. [Google Scholar] [CrossRef]
- Okamoto, K.; Hirai, S.; Amari, M.; Watanabe, M.; Sakurai, A. Bunina bodies in amyotrophic lateral sclerosis immunostained with rabbit anti-cystatin C serum. Neurosci. Lett. 1993, 162, 125–128. [Google Scholar] [CrossRef]
- Watanabe, S.; Komine, O.; Endo, F.; Wakasugi, K.; Yamanaka, K. Intracerebroventricular administration of Cystatin C ameliorates disease in SOD1-linked amyotrophic lateral sclerosis mice. J. Neurochem. 2018, 145, 80–89. [Google Scholar] [CrossRef]
- Iliff, J.J.; Wang, M.; Liao, Y.; Plogg, B.A.; Peng, W.; Gundersen, G.A.; Benveniste, H.; Vates, G.E.; Deane, R.; Goldman, S.A.; et al. A Paravascular Pathway Facilitates CSF Flow Through the Brain Parenchyma and the Clearance of Interstitial Solutes, Including Amyloid β. Sci. Transl. Med. 2012, 4, 147ra111. [Google Scholar] [CrossRef]
- Noguchi, Y.; Kato, M.; Ozeki, K.; Ishigai, M. Pharmacokinetics of an intracerebroventricularly administered antibody in rats. mAbs 2017, 9, 1210–1215. [Google Scholar] [CrossRef] [PubMed]
- Dedrick, R.L.; Flessner, M.F. Pharmacokinetic considerations on monoclonal antibodies. Prog. Clin. Biol. Res. 1989, 288, 429–438. [Google Scholar] [PubMed]
- Lonser, R.R.; Sarntinoranont, M.; Morrison, P.F.; Oldfield, E.H. Convection-enhanced delivery to the central nervous system. J. Neurosurg. 2015, 122, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Bobo, R.H.; Laske, D.W.; Akbasak, A.; Morrison, P.F.; Dedrick, R.L.; Oldfield, E.H. Convection-enhanced delivery of macromolecules in the brain. Proc. Natl. Acad. Sci. USA 1994, 91, 2076–2080. [Google Scholar] [CrossRef]
- Weber, F.W.; Floeth, F.; Asher, A.; Bucholz, R.; Berge, M.; Pradoss, M.; Chang, S.; Bruces, J.; Hall, W.; Raino, N.G.; et al. Local Convection Enhanced Delivery of IL4-Pseudomonas Exotoxin (NBI-3001) for Treatment of Patients with Recurrent Malignant Glioma; Springer: Vienna, Austria, 2003; pp. 93–103. [Google Scholar] [CrossRef]
- Kunwar, S.; Chang, S.; Westphal, M.; Vogelbaum, M.; Sampson, J.; Barnett, G.; Shaffrey, M.; Ram, Z.; Piepmeier, J.; Prados, M.; et al. Phase III randomized trial of CED of IL13-PE38QQR vs. Gliadel wafers for recurrent glioblastoma. Neuro-Oncology 2010, 12, 871–881. [Google Scholar] [CrossRef]
- Salvatore, M.F.; Ai, Y.; Fischer, B.; Zhang, A.M.; Grondin, R.C.; Zhang, Z.; Gerhardt, G.A.; Gash, D.M. Point source concentration of GDNF may explain failure of phase II clinical trial. Exp. Neurol. 2006, 202, 497–505. [Google Scholar] [CrossRef]
- Pardridge, W.M. Biopharmaceutical drug targeting to the brain. J. Drug Target. 2010, 18, 157–167. [Google Scholar] [CrossRef]
