
Pregnenolone Inhibits Doxorubicin-Induced Cardiac Oxidative Stress, Inflammation, and Apoptosis—Role of Matrix Metalloproteinase 2 and NADPH Oxidase 1
 , ,
, ,  , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Results
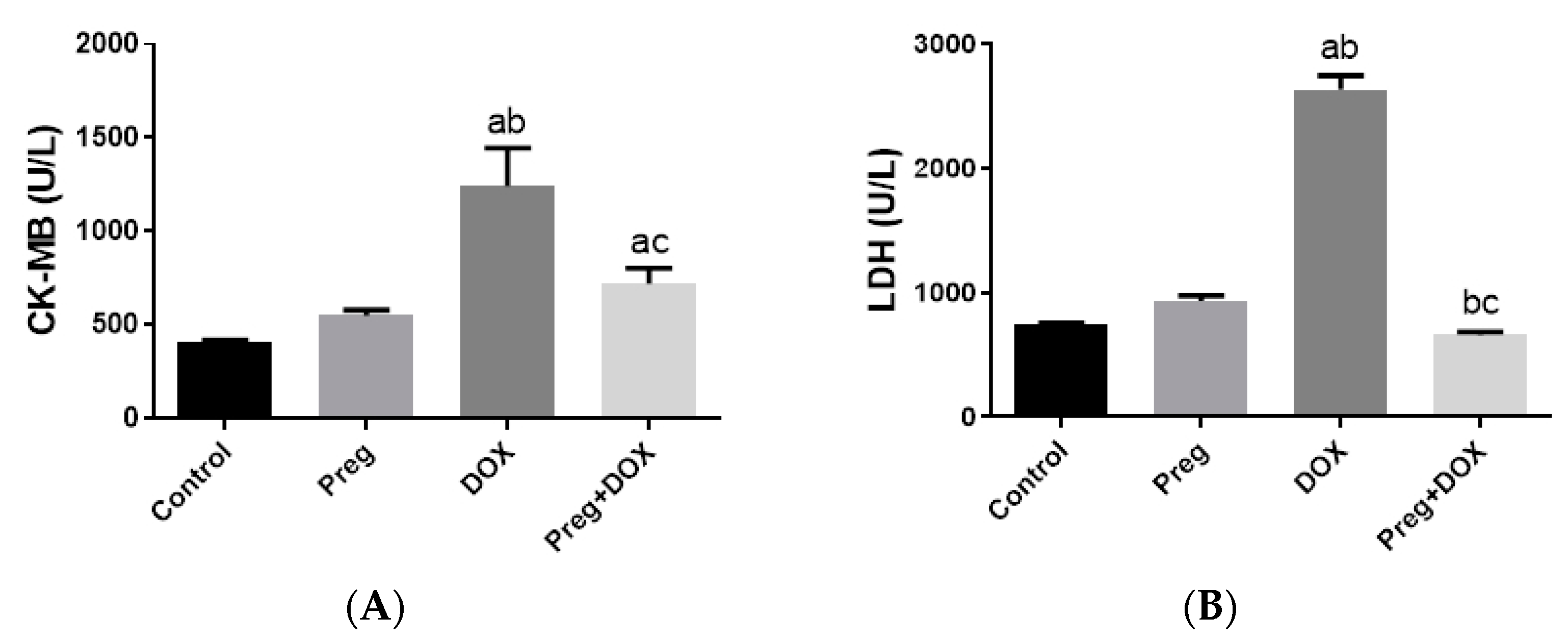
2.1. Pregnenolone Protects against DOX-Induced Cardiac Injury in Rats
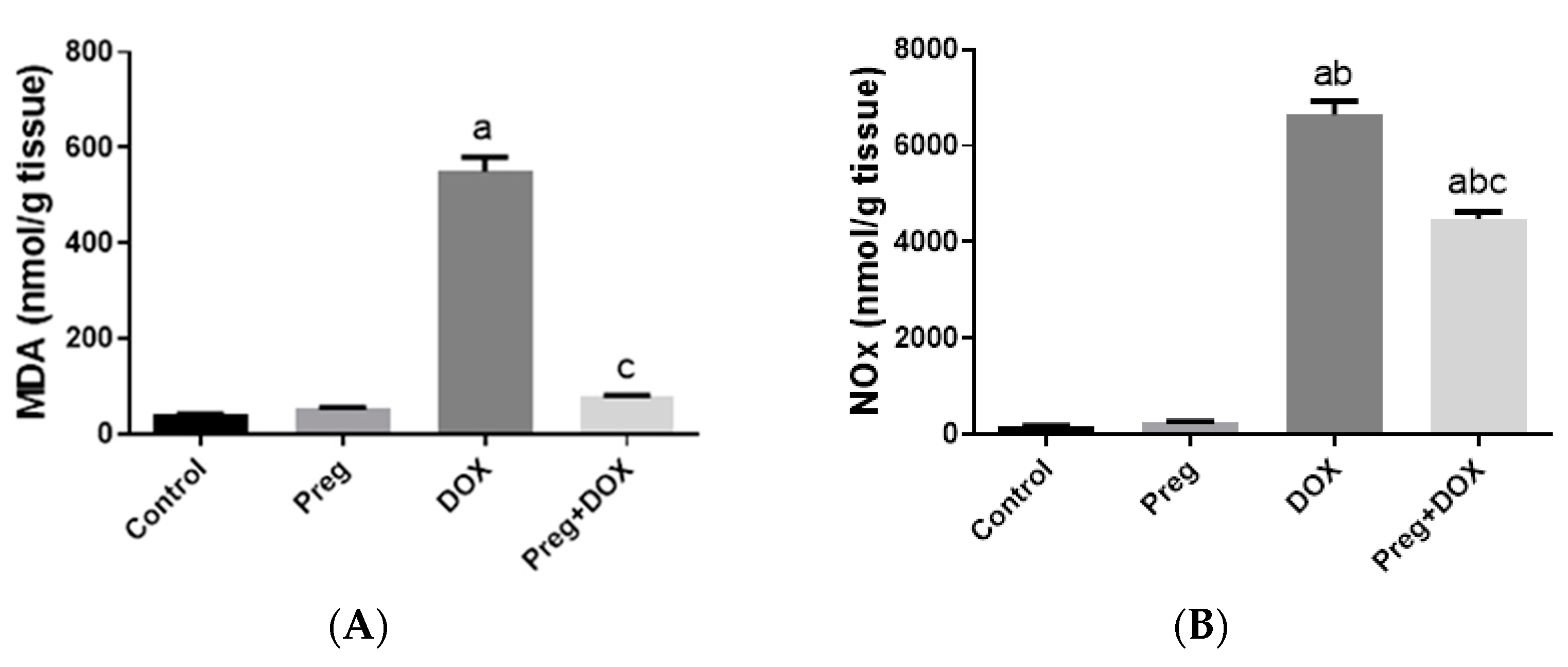
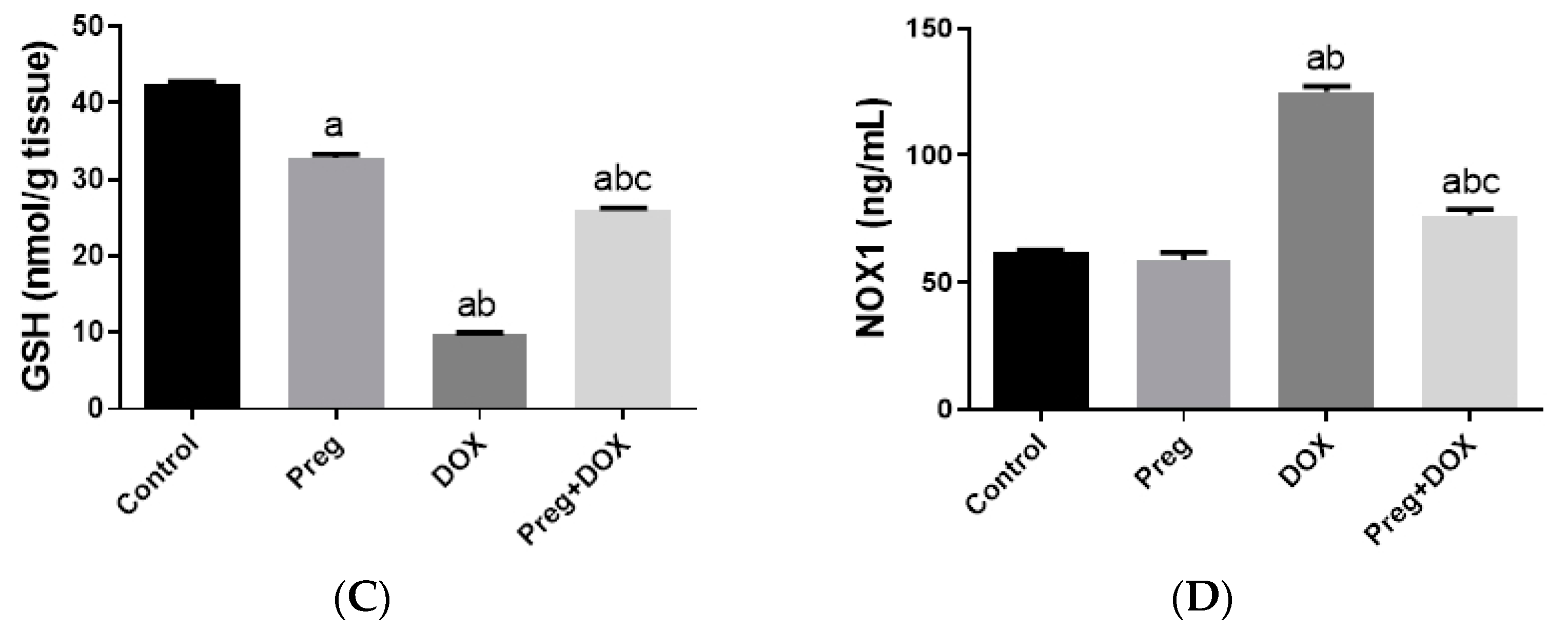
2.2. Pregnenolone Prevents DOX-Induced Myocardial Oxidative Stress
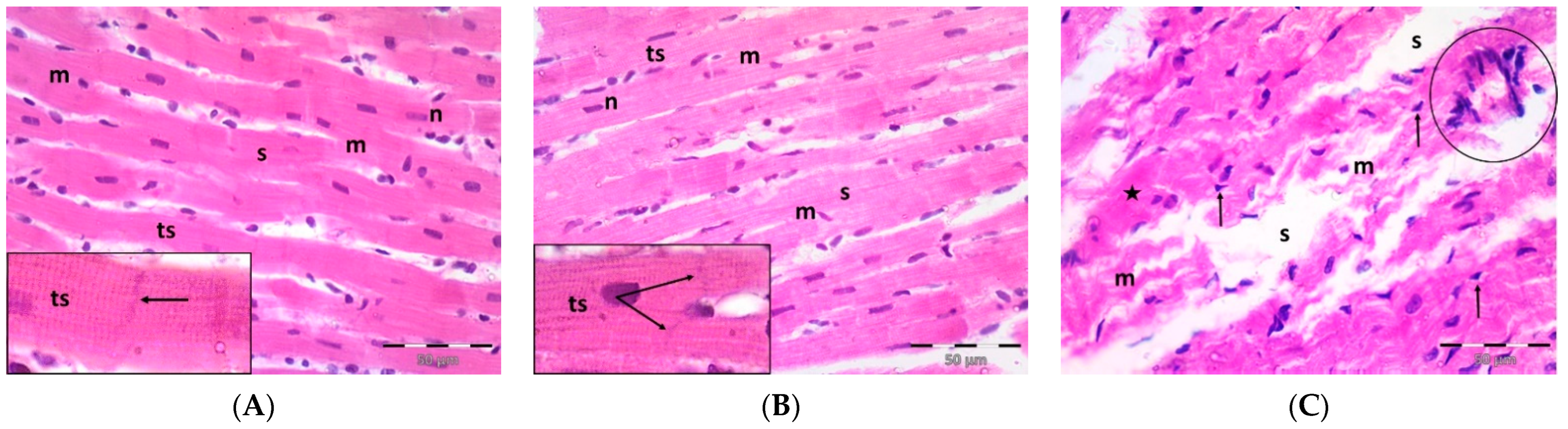
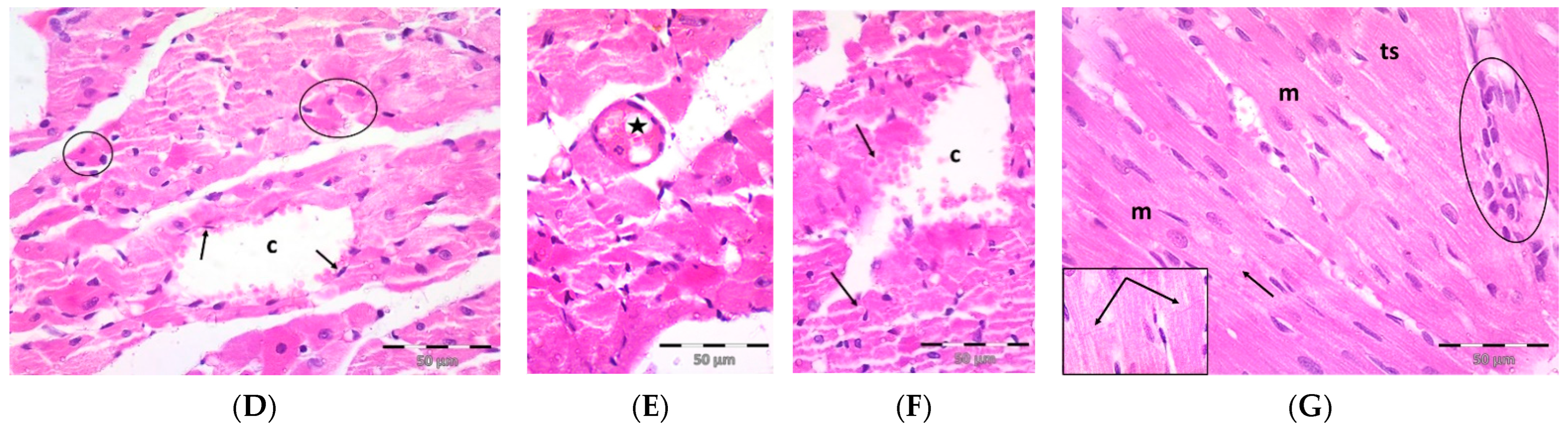
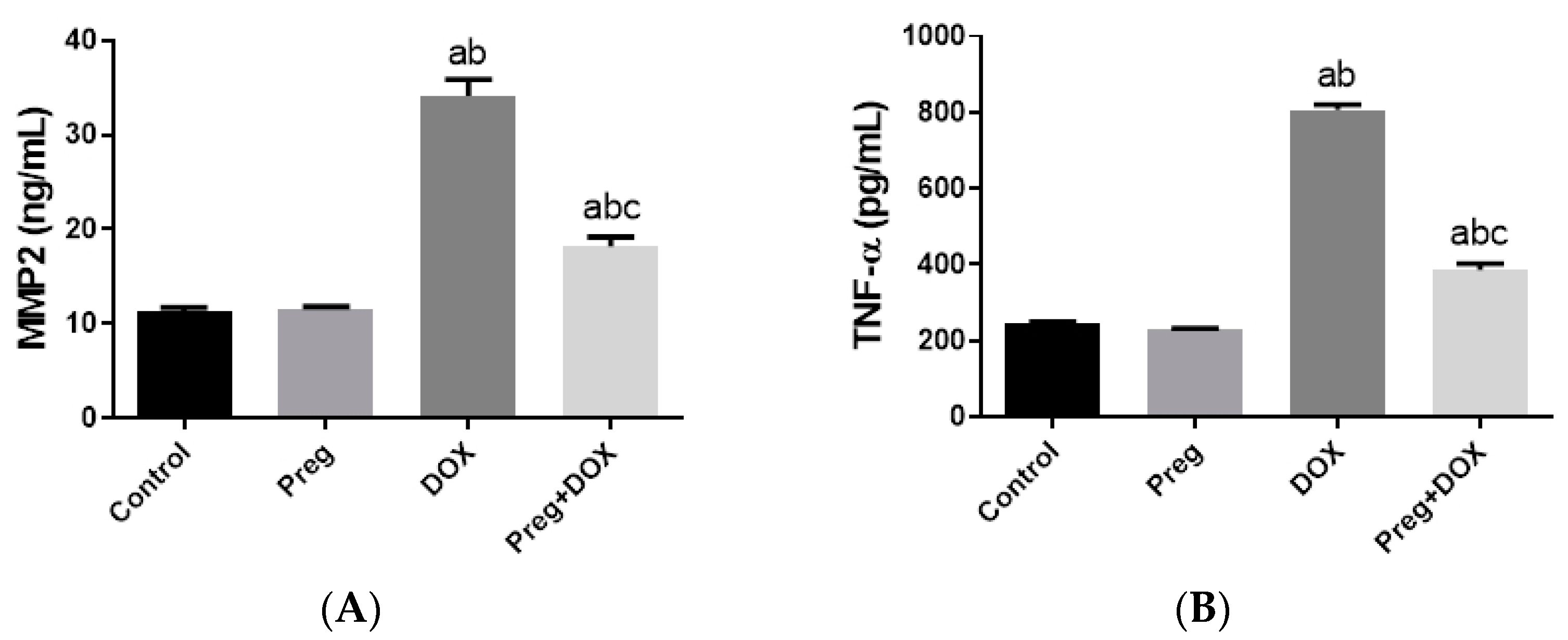
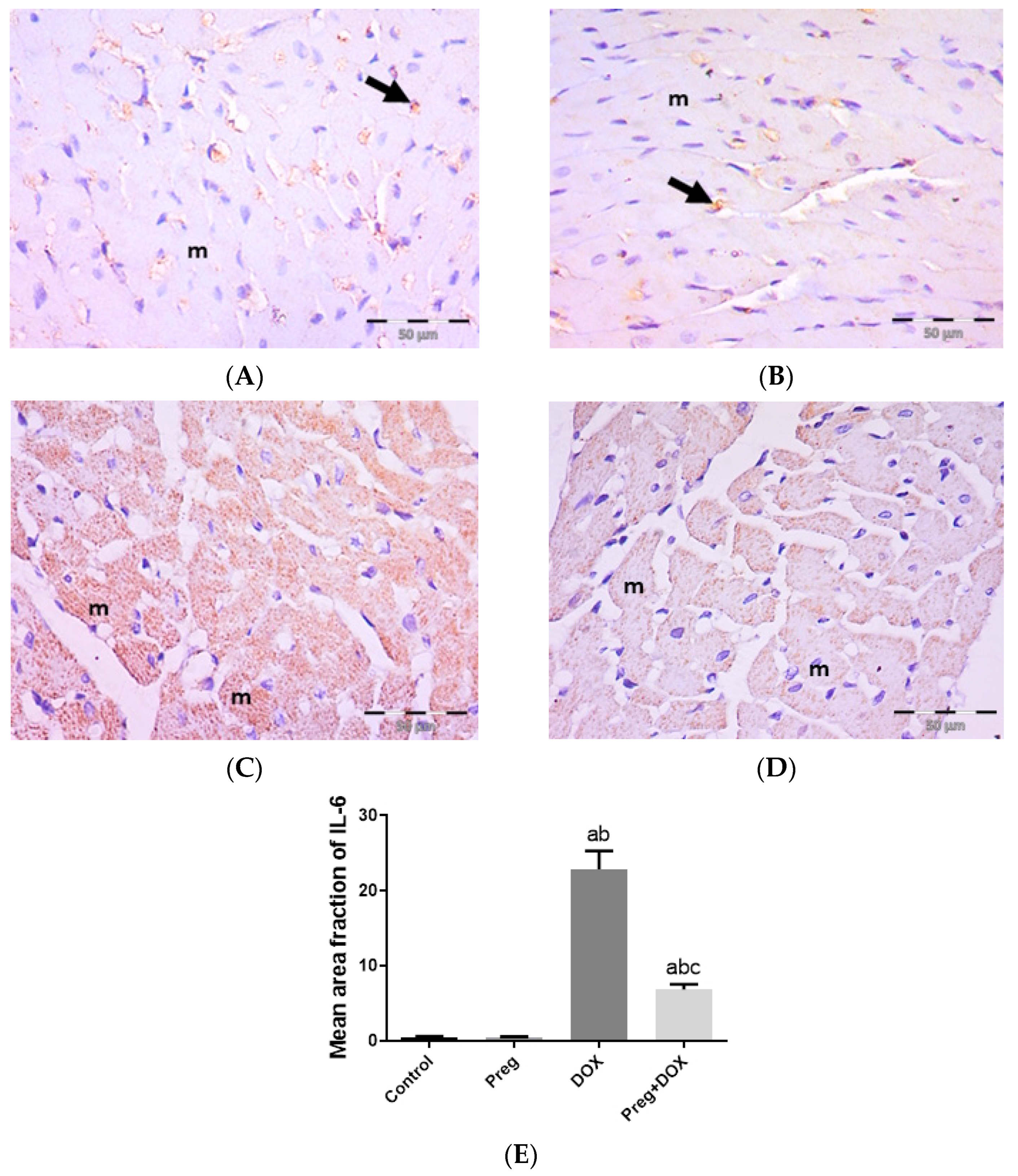
2.3. Pregnenolone Protects against DOX-Induced Cardiac Remodeling and Inflammation
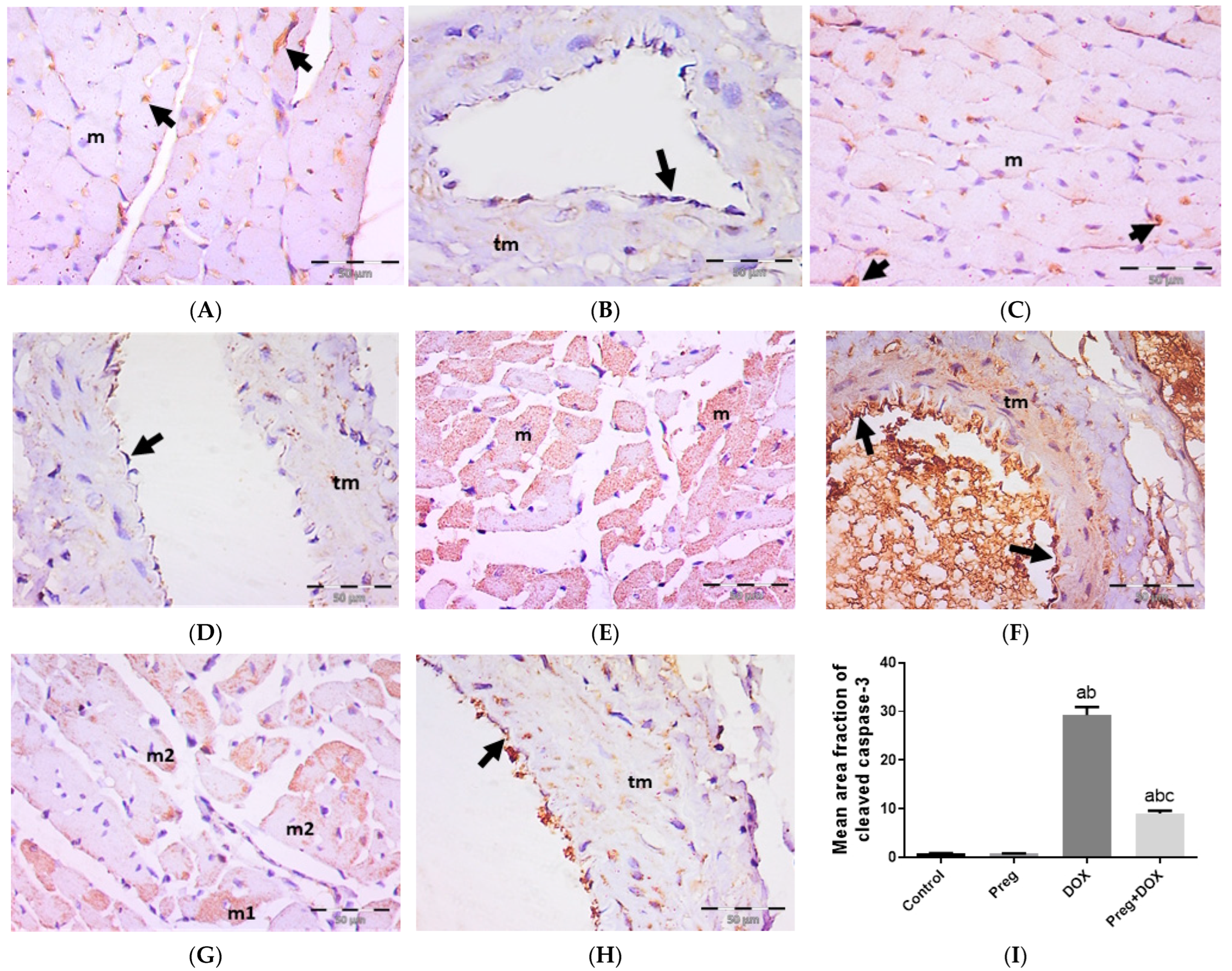
2.4. Pregnenolone Protects against DOX-Induced Activation of Apoptosis
3. Discussion
4. Materials and Methods
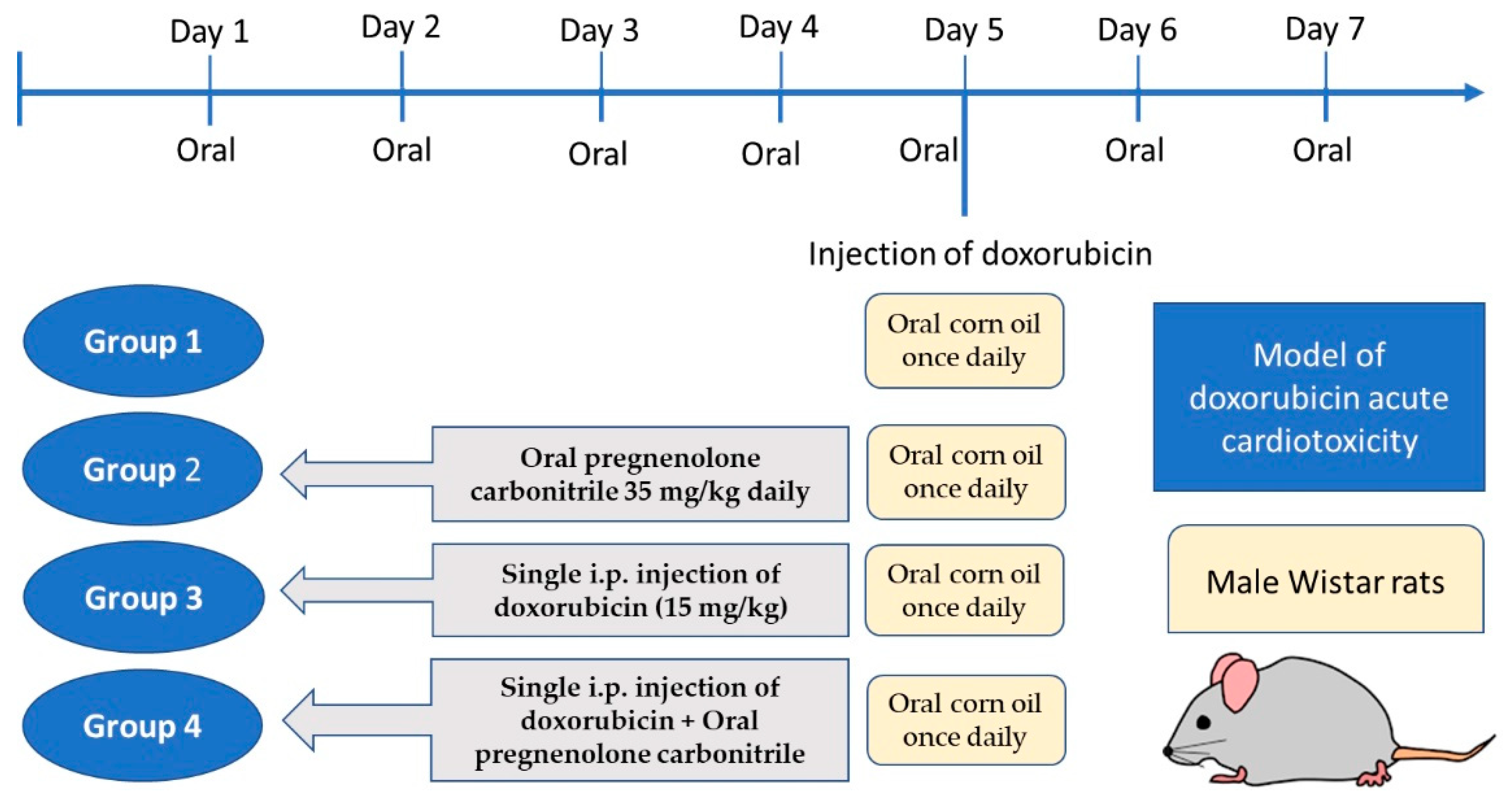
4.1. Experimental Animals and Study Design
4.2. Sample Collection
4.3. Determination of Serum CK-MB, LDH, and Redox State
4.4. Determination of Cardiac NOX1, MMP2, and TNF-α Levels
4.5. Histopathology and Immunohistochemistry
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gadducci, A.; Cosio, S. Trabectedin and lurbinectedin: Mechanisms of action, clinical impact, and future perspectives in uterine and soft tissue sarcoma, ovarian carcinoma, and endometrial carcinoma. Front. Oncol. 2022, 12, 914342. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Wu, Y.; Chen, S.; Wu, J.; Zhu, W.; Liu, H.; Chen, M.; Xu, B. Curative Effect and Survival Assessment Comparing Gemcitabine and Cisplatin Versus Methotrexate, Vinblastine, Doxorubicin and Cisplatin as Neoadjuvant Therapy for Bladder Cancer: A Systematic Review and Meta-Analysis. Front. Oncol. 2021, 11, 678896. [Google Scholar] [CrossRef] [PubMed]
- Bagdasaryan, A.A.; Chubarev, V.N.; Smolyarchuk, E.A.; Drozdov, V.N.; Krasnyuk, I.I.; Liu, J.; Fan, R.; Tse, E.; Shikh, E.V.; Sukocheva, O.A. Pharmacogenetics of Drug Metabolism: The Role of Gene Polymorphism in the Regulation of Doxorubicin Safety and Efficacy. Cancers 2022, 14, 5436. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.M.; Zavorka Thomas, M.; Magdy, T.; Eisenmann, E.D.; Uddin, M.E.; DiGiacomo, D.F.; Pan, A.; Keiser, M.; Otter, M.; Xia, S.H. Targeting OCT3 attenuates doxorubicin-induced cardiac injury. Proc. Natl. Acad. Sci. USA 2021, 118, e2020168118. [Google Scholar] [CrossRef]
- Chen, Y.; Shi, S.; Dai, Y. Research progress of therapeutic drugs for doxorubicin-induced cardiomyopathy. Biomed. Pharmacother. 2022, 156, 113903. [Google Scholar] [CrossRef]
