
Study of lncRNAs in Pediatric Neurological Diseases: Methods, Analysis of the State-of-Art and Possible Therapeutic Implications

 , , and
, , and 
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Long Non-Coding RNAs
1.2. Methodological Approaches for the Study of lncRNA Expression and Function
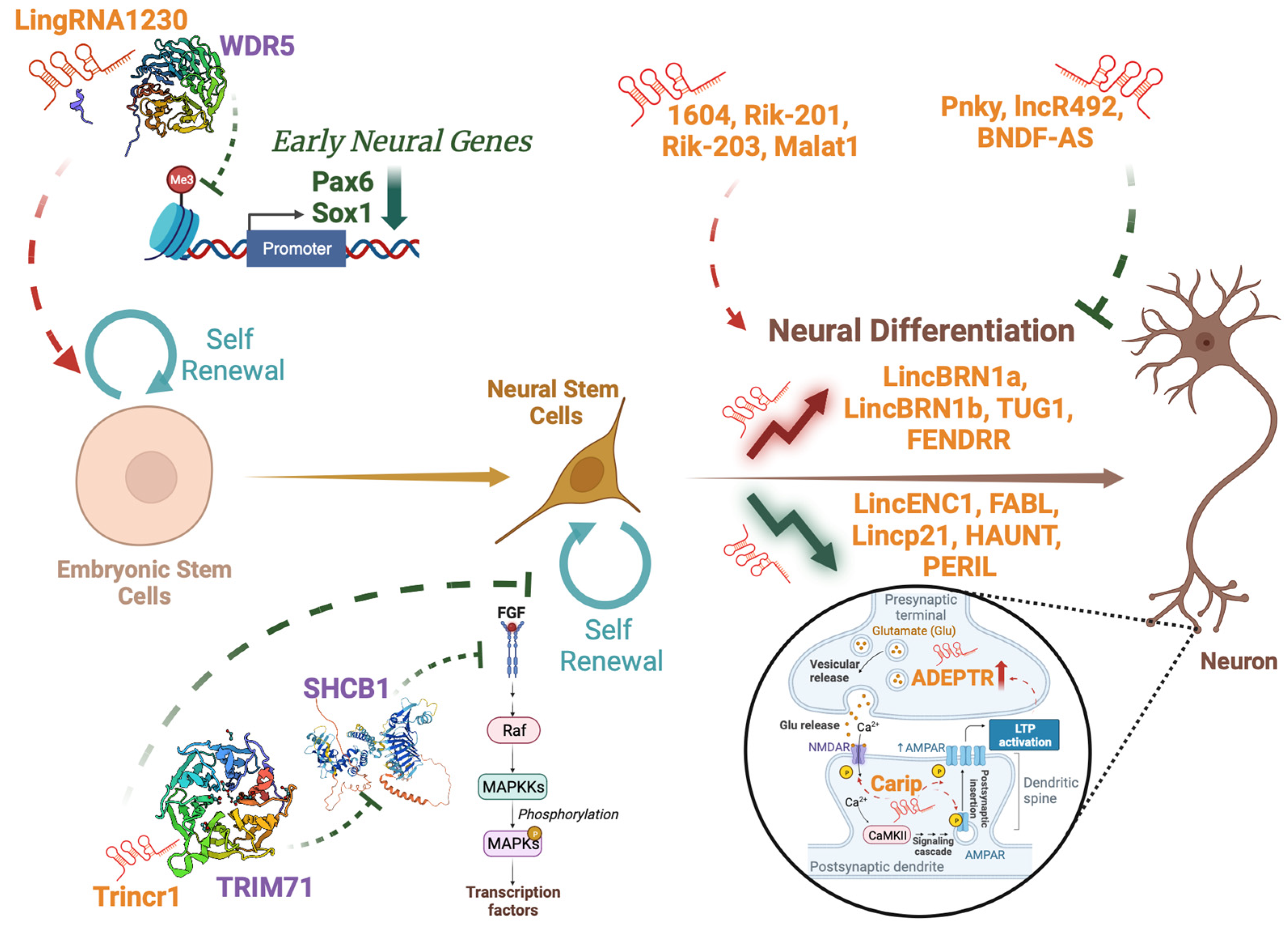
1.3. Role of LncRNAS in Neural Development
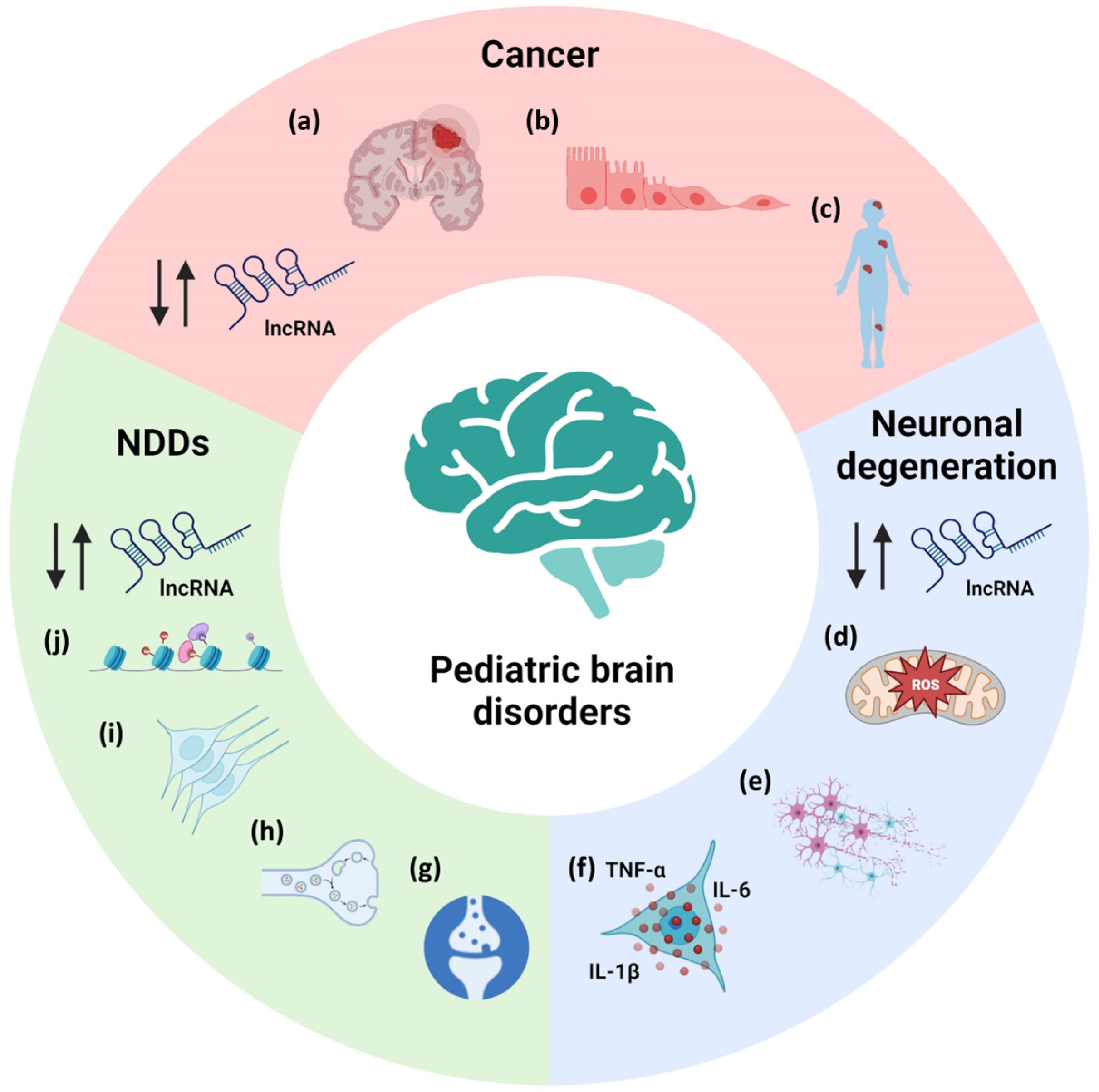
2. LncRNAs in Pediatric Neurological Diseases
2.1. LncRNAs in Pediatric Neurodevelopmental Disorders
2.2. LncRNAs in Pediatric Neurodegenerative Diseases
2.3. LncRNAs in Pediatric Brain Tumors
3. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene Regulation by Long Non-Coding RNAs and Its Biological Functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.-L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long Non-Coding RNAs: Definitions, Functions, Challenges and Recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.C.R.; Acuña, S.M.; Aoki, J.I.; Floeter-Winter, L.M.; Muxel, S.M. Long Non-Coding RNAs in the Regulation of Gene Expression: Physiology and Disease. Non-Coding RNA 2019, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Profumo, V.; Forte, B.; Percio, S.; Rotundo, F.; Doldi, V.; Ferrari, E.; Fenderico, N.; Dugo, M.; Romagnoli, D.; Benelli, M.; et al. LEADeR Role of miR-205 Host Gene as Long Noncoding RNA in Prostate Basal Cell Differentiation. Nat. Commun. 2019, 10, 307. [Google Scholar] [CrossRef]
- Tassinari, M.; Richter, S.N.; Gandellini, P. Biological Relevance and Therapeutic Potential of G-Quadruplex Structures in the Human Noncoding Transcriptome. Nucleic Acids Res. 2021, 49, 3617–3633. [Google Scholar] [CrossRef]