- Sampson, J.H.; Akabani, G.; Archer, G.E.; Berger, M.S.; Coleman, R.E.; Friedman, A.H.; Friedman, H.S.; Greer, K.; Herndon, J.E.; Kunwar, S.; et al. Intracerebral infusion of an EGFR-targeted toxin in recurrent malignant brain tumors. Neuro-Oncology 2008, 10, 320–329. [Google Scholar] [CrossRef]
- Looseley, A. Corning and Cocaine: The Advent of Spinal Anaesthesia; Grand Rounds: Beckenham, UK, 2009; Volume 9. [Google Scholar] [CrossRef]
- Cudkowicz, M.E.; Warren, L.; Francis, J.W.; Lloyd, K.J.; Friedlander, R.M.; Borges, L.F.; Kassem, N.; Munsta, T.L.; Brown, R.H. Intrathecal administration of recombinant human superoxide dismutase 1 in amyotrophic lateral sclerosis: A preliminary safety and pharmacokinetic study. Neurology 1997, 49, 213–222. [Google Scholar] [CrossRef]
- Pizzo, M.E.; Wolak, D.J.; Kumar, N.N.; Brunette, E.; Brunnquell, C.L.; Hannocks, M.; Abbott, N.J.; Meyerand, M.E.; Sorokin, L.; Stanimirovic, D.B.; et al. Intrathecal antibody distribution in the rat brain: Surface diffusion, perivascular transport and osmotic enhancement of delivery. J. Physiol. 2018, 596, 445–475. [Google Scholar] [CrossRef]
- Dhuria, S.V.; Hanson, L.R.; Frey, W.H., 2nd. Intranasal delivery to the central nervous system: Mechanisms and experimental considerations. J. Pharm. Sci. 2010, 99, 1654–1673. [Google Scholar] [CrossRef] [PubMed]
- Falcone, J.A.; Salameh, T.S.; Yi, X.; Cordy, B.J.; Mortell, W.G.; Kabanov, A.V.; Banks, W.A. Intranasal Administration as a Route for Drug Delivery to the Brain: Evidence for a Unique Pathway for Albumin. Experiment 2014, 351, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Craft, S.; Baker, L.D.; Montine, T.J.; Minoshima, S.; Watson, G.S.; Claxton, A.; Arbuckle, M.; Callaghan, M.; Tsai, E.; Plymate, S.R.; et al. Intranasal insulin therapy for Alzheimer disease and amnestic mild cognitive impairment: A pilot clinical trial. Arch. Neurol. 2012, 69, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Benedict, C.; Hallschmid, M.; Schmitz, K.; Schultes, B.; Ratter, F.; Fehm, H.L.; Born, J.; Kern, W. Intranasal Insulin Improves Memory in Humans: Superiority of Insulin Aspart. Neuropsychopharmacology 2007, 32, 239–243. [Google Scholar] [CrossRef]
- Thorne, R.; Pronk, G.; Padmanabhan, V.; Frey, W. Delivery of insulin-like growth factor-I to the rat brain and spinal cord along olfactory and trigeminal pathways following intranasal administration. Neuroscience 2004, 127, 481–496. [Google Scholar] [CrossRef]
- Banks, W.A.; During, M.J.; Niehoff, M.L. Brain Uptake of the Glucagon-Like Peptide-1 Antagonist Exendin(9-39) after Intranasal Administration. Experiment 2004, 309, 469–475. [Google Scholar] [CrossRef]
- Nonaka, N.; Farr, S.A.; Kageyama, H.; Shioda, S.; Banks, W.A. Delivery of Galanin-Like Peptide to the Brain: Targeting with Intranasal Delivery and Cyclodextrins. Experiment 2008, 325, 513–519. [Google Scholar] [CrossRef]