- Jones, I.C.; Dass, C.R. Doxorubicin-induced cardiotoxicity: Causative factors and possible interventions. J. Pharm. Pharmacol. 2022, 74, 1677–1688. [Google Scholar] [CrossRef]
- Stansfeld, A.; Radia, U.; Goggin, C.; Mahalingam, P.; Benson, C.; Napolitano, A.; Jones, R.L.; Rosen, S.D.; Karavasilis, V. Pharmacological strategies to reduce anthracycline-associated cardiotoxicity in cancer patients. Expert Opin. Pharmacother. 2022, 23, 1641–1650. [Google Scholar] [CrossRef]
- Wang, Y.C.; Wang, L.T.; Hung, T.I.; Hong, Y.R.; Chen, C.H.; Ho, C.J.; Wang, C. Severe cellular stress drives apoptosis through a dual control mechanism independently of p53. Cell Death Discov. 2022, 8, 282. [Google Scholar] [CrossRef]
- Rawat, P.S.; Jaiswal, A.; Khurana, A.; Bhatti, J.S.; Navik, U. Doxorubicin-induced cardiotoxicity: An update on the molecular mechanism and novel therapeutic strategies for effective management. Biomed. Pharmacother. 2021, 139, 111708. [Google Scholar] [CrossRef]
- Jong, J.; Pinney, J.R.; Packard, R.R.S. Anthracycline-induced cardiotoxicity: From pathobiology to identification of molecular targets for nuclear imaging. Front. Cardiovasc. Med. 2022, 9, 919719. [Google Scholar] [CrossRef]
- Seara, F.A.C.; Kasai-Brunswick, T.H.; Nascimento, J.H.M.; Campos-de-Carvalho, A.C. Anthracycline-induced cardiotoxicity and cell senescence: New therapeutic option? Cell. Mol. Life Sci. 2022, 79, 568. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Liu, D.; Song, H.; Tian, X.; Yan, C.; Han, Y. Overexpression of Kininogen-1 aggravates oxidative stress and mitochondrial dysfunction in DOX-induced cardiotoxicity. Biochem. Biophys. Res. Commun. 2021, 550, 142–150. [Google Scholar] [CrossRef]
- Mizuta, Y.; Akahoshi, T.; Eto, H.; Hyodo, F.; Murata, M.; Tokuda, K.; Eto, M.; Yamaura, K. Noninvasive Diagnosis of the Mitochondrial Function of Doxorubicin-Induced Cardiomyopathy Using In Vivo Dynamic Nuclear Polarization-Magnetic Resonance Imaging. Antioxidants 2022, 11, 1454. [Google Scholar] [CrossRef] [PubMed]
- Kajarabille, N.; Latunde-Dada, G.O. Programmed Cell-Death by Ferroptosis: Antioxidants as Mitigators. Int. J. Mol. Sci. 2019, 20, 4968. [Google Scholar] [CrossRef] [PubMed]
- Kourek, C.; Touloupaki, M.; Rempakos, A.; Loritis, K.; Tsougkos, E.; Paraskevaidis, I.; Briasoulis, A. Cardioprotective Strategies from Cardiotoxicity in Cancer Patients: A Comprehensive Review. J. Cardiovasc. Dev. Dis. 2022, 9, 259. [Google Scholar] [CrossRef] [PubMed]
- Luu, A.Z.; Chowdhury, B.; Al-Omran, M.; Teoh, H.; Hess, D.A.; Verma, S. Role of Endothelium in Doxorubicin-Induced Cardiomyopathy. JACC Basic Transl. Sci. 2018, 3, 861–870. [Google Scholar] [CrossRef]