- Pagani, G.; Pandini, C.; Gandellini, P. Navigating the Multiverse of Antisense RNAs: The Transcription- and RNA-Dependent Dimension. Non-Coding RNA 2022, 8, 74. [Google Scholar] [CrossRef]
- Pandini, C.; Garofalo, M.; Rey, F.; Garau, J.; Zucca, S.; Sproviero, D.; Bordoni, M.; Berzero, G.; Davin, A.; Poloni, T.E.; et al. MINCR: A Long Non-Coding RNA Shared between Cancer and Neurodegeneration. Genomics 2021, 113, 4039–4051. [Google Scholar] [CrossRef]
- Rey, F.; Pandini, C.; Messa, L.; Launi, R.; Barzaghini, B.; Zangaglia, R.; Raimondi, M.T.; Gagliardi, S.; Cereda, C.; Zuccotti, G.V.; et al. α-Synuclein Antisense Transcript SNCA-AS1 Regulates Synapses- and Aging-Related Genes Suggesting Its Implication in Parkinson’s Disease. Aging Cell 2021, 20, e13504. [Google Scholar] [CrossRef]
- Rey, F.; Maghraby, E.; Messa, L.; Esposito, L.; Barzaghini, B.; Pandini, C.; Bordoni, M.; Gagliardi, S.; Diamanti, L.; Raimondi, M.T.; et al. Identification of a Novel Pathway in Sporadic Amyotrophic Lateral Sclerosis Mediated by the Long Non-Coding RNA ZEB1-AS1. Neurobiol. Dis. 2023, 178, 106030. [Google Scholar] [CrossRef]
- Wang, H.-L.V.; Chekanova, J.A. An Overview of Methodologies in Studying lncRNAs in the High-Throughput Era: When Acronyms ATTACK! Methods Mol. Biol. 2019, 1933, 1–30. [Google Scholar] [CrossRef]
- Core, L.J.; Waterfall, J.J.; Lis, J.T. Nascent RNA Sequencing Reveals Widespread Pausing and Divergent Initiation at Human Promoters. Science 2008, 322, 1845–1848. [Google Scholar] [CrossRef] [PubMed]