- Ross, T.; Martinez, P.; Renner, J.; Thorne, R.; Hanson, L.; Frey, W. Intranasal administration of interferon beta bypasses the blood–brain barrier to target the central nervous system and cervical lymph nodes: A non-invasive treatment strategy for multiple sclerosis. J. Neuroimmunol. 2004, 151, 66–77. [Google Scholar] [CrossRef]
- Dhuria, S.V.; Hanson, L.R.; Frey, W.H. Intranasal drug targeting of hypocretin-1 (orexin-A) to the central nervous system. J. Pharm. Sci. 2004, 98, 2501–2515. [Google Scholar] [CrossRef]
- Hellard, E.A.R.; Impastato, R.A.; Gilpin, N.W. Intra-cerebral and intra-nasal melanocortin-4 receptor antagonist blocks withdrawal hyperalgesia in alcohol-dependent rats. Addict. Biol. 2017, 22, 692–701. [Google Scholar] [CrossRef]
- Craft, S.; Raman, R.; Chow, T.W.; Rafii, M.S.; Sun, C.-K.; Rissman, R.A.; Donohue, M.C.; Brewer, J.B.; Jenkins, C.; Harless, K.; et al. Safety, Efficacy, and Feasibility of Intranasal Insulin for the Treatment of Mild Cognitive Impairment and Alzheimer Disease Dementia. JAMA Neurol. 2020, 77, 1099–1109. [Google Scholar] [CrossRef]
- Kang, C.E.; Tator, C.H.; Shoichet, M.S. Poly(ethylene glycol) modification enhances penetration of fibroblast growth factor 2 to injured spinal cord tissue from an intrathecal delivery system. J. Control. Release 2010, 144, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Pizzo, D.; Thal, L. Intraparenchymal nerve growth factor improves behavioral deficits while minimizing the adverse effects of intracerebroventricular delivery. Neuroscience 2004, 124, 743–755. [Google Scholar] [CrossRef]
- Izadpanah, M.; Dargahi, L.; Ai, J.; Taei, A.A.; Barough, S.E.; Mowla, S.J.; TavoosiDana, G.; Farahmandfar, M. Extracellular Vesicles as a Neprilysin Delivery System Memory Improvement in Alzheimer’s Disease. Iran. J. Pharm. Res. 2020, 19, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Kauer, T.M.; Figueiredo, J.-L.; Hingtgen, S.; Shah, K. Encapsulated therapeutic stem cells implanted in the tumor resection cavity induce cell death in gliomas. Nat. Neurosci. 2012, 15, 197–204. [Google Scholar] [CrossRef]
- Laske, D.W.; Youle, R.J.; Oldfield, E.H. Tumor regression with regional distribution of the targeted toxin TF-CRM107 in patients with malignant brain tumors. Nat. Med. 1997, 3, 1362–1368. [Google Scholar] [CrossRef] [PubMed]
- Frankel, A.; Liu, J.-S.; Rizzieri, D.; Hogge, D. Phase I clinical study of diphtheria toxin-interleukin 3 fusion protein in patients with acute myeloid leukemia and myelodysplasia. Leuk. Lymphoma 2008, 49, 543–553. [Google Scholar] [CrossRef] [PubMed]
- Pastan, I.; FitzGerald, D. Recombinant Toxins for Cancer Treatment. Science 1991, 254, 1173–1177. [Google Scholar] [CrossRef]