- He, H.; Wang, L.; Qiao, Y.; Zhou, Q.; Li, H.; Chen, S.; Yin, D.; Huang, Q.; He, M. Doxorubicin Induces Endotheliotoxicity and Mitochondrial Dysfunction via ROS/eNOS/NO Pathway. Front. Pharmacol. 2019, 10, 1531. [Google Scholar] [CrossRef]
- Hodjat, M.; Haller, H.; Dumler, I.; Kiyan, Y. Urokinase receptor mediates doxorubicin-induced vascular smooth muscle cell senescence via proteasomal degradation of TRF2. J. Vasc. Res. 2013, 50, 109–123. [Google Scholar] [CrossRef]
- França, A.S.; Rossoni, L.V.; Amaral, S.M.; Vassallo, D.V. Reactivity of the isolated perfused rat tail vascular bed. Braz. J. Med. Biol. Res. Rev. Bras. Pesqui. Med. Biol. 1997, 30, 891–895. [Google Scholar] [CrossRef]
- Bosman, M.; Krüger, D.N.; Favere, K.; Wesley, C.D.; Neutel, C.H.G.; Van Asbroeck, B.; Diebels, O.R.; Faes, B.; Schenk, T.J.; Martinet, W.; et al. Doxorubicin Impairs Smooth Muscle Cell Contraction: Novel Insights in Vascular Toxicity. Int. J. Mol. Sci. 2021, 22, 12812. [Google Scholar] [CrossRef]
- Jin, J.S.; Yao, C.W.; Chin, T.Y.; Chueh, S.H.; Lee, W.H.; Chen, A. Adriamycin impairs the contraction of mesangial cells through the inhibition of protein kinase C and intracellular calcium. Am. J. Physiol. Ren. Physiol. 2004, 287, F188–F194. [Google Scholar] [CrossRef] [PubMed]
- Kemp, M.; Donovan, J.; Higham, H.; Hooper, J. Biochemical markers of myocardial injury. Br. J. Anaesth. 2004, 93, 63–73. [Google Scholar] [CrossRef]
- Cao, Z.; Jia, Y.; Zhu, B. BNP and NT-proBNP as Diagnostic Biomarkers for Cardiac Dysfunction in Both Clinical and Forensic Medicine. Int. J. Mol. Sci. 2019, 20, 1820. [Google Scholar] [CrossRef]
- Sun, L.; Sun, Z.; Wang, Q.; Zhang, Y.; Jia, Z. Role of nuclear receptor PXR in immune cells and inflammatory diseases. Front. Immunol. 2022, 13, 969399. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Huang, L. Xenobiotic receptors in mediating the effect of sepsis on drug metabolism. Acta Pharm. Sin. B 2020, 10, 33–41. [Google Scholar] [CrossRef]
- Marx, C.E.; Bradford, D.W.; Hamer, R.M.; Naylor, J.C.; Allen, T.B.; Lieberman, J.A.; Strauss, J.L.; Kilts, J.D. Pregnenolone as a novel therapeutic candidate in schizophrenia: Emerging preclinical and clinical evidence. Neuroscience 2011, 191, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Bracci, M.; Zingaretti, L.; Martelli, M.; Lazzarini, R.; Salvio, G.; Amati, M.; Milinkovic, M.; Ulissi, A.; Medori, A.R.; Vitale, E.; et al. Alterations in Pregnenolone and Testosterone Levels in Male Shift Workers. Int. J. Environ. Res. Public Health 2023, 20, 3195. [Google Scholar] [CrossRef]
- Mannowetz, N.; Miller, M.R.; Lishko, P.V. Regulation of the sperm calcium channel CatSper by endogenous steroids and plant triterpenoids. Proc. Natl. Acad. Sci. USA 2017, 114, 5743–5748. [Google Scholar] [CrossRef]
- Fung, L.K.; Libove, R.A.; Phillips, J.; Haddad, F.; Hardan, A.Y. Brief report: An open-label study of the neurosteroid pregnenolone in adults with autism spectrum disorder. J. Autism Dev. Disord. 2014, 44, 2971–2977. [Google Scholar] [CrossRef]
- Shah, Y.M.; Ma, X.; Morimura, K.; Kim, I.; Gonzalez, F.J. Pregnane X receptor activation ameliorates DSS-induced inflammatory bowel disease via inhibition of NF-kappaB target gene expression. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G1114–G1122. [Google Scholar] [CrossRef]
- Okamura, M.; Shizu, R.; Abe, T.; Kodama, S.; Hosaka, T.; Sasaki, T.; Yoshinari, K. PXR Functionally Interacts with NF-κB and AP-1 to Downregulate the Inflammation-Induced Expression of Chemokine CXCL2 in Mice. Cells 2020, 9, 2296. [Google Scholar] [CrossRef] [PubMed]
- Lejri, I.; Grimm, A.; Hallé, F.; Abarghaz, M.; Klein, C.; Maitre, M.; Schmitt, M.; Bourguignon, J.-J.; Mensah-Nyagan, A.G.; Bihel, F. TSPO ligands boost mitochondrial function and pregnenolone synthesis. J. Alzheimer’s Dis. 2019, 72, 1045–1058. [Google Scholar] [CrossRef] [PubMed]
- Andrabi, S.S.; Kaushik, P.; Mumtaz, S.M.; Alam, M.M.; Tabassum, H.; Parvez, S. Pregnenolone Attenuates the Ischemia-Induced Neurological Deficit in the Transient Middle Cerebral Artery Occlusion Model of Rats. ACS Omega 2022, 7, 19122–19130. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, C.; Guo, H.; Chen, J.; Tao, Y.; Wang, F.; Lin, X.; Liu, Q.; Su, L.; Qin, A. Pregnenolone Inhibits Osteoclast Differentiation and Protects Against Lipopolysaccharide-Induced Inflammatory Bone Destruction and Ovariectomy-Induced Bone Loss. Front. Pharmacol. 2020, 11, 360. [Google Scholar] [CrossRef]
- Hayıroğlu, M.; Çınar, T.; Çiçek, V.; Asal, S.; Kılıç, Ş.; Keser, N.; Uzun, M.; Orhan, A.L. A simple formula to predict echocardiographic diastolic dysfunction-electrocardiographic diastolic index. Herz 2021, 46, 159–165. [Google Scholar] [CrossRef]
- Murugan, S.; Jakka, P.; Namani, S.; Mujumdar, V.; Radhakrishnan, G. The neurosteroid pregnenolone promotes degradation of key proteins in the innate immune signaling to suppress inflammation. J. Biol. Chem. 2019, 294, 4596–4607. [Google Scholar] [CrossRef]
- Ikewuchi, J.C.; Ikewuchi, C.C.; Ifeanacho, M.O.; Jaja, V.S.; Okezue, E.C.; Jamabo, C.N.; Adeku, K.A. Attenuation of doxorubicin-induced cardiotoxicity in Wistar rats by aqueous leaf-extracts of Chromolaena odorata and Tridax procumbens. J. Ethnopharmacol. 2021, 274, 114004. [Google Scholar] [CrossRef] [PubMed]
- Yuzhik, E.I.; Klinnikova, M.G.; Koldysheva, E.V.; Kapustina, V.I.; Lushnikova, E.L. Ultrastructural Myocardial Reorganization during Experimental Treatment with Doxorubicin and Atorvastatin. Bull. Exp. Biol. Med. 2022, 174, 109–115. [Google Scholar] [CrossRef]