- Tong, W.; Dwyer, C.A.; Thacker, B.E.; Glass, C.A.; Brown, J.R.; Hamill, K.; Moremen, K.W.; Sarrazin, S.; Gordts, P.L.; Dozier, L.E.; et al. Guanidinylated Neomycin Conjugation Enhances Intranasal Enzyme Replacement in the Brain. Mol. Ther. 2017, 25, 2743–2752. [Google Scholar] [CrossRef]
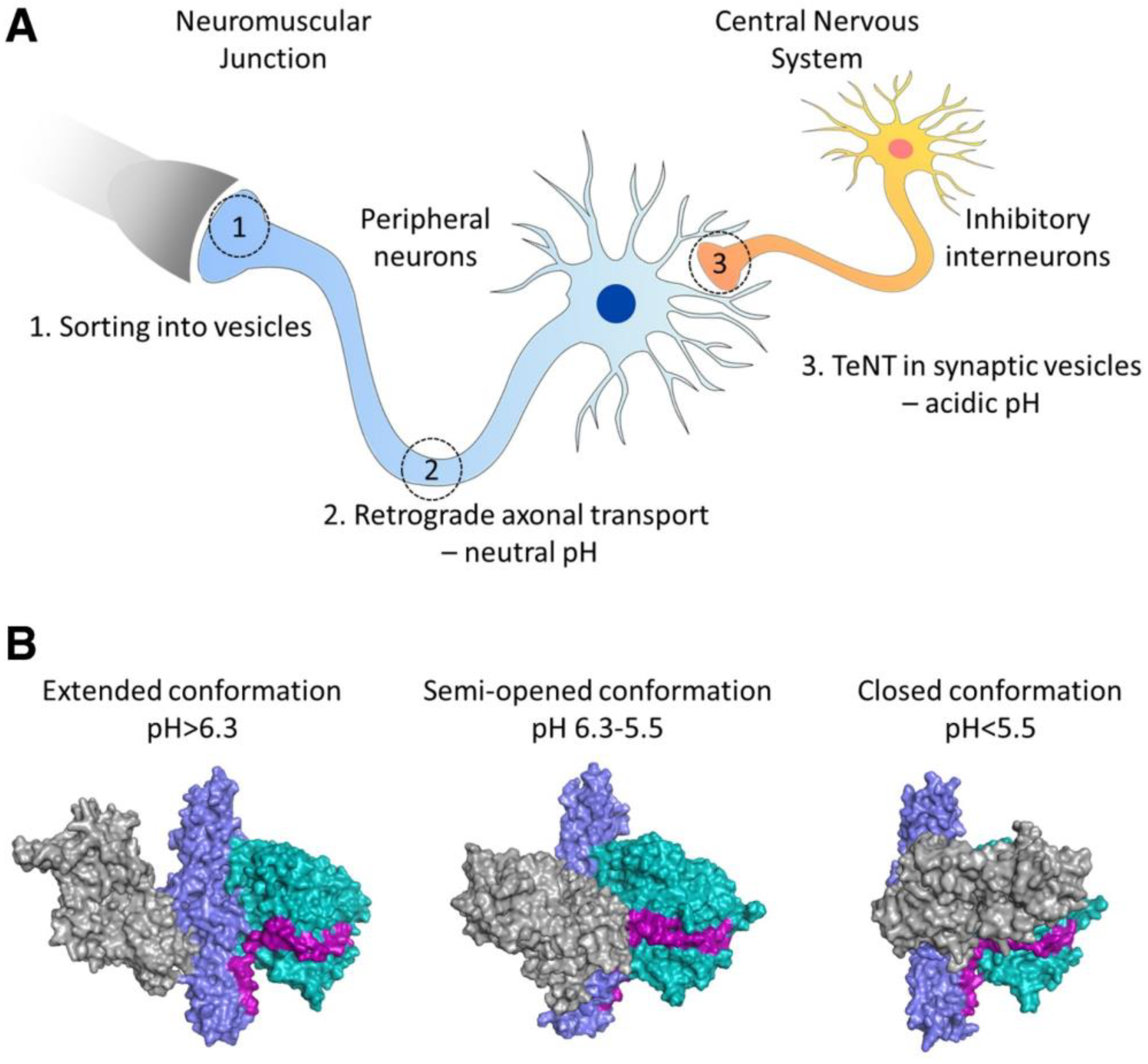
- Masuyer, G.; Conrad, J.; Stenmark, P. The structure of the tetanus toxin reveals pH-mediated domain dynamics. EMBO Rep. 2017, 18, 1306–1317. [Google Scholar] [CrossRef]
- Bizzini, B.; Stoeckel, K.; Schwab, M. An Antigenic Polypeptide Fragment Isolated from Tetanus Toxin: Chemical Characterization, Binding to Gangliosides and Retrograde Axonal Transport in Various Neuron Systems. J. Neurochem. 1977, 28, 529–542. [Google Scholar] [CrossRef]
- Fishman, P.S.; Savitt, J.M.; Farrand, D.A. Enhanced CNS uptake of systemically administered proteins through conjugation with tetanus C-fragment. J. Neurol. Sci. 1990, 98, 311–325. [Google Scholar] [CrossRef]
- Figueiredo, D.M.; Hallewell, R.A.; Chen, L.L.; Fairweather, N.F.; Dougan, G.; Savitt, J.M.; Parks, D.A.; Fishman, P.S. Delivery of Recombinant Tetanus–Superoxide Dismutase Proteins to Central Nervous System Neurons by Retrograde Axonal Transport. Exp. Neurol. 1997, 145, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Chian, R.-J.; Li, J.; Ay, I.; Celia, S.A.; Kashi, B.B.; Tamrazian, E.; Matthews, J.C.; Bronson, R.T.; Rossomando, A.; Pepinsky, R.B.; et al. IGF-1:Tetanus toxin fragment C fusion protein improves delivery of IGF-1 to spinal cord but fails to prolong survival of ALS mice. Brain Res. 2009, 1287, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Toivonen, J.M.; Oliván, S.; Osta, R. Tetanus Toxin C-Fragment: The Courier and the Cure? Toxins 2010, 2, 2622–2644. [Google Scholar] [CrossRef] [PubMed]
- Bailon, P.; Won, C.-Y. PEG-modified biopharmaceuticals. Expert Opin. Drug Deliv. 2009, 6, 1–16. [Google Scholar] [CrossRef]
- Brem, H.; Piantadosi, S.; Burger, P.C.; Walker, M.; Selker, R.; Vick, N.A.; Black, K.; Sisti, M.; Brem, S.; Mohr, G.; et al. Placebo-controlled trial of safety and efficacy of intraoperative controlled delivery by biodegradable polymers of chemotherapy for recurrent gliomas. Lancet 1995, 345, 1008–1012. [Google Scholar] [CrossRef]
- Tsai, N.-M.; Chen, Y.-L.; Lee, C.-C.; Lin, P.-C.; Cheng, Y.-L.; Chang, W.-L.; Lin, S.-Z.; Harn, H.-J. The natural compound n-butylidenephthalide derived from Angelica sinensis inhibits malignant brain tumor growth in vitro and in vivo3. J. Neurochem. 2006, 99, 1251–1262. [Google Scholar] [CrossRef]
- A Phase I/IIa Study of Cerebraca Wafer Plus Adjuvant Temozolomide (TMZ) in Patients with Recurrent High Grade Glioma. Available online: https://clinicaltrials.gov/ct2/show/NCT03234595 (accessed on 22 April 2023).