- Lewis, R.J.; Sax, N.I. Sax’s Dangerous Properties of Industrial Materials, 11th ed.; Lewis, R.J., Sr., Ed.; Wiley-Interscience: Hoboken, NJ, USA, 2004; 88p. [Google Scholar]
- Alkhanjaf, A.A.M.; Athar, M.T.; Ullah, Z.; Alsayhab, A.M.H.; Umar, A.; Shaikh, I.A. Farnesol Protects against Cardiotoxicity Caused by Doxorubicin-Induced Stress, Inflammation, and Cell Death: An In Vivo Study in Wistar Rats. Molecules 2022, 27, 8589. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Jin, D.; Qi, J.; Wang, X.; Zhang, C.; An, P.; Luo, Y.; Luo, J. Fucoidan Protects against Doxorubicin-Induced Cardiotoxicity by Reducing Oxidative Stress and Preventing Mitochondrial Function Injury. Int. J. Mol. Sci. 2022, 23, 10685. [Google Scholar] [CrossRef]
- Al-Hadi, H.A.; Fox, K.A. Cardiac markers in the early diagnosis and management of patients with acute coronary syndrome. Sultan Qaboos Univ. Med. J. 2009, 9, 231–246. [Google Scholar] [PubMed]
- Mukhopadhyay, P.; Rajesh, M.; Bátkai, S.; Kashiwaya, Y.; Haskó, G.; Liaudet, L.; Szabó, C.; Pacher, P. Role of superoxide, nitric oxide, and peroxynitrite in doxorubicin-induced cell death in vivo and in vitro. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H1466–H1483. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Wang, X.; Xiong, L.L.; Zhu, Y.; Chen, H.L.; Chen, J.X.; Wang, X.X.; Li, R.L.; Guo, Z.Y.; Li, P.; et al. Salidroside improves doxorubicin-induced cardiac dysfunction by suppression of excessive oxidative stress and cardiomyocyte apoptosis. J. Cardiovasc. Pharmacol. 2013, 62, 512–523. [Google Scholar] [CrossRef]
- Sirangelo, I.; Liccardo, M.; Iannuzzi, C. Hydroxytyrosol Prevents Doxorubicin-Induced Oxidative Stress and Apoptosis in Cardiomyocytes. Antioxidants 2022, 11, 1087. [Google Scholar] [CrossRef] [PubMed]
- Cheah, I.K.; Tang, R.M.Y.; Wang, X.; Sachaphibulkij, K.; Chong, S.Y.; Lim, L.H.K.; Wang, J.-W.; Halliwell, B. Protection against Doxorubicin-Induced Cardiotoxicity by Ergothioneine. Antioxidants 2023, 12, 320. [Google Scholar] [CrossRef] [PubMed]
- Koss-Mikołajczyk, I.; Todorovic, V.; Sobajic, S.; Mahajna, J.; Gerić, M.; Tur, J.A.; Bartoszek, A. Natural Products Counteracting Cardiotoxicity during Cancer Chemotherapy: The Special Case of Doxorubicin, a Comprehensive Review. Int. J. Mol. Sci. 2021, 22, 10037. [Google Scholar] [CrossRef]
- Wang, J.; Yao, L.; Wu, X.; Guo, Q.; Sun, S.; Li, J.; Shi, G.; Caldwell, R.B.; Caldwell, R.W.; Chen, Y. Protection against Doxorubicin-Induced Cardiotoxicity through Modulating iNOS/ARG 2 Balance by Electroacupuncture at PC6. Oxidative Med. Cell. Longev. 2021, 2021, 6628957. [Google Scholar] [CrossRef] [PubMed]
- Elbatreek, M.H.; Mucke, H.; Schmidt, H. NOX Inhibitors: From Bench to Naxibs to Bedside. In Reactive Oxygen Species; Handbook of Experimental Pharmacology; Springer: Cham, Switzerland, 2021; Volume 264, pp. 145–168. [Google Scholar] [CrossRef]
- Zeng, C.; Duan, F.; Hu, J.; Luo, B.; Huang, B.; Lou, X.; Sun, X.; Li, H.; Zhang, X.; Yin, S.; et al. NLRP3 inflammasome-mediated pyroptosis contributes to the pathogenesis of non-ischemic dilated cardiomyopathy. Redox Biol. 2020, 34, 101523. [Google Scholar] [CrossRef]
- Zheng, H.; Xu, N.; Zhang, Z.; Wang, F.; Xiao, J.; Ji, X. Setanaxib (GKT137831) Ameliorates Doxorubicin-Induced Cardiotoxicity by Inhibiting the NOX1/NOX4/Reactive Oxygen Species/MAPK Pathway. Front. Pharmacol. 2022, 13, 823975. [Google Scholar] [CrossRef]
- Iwata, K.; Matsuno, K.; Murata, A.; Zhu, K.; Fukui, H.; Ikuta, K.; Katsuyama, M.; Ibi, M.; Matsumoto, M.; Ohigashi, M.; et al. Up-regulation of NOX1/NADPH oxidase following drug-induced myocardial injury promotes cardiac dysfunction and fibrosis. Free. Radic. Biol. Med. 2018, 120, 277–288. [Google Scholar] [CrossRef]
- Kandasamy, A.D.; Chow, A.K.; Ali, M.A.; Schulz, R. Matrix metalloproteinase-2 and myocardial oxidative stress injury: Beyond the matrix. Cardiovasc. Res. 2010, 85, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, P.R.; Nascimento, L.D.; Gerlach, R.F.; Rodrigues, K.E.; Prado, A.F. Matrix Metalloproteinase 2 as a Pharmacological Target in Heart Failure. Pharmaceuticals 2022, 15, 920. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.Y.H.; Roczkowsky, A.; Cho, W.J.; Poirier, M.; Sergi, C.; Keschrumrus, V.; Churko, J.M.; Granzier, H.; Schulz, R. MMP inhibitors attenuate doxorubicin cardiotoxicity by preventing intracellular and extracellular matrix remodelling. Cardiovasc. Res. 2021, 117, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Akolkar, G.; da Silva Dias, D.; Ayyappan, P.; Bagchi, A.K.; Jassal, D.S.; Salemi, V.M.C.; Irigoyen, M.C.; De Angelis, K.; Singal, P.K. Vitamin C mitigates oxidative/nitrosative stress and inflammation in doxorubicin-induced cardiomyopathy. Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H795–H809. [Google Scholar] [CrossRef]
- Hafez, H.M.; Hassanein, H. Montelukast ameliorates doxorubicin-induced cardiotoxicity via modulation of p-glycoprotein and inhibition of ROS-mediated TNF-α/NF-κB pathways. Drug Chem. Toxicol. 2022, 45, 548–559. [Google Scholar] [CrossRef]
- Christidi, E.; Brunham, L.R. Regulated cell death pathways in doxorubicin-induced cardiotoxicity. Cell Death Dis. 2021, 12, 339. [Google Scholar] [CrossRef]