- Zhang, J.; Chen, C.; Li, A.; Jing, W.; Sun, P.; Huang, X.; Liu, Y.; Zhang, S.; Du, W.; Zhang, R.; et al. Immunostimulant hydrogel for the inhibition of malignant glioma relapse post-resection. Nat. Nanotechnol. 2021, 16, 538–548. [Google Scholar] [CrossRef]
- Xu, H.-L.; Tian, F.-R.; Lu, C.-T.; Xu, J.; Fan, Z.-L.; Yang, J.-J.; Chen, P.-P.; Huang, Y.-D.; Xiao, J.; Zhao, Y.-Z. Thermo-sensitive hydrogels combined with decellularised matrix deliver bFGF for the functional recovery of rats after a spinal cord injury. Sci. Rep. 2016, 6, 38332. [Google Scholar] [CrossRef]
- Xu, H.-L.; Tian, F.-R.; Xiao, J.; Chen, P.-P.; Xu, J.; Fan, Z.-L.; Yang, J.-J.; Lu, C.-T.; Zhao, Y.-Z. Sustained-release of FGF-2 from a hybrid hydrogel of heparin-poloxamer and decellular matrix promotes the neuroprotective effects of proteins after spinal injury. Int. J. Nanomed. 2018, 13, 681–694. [Google Scholar] [CrossRef] [PubMed]
- Donaghue, I.E.; Tator, C.H.; Shoichet, M.S. Local Delivery of Neurotrophin-3 and Anti-NogoA Promotes Repair After Spinal Cord Injury. Tissue Eng. Part A 2016, 22, 733–741. [Google Scholar] [CrossRef]
- Nguyen, L.H.; Gao, M.; Lin, J.; Wu, W.; Wang, J.; Chew, S.Y. Three-dimensional aligned nanofibers-hydrogel scaffold for controlled non-viral drug/gene delivery to direct axon regeneration in spinal cord injury treatment. Sci. Rep. 2017, 7, 42212. [Google Scholar] [CrossRef] [PubMed]
- Pakulska, M.M.; Tator, C.H.; Shoichet, M.S. Local delivery of chondroitinase ABC with or without stromal cell-derived factor 1α promotes functional repair in the injured rat spinal cord. Biomaterials 2017, 134, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, H.; Xu, H.; Zhao, Y.; Li, Z.; Li, J.; Wang, H.; Zhuge, D.; Guo, X.; Xu, H.; et al. Novel multi-drug delivery hydrogel using scar-homing liposomes improves spinal cord injury repair. Theranostics 2018, 8, 4429–4446. [Google Scholar] [CrossRef]
- Ghosh, B.; Wang, Z.; Nong, J.; Urban, M.W.; Zhang, Z.; Trovillion, V.A.; Wright, M.C.; Zhong, Y.; Lepore, A.C. Local BDNF Delivery to the Injured Cervical Spinal Cord using an Engineered Hydrogel Enhances Diaphragmatic Respiratory Function. J. Neurosci. 2018, 38, 5982–5995. [Google Scholar] [CrossRef]
- He, Z.; Zang, H.; Zhu, L.; Huang, K.; Yi, T.; Zhang, S.; Cheng, S. An anti-inflammatory peptide and brain-derived neurotrophic factor-modified hyaluronan-methylcellulose hydrogel promotes nerve regeneration in rats with spinal cord injury. Int. J. Nanomed. 2019, 14, 721–732. [Google Scholar] [CrossRef]
- Lescure, F.; Seguin, C.; Breton, P.; Bourrinet, P.; Roy, D.; Couvreur, P. Preparation and Characterization of Novel Poly(methylidene Malonate 2.1.2.)-Made Nanoparticles. Pharm. Res. 1994, 11, 1270–1277. [Google Scholar] [CrossRef]
- Fournier, E.; Passirani, C.; Colin, N.; Sagodira, S.; Menei, P.; Benoit, J.-P.; Montero-Menei, C.N. The brain tissue response to biodegradable poly(methylidene malonate 2.1.2)-based microspheres in the rat. Biomaterials 2006, 27, 4963–4974. [Google Scholar] [CrossRef]