- Leskiewicz, M.; Jantas, D.; Budziszewska, B.; Lason, W. Excitatory neurosteroids attenuate apoptotic and excitotoxic cell death in primary cortical neurons. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2008, 59, 457–475. [Google Scholar]
- Xiao, X.; Chen, L.; Ouyang, Y.; Zhu, W.; Qiu, P.; Su, X.; Dou, Y.; Tang, L.; Yan, M.; Zhang, H.; et al. Pregnenolone, a cholesterol metabolite, induces glioma cell apoptosis via activating extrinsic and intrinsic apoptotic pathways. Oncol. Lett. 2014, 8, 645–650. [Google Scholar] [CrossRef]
- Brown, E.S.; Park, J.; Marx, C.E.; Hynan, L.S.; Gardner, C.; Davila, D.; Nakamura, A.; Sunderajan, P.; Lo, A.; Holmes, T. A randomized, double-blind, placebo-controlled trial of pregnenolone for bipolar depression. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2014, 39, 2867–2873. [Google Scholar] [CrossRef]
- Podyacheva, E.Y.; Kushnareva, E.A.; Karpov, A.A.; Toropova, Y.G. Analysis of Models of Doxorubicin-Induced Cardiomyopathy in Rats and Mice. A Modern View from the Perspective of the Pathophysiologist and the Clinician. Front. Pharmacol. 2021, 12, 670479. [Google Scholar] [CrossRef]
- Lowes, S.; Haslam, I.S.; Fihn, B.M.; Hilgendorf, C.; Karlsson, J.E.; Simmons, N.L.; Ungell, A.L. The Effects of Pregnenolone 16α-Carbonitrile Dosing on Digoxin Pharmacokinetics and Intestinal Absorption in the Rat. Pharmaceutics 2010, 2, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.; Morsy, M.A.; Jacob, S. Dose translation between laboratory animals and human in preclinical and clinical phases of drug development. Drug Dev. Res. 2018, 79, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Refaie, M.M.M.; Abdel-Gaber, S.A.; Rahman, S.; Hafez, S.; Khalaf, H.M. Cardioprotective effects of bosentan in 5-fluorouracil-induced cardiotoxicity. Toxicology 2022, 465, 153042. [Google Scholar] [CrossRef] [PubMed]
- Senousy, S.R.; El-Daly, M.; Ibrahim, A.R.N.; Khalifa, M.M.A.; Ahmed, A.-S.F. Effect of Celecoxib and Infliximab against Multiple Organ Damage Induced by Sepsis in Rats: A Comparative Study. Biomedicines 2022, 10, 1613. [Google Scholar] [CrossRef]
- Matouk, A.I.; El-Daly, M.; Habib, H.A.; Senousy, S.; Naguib Abdel Hafez, S.M.; Kasem, A.W.; Almalki, W.H.; Alzahrani, A.; Alshehri, A.; Ahmed, A.F. Protective effects of menthol against sepsis-induced hepatic injury: Role of mediators of hepatic inflammation, apoptosis, and regeneration. Front. Pharmacol. 2022, 13, 952337. [Google Scholar] [CrossRef]
- Morsy, M.A.; Gupta, S.; Nair, A.B.; Venugopala, K.N.; Greish, K.; El-Daly, M. Protective Effect of Spirulina platensis Extract against Dextran-Sulfate-Sodium-Induced Ulcerative Colitis in Rats. Nutrients 2019, 11, 2309. [Google Scholar] [CrossRef]
- Suvarna, K.S.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques; E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Morsy, M.A.; El-Sheikh, A.A.K.; Abdel-Hafez, S.M.N.; Kandeel, M.; Abdel-Gaber, S.A. Paeonol Protects Against Methotrexate-Induced Nephrotoxicity via Upregulation of P-gp Expression and Inhibition of TLR4/NF-κB Pathway. Front. Pharmacol. 2022, 13, 774387. [Google Scholar] [CrossRef]
- Landmann, M.; Scheibner, D.; Graaf, A.; Gischke, M.; Koethe, S.; Fatola, O.I.; Raddatz, B.; Mettenleiter, T.C.; Beer, M.; Grund, C.; et al. A Semiquantitative Scoring System for Histopathological and Immunohistochemical Assessment of Lesions and Tissue Tropism in Avian Influenza. Viruses 2021, 13, 868. [Google Scholar] [CrossRef]
- Wen, Y.; Liu, R.; Lin, N.; Luo, H.; Tang, J.; Huang, Q.; Sun, H.; Tang, L. NADPH Oxidase Hyperactivity Contributes to Cardiac Dysfunction and Apoptosis in Rats with Severe Experimental Pancreatitis through ROS-Mediated MAPK Signaling Pathway. Oxidative Med. Cell. Longev. 2019, 2019, 4578175. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Histological Changes | Control | Preg | DOX | Preg + DOX |
|---|---|---|---|---|
| Distorted architecture of cardiac muscles | − | − | +++ | + |
| Loss of muscular striations | − | − | ++ | + |
| Cells with pyknotic nuclei | − | − | +++ | + |
| Vascular congestion | − | − | +++ | + |
| Hemorrhage | − | − | ++ | - |
| Inflammatory cellular infiltrate | − | − | ++ | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morsy, M.A.; Abdel-Gaber, S.A.; Mokhemer, S.A.; Kandeel, M.; Sedik, W.F.; Nair, A.B.; Venugopala, K.N.; Khalil, H.E.; Al-Dhubiab, B.E.; Mohamed, M.Z. Pregnenolone Inhibits Doxorubicin-Induced Cardiac Oxidative Stress, Inflammation, and Apoptosis—Role of Matrix Metalloproteinase 2 and NADPH Oxidase 1. Pharmaceuticals 2023, 16, 665. https://doi.org/10.3390/ph16050665
Morsy MA, Abdel-Gaber SA, Mokhemer SA, Kandeel M, Sedik WF, Nair AB, Venugopala KN, Khalil HE, Al-Dhubiab BE, Mohamed MZ. Pregnenolone Inhibits Doxorubicin-Induced Cardiac Oxidative Stress, Inflammation, and Apoptosis—Role of Matrix Metalloproteinase 2 and NADPH Oxidase 1. Pharmaceuticals. 2023; 16(5):665. https://doi.org/10.3390/ph16050665
Chicago/Turabian StyleMorsy, Mohamed A., Seham A. Abdel-Gaber, Sahar A. Mokhemer, Mahmoud Kandeel, Wael F. Sedik, Anroop B. Nair, Katharigatta N. Venugopala, Hany Ezzat Khalil, Bandar E. Al-Dhubiab, and Mervat Z. Mohamed. 2023. "Pregnenolone Inhibits Doxorubicin-Induced Cardiac Oxidative Stress, Inflammation, and Apoptosis—Role of Matrix Metalloproteinase 2 and NADPH Oxidase 1" Pharmaceuticals 16, no. 5: 665. https://doi.org/10.3390/ph16050665