- Hsiao, C.-Y.; Liu, S.-J.; Ueng, S.W.-N.; Chan, E.-C. The influence of γ irradiation and ethylene oxide treatment on the release characteristics of biodegradable poly(lactide-co-glycolide) composites. Polym. Degrad. Stab. 2012, 97, 715–720. [Google Scholar] [CrossRef]
- Wang, T.; Suita, Y.; Miriyala, S.; Dean, J.; Tapinos, N.; Shen, J. Advances in Lipid-Based Nanoparticles for Cancer Chemoimmunotherapy. Pharmaceutics 2021, 13, 520. [Google Scholar] [CrossRef] [PubMed]
- Pilkington, E.H.; Suys, E.J.; Trevaskis, N.L.; Wheatley, A.K.; Zukancic, D.; Algarni, A.; Al-Wassiti, H.; Davis, T.P.; Pouton, C.W.; Kent, S.J.; et al. From influenza to COVID-19: Lipid nanoparticle mRNA vaccines at the frontiers of infectious diseases. Acta Biomater. 2021, 131, 16–40. [Google Scholar] [CrossRef] [PubMed]
- Pakulska, M.M.; Donaghue, I.E.; Obermeyer, J.M.; Tuladhar, A.; McLaughlin, C.K.; Shendruk, T.N.; Shoichet, M.S. Encapsulation-free controlled release: Electrostatic adsorption eliminates the need for protein encapsulation in PLGA nanoparticles. Sci. Adv. 2016, 2, e1600519. [Google Scholar] [CrossRef] [PubMed]
- Donaghue, I.E.; Tator, C.H.; Shoichet, M.S. Sustained delivery of bioactive neurotrophin-3 to the injured spinal cord. Biomater. Sci. 2015, 3, 65–72. [Google Scholar] [CrossRef]
- Wang, X.; Bodman, A.; Shi, C.; Guo, D.; Wang, L.; Luo, J.; Hall, W.A. Tunable Lipidoid-Telodendrimer Hybrid Nanoparticles for Intracellular Protein Delivery in Brain Tumor Treatment. Small 2016, 12, 4185–4192. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, J.; Feng, C.; Shao, X.; Liu, Q.; Zhang, Q.; Pang, Z.; Jiang, X. Intranasal nanoparticles of basic fibroblast growth factor for brain delivery to treat Alzheimer’s disease. Int. J. Pharm. 2014, 461, 192–202. [Google Scholar] [CrossRef]
- Zhao, Y.-Z.; Li, X.; Lu, C.-T.; Lin, M.; Chen, L.-J.; Xiang, Q.; Zhang, M.; Jin, R.-R.; Jiang, X.; Shen, X.-T.; et al. Gelatin nanostructured lipid carriers-mediated intranasal delivery of basic fibroblast growth factor enhances functional recovery in hemiparkinsonian rats. Nanomed. Nanotechnol. Biol. Med. 2014, 10, 755–764. [Google Scholar] [CrossRef]
- Gartziandia, O.; Herran, E.; Pedraz, J.L.; Carro, E.; Igartua, M.; Hernandez, R.M. Chitosan coated nanostructured lipid carriers for brain delivery of proteins by intranasal administration. Colloids Surf. B Biointerfaces 2015, 134, 304–313. [Google Scholar] [CrossRef]
- Ribovski, L.; Hamelmann, N.M.; Paulusse, J.M.J. Polymeric Nanoparticles Properties and Brain Delivery. Pharmaceutics 2021, 13, 2045. [Google Scholar] [CrossRef]
- Khojasteh, A.; Oraee-Yazdani, S.; Dehghani, L.; Soleimani, M.; Keshel, S.H.; Saadatnia, M.; Saboori, M.; Zali, A.; Hashemi, S.M.; Soleimani, R. Safety of intraparenchymal injection of allogenic placenta mesenchymal stem cells derived exosome in patients undergoing decompressive craniectomy following malignant middle cerebral artery infarct, a pilot randomized clinical trial. Int. J. Prev. Med. 2022, 13, 7. [Google Scholar] [CrossRef]
- The Safety and the Efficacy Evaluation of Allogenic Adipose MSC-Exos in Patients with Alzheimer’s Disease. Identifier NCT04388982. 2020. Available online: https://clinicaltrials.gov/ct2/show/NCT04388982 (accessed on 17 April 2023).
- Mohammed, I.; Ijaz, S.; Mokhtari, T.; Gholaminejhad, M.; Mahdavipour, M.; Jameie, B.; Akbari, M.; Hassanzadeh, G. Subventricular zone-derived extracellular vesicles promote functional recovery in rat model of spinal cord injury by inhibition of NLRP3 inflammasome complex formation. Metab. Brain Dis. 2020, 35, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Haney, M.J.; Klyachko, N.L.; Zhao, Y.; Gupta, R.; Plotnikova, E.G.; He, Z.; Patel, T.; Piroyan, A.; Sokolsky, M.; Kabanov, A.V.; et al. Exosomes as drug delivery vehicles for Parkinson’s disease therapy. J. Control. Release 2015, 207, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Hayes, S.H.; Liu, Q.; Selvakumaran, S.; Haney, M.J.; Batrakova, E.V.; Allman, B.L.; Walton, P.A.; Kiser, P.; Whitehead, S.N. Brain Targeting and Toxicological Assessment of the Extracellular Vesicle-Packaged Antioxidant Catalase-SKL Following Intranasal Administration in Mice. Neurotox. Res. 2021, 39, 1418–1429. [Google Scholar] [CrossRef]
- Hung, K.; Meitlis, I.; Hale, M.; Chen, C.-Y.; Singh, S.; Jackson, S.W.; Miao, C.H.; Khan, I.F.; Rawlings, D.J.; James, R.G. Engineering Protein-Secreting Plasma Cells by Homology-Directed Repair in Primary Human B Cells. Mol. Ther. 2018, 26, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Riglar, D.; Silver, P.A. Engineering bacteria for diagnostic and therapeutic applications. Nat. Rev. Genet. 2018, 16, 214–225. [Google Scholar] [CrossRef]
- Brown, C.E.; Badie, B.; Barish, M.E.; Weng, L.; Ostberg, J.R.; Chang, W.-C.; Naranjo, A.; Starr, R.; Wagner, J.; Wright, C.; et al. Bioactivity and Safety of IL13Rα2-Redirected Chimeric Antigen Receptor CD8+ T Cells in Patients with Recurrent Glioblastoma. Clin. Cancer Res. 2015, 21, 4062–4072. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Brawley, V.; Hegde, M.; Bielamowicz, K.; Kalra, M.; Landi, D.; Robertson, C.; Gray, T.L.; Diouf, O.; Wakefield, A.; et al. HER2-Specific Chimeric Antigen Receptor–Modified Virus-Specific T Cells for Progressive Glioblastoma: A Phase 1 Dose-Escalation Trial. JAMA Oncol. 2017, 3, 1094–1101. [Google Scholar] [CrossRef]
- Goff, S.L.; Morgan, R.A.; Yang, J.C.; Sherry, R.M.; Robbins, P.F.; Restifo, N.P.; Feldman, S.A.; Lu, Y.-C.; Lu, L.; Zheng, Z.; et al. Pilot Trial of Adoptive Transfer of Chimeric Antigen Receptor–transduced T Cells Targeting EGFRvIII in Patients with Glioblastoma. J. Immunother. 2019, 42, 126–135. [Google Scholar] [CrossRef]
- Portnow, J.; Synold, T.W.; Badie, B.; Tirughana, R.; Lacey, S.F.; D’Apuzzo, M.; Metz, M.Z.; Najbauer, J.; Bedell, V.; Vo, T.; et al. Neural Stem Cell–Based Anticancer Gene Therapy: A First-in-Human Study in Recurrent High-Grade Glioma Patients. Clin. Cancer Res. 2017, 23, 2951–2960. [Google Scholar] [CrossRef]
- Killer, K.; Le, O.; Beauséjour, C. The Intracerebroventricular Injection of Murine Mesenchymal Stromal Cells Engineered to Secrete Epidermal Growth Factor Does Not Prevent Loss of Neurogenesis in Irradiated Mice. Radiat. Res. 2021, 196, 315–322. [Google Scholar] [CrossRef]
- Aronson, J.; Katnani, H.; Pomerantseva, I.; Shapir, N.; Tse, H.; Miari, R.; Goltsman, H.; Mwizerwa, O.; Neville, C.; Neil, G.; et al. Sustained intrathecal therapeutic protein delivery using genetically transduced tissue implants in a freely moving rat model. Int. J. Pharm. 2017, 534, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.K.; Schwendeman, S.P. Characterization of Octreotide–PLGA Binding by Isothermal Titration Calorimetry. Biomacromolecules 2020, 21, 4087–4093. [Google Scholar] [CrossRef] [PubMed]
- Kreitz, J.; Friedrich, M.J.; Guru, A.; Lash, B.; Saito, M.; Macrae, R.K.; Zhang, F. Programmable protein delivery with a bacterial contractile injection system. Nature 2023, 616, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Shiue, S.-J.; Rau, R.-H.; Shiue, H.-S.; Hung, Y.-W.; Li, Z.-X.; Yang, K.D.; Cheng, J.-K. Mesenchymal stem cell exosomes as a cell-free therapy for nerve injury–induced pain in rats. Pain 2018, 160, 210–223. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pros | Cons | Clinical Applications | |
|---|---|---|---|
| ICV | Direct introduction of nearly 100% therapeutics into the CSF in the lateral ventricle with minimal drug-protein bounding. | Low drug distribution in the brain parenchyma that is away from the injection site; Bulk flow of CSF may result in fast clearance and systemic exposure; Complications associated with ICV device implantation. | Brineura [16]; tripeptidyl peptidase 1 (Phase I/II) [17,18]; idursulfase-β (Phase I/II) [19]; telbermin (Phase I) [20] |
| CED | Improved drug distribution in the brain parenchyma; Topographically more restricted, thus less complications compared to ICV. | Inefficient diffusion in the parenchyma for some macromolecules; Complications from the invasiveness of the CED. | Omburtamab (Phase I) [21,22] |
| IT | Direct injection of nearly 100% therapeutics into the CSF in the spinal subarachnoid space with long half-life in the CSF due to minimal drug-protein bounding and metabolism; Less invasive compared to ICV and CED | Less macromolecule distribution in the parenchyma compared to ICV and CED; Potential complications from IT device implantation. | Recombinant human arylsulfatase A(Phase I/II) [23] |
| IN | Noninvasive | Potential systemic exposure; Device design is critical. | Insulin & insulin detemir (Phase II) [6]; neuropeptide Y (Phase I) [7] |
| Disease Model | Formulation | Cargo Therapeutic | Carrier Material | Release Duration | Ref. |
|---|---|---|---|---|---|
| GBM (GL261 mouse model) | Hydrogel | CXCL-10 | Self-assembled oligopeptide | >12 days | [73] |
| Spinal cord injury (Rat model) | Hydrogel | basic fibroblast growth factor (bFGF) or FGF2 | Heparin-modified poloxamer and lyophilized acellular spinal cord | >7 days | [74,75] |
| Nanoparticle (NP)-hydrogel composite | neurotrophin-3 (NT-3) + antibody 11c7 (anti-NogoA) | Hyaluronan-methylcellulose (HAMC) and poly(lactic-co-glycolic acid) (PLGA) | NT-3: >58 days; anti-NogoA: >10 days | [76] | |
| Hydrogel | NT-3 | Heparin-contained collagen | >90 days | [77] | |
| NP-hydrogel composite | chondroitinase ABC (ChABC) + stromal cell-derived factor 1α (SDF) | SH3 binding peptide-modified methylcellulose and PLGA | ChABC: >7 days; SDF: >14 days | [78] | |
| Liposome-hydrogel composite | brain-derived neurotrophic factor (BDNF) + acidic FGF (aFGF) | Heparin-modified poloxamer and scar-targeted tetrapeptide-modified liposomes | BDNF/aFGF: >21 days | [79] | |
| NP-hydrogel composite | BDNF | Agarose + polysaccharide polyelectrolyte complexes | >17 days | [80] | |
| Hydrogel | Anti-inflammatory peptide KAFAK + BDNF | HAMC | KAFAK/BDNF: >4 days (only ~50% of payloads released) | [81] |
| Disease Model | Formulation | Cargo Therapeutic | Administration | Ref. |
|---|---|---|---|---|
| Spinal cord injury (Rat model) | PLGA NP | NT-3 | IT | [88] |
| Brain tumor (Mouse model) | Lipidoid-telodendrimer binary hybrid NP | DT390 | CED | [89] |
| AD (Rat model) | Lectin-modified PEG-PLGA NP | bFGF | IN | [90] |
| PD (Rat model) | Phospholipid-based gelatin NP | bFGF | IN | [91] |
| Mouse model | Chitosan coated nanostructured lipid carriers | human IGF-I (hIGF-I) | IN | [92] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, W.; Shen, J. Local Delivery Strategies for Peptides and Proteins into the CNS: Status Quo, Challenges, and Future Perspectives. Pharmaceuticals 2023, 16, 810. https://doi.org/10.3390/ph16060810
Yue W, Shen J. Local Delivery Strategies for Peptides and Proteins into the CNS: Status Quo, Challenges, and Future Perspectives. Pharmaceuticals. 2023; 16(6):810. https://doi.org/10.3390/ph16060810
Chicago/Turabian StyleYue, Weizhou, and Jie Shen. 2023. "Local Delivery Strategies for Peptides and Proteins into the CNS: Status Quo, Challenges, and Future Perspectives" Pharmaceuticals 16, no. 6: 810. https://doi.org/10.3390/ph16060810