- Churchman, L.S.; Weissman, J.S. Nascent Transcript Sequencing Visualizes Transcription at Nucleotide Resolution. Nature 2011, 469, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome Engineering Using the CRISPR-Cas9 System. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.; Quinn, J.; Chang, H.Y. Chromatin Isolation by RNA Purification (ChIRP). J. Vis. Exp. JoVE 2012, 61, e3912. [Google Scholar] [CrossRef]
- Engreitz, J.; Lander, E.S.; Guttman, M. RNA Antisense Purification (RAP) for Mapping RNA Interactions with Chromatin. Methods Mol. Biol. 2015, 1262, 183–197. [Google Scholar] [CrossRef]
- Simon, M.D. Capture Hybridization Analysis of RNA Targets (CHART). Curr. Protoc. Mol. Biol. 2013, 101, 21–25. [Google Scholar] [CrossRef]
- Gagliardi, M.; Matarazzo, M.R. RIP: RNA Immunoprecipitation. Methods Mol. Biol. 2016, 1480, 73–86. [Google Scholar] [CrossRef]
- Marín-Béjar, O.; Huarte, M. RNA Pulldown Protocol for in Vitro Detection and Identification of RNA-Associated Proteins. Methods Mol. Biol. 2015, 1206, 87–95. [Google Scholar] [CrossRef]
- Stiles, J.; Jernigan, T.L. The Basics of Brain Development. Neuropsychol. Rev. 2010, 20, 327–348. [Google Scholar] [CrossRef]
- Vijayraghavan, D.S.; Davidson, L.A. Mechanics of Neurulation: From Classical to Current Perspectives on the Physical Mechanics That Shape, Fold, and Form the Neural Tube. Birth Defects Res. 2017, 109, 153–168. [Google Scholar] [CrossRef]
- Butler, S.J.; Bronner, M.E. From Classical to Current: Analyzing Peripheral Nervous System and Spinal Cord Lineage and Fate. Dev. Biol. 2015, 398, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Yuasa-Kawada, J.; Kinoshita-Kawada, M.; Tsuboi, Y.; Wu, J.Y. Neuronal Guidance Genes in Health and Diseases. Protein Cell 2023, 14, 238–261. [Google Scholar] [CrossRef] [PubMed]
- Qi, C.; Luo, L.-D.; Feng, I.; Ma, S. Molecular Mechanisms of Synaptogenesis. Front. Synaptic Neurosci. 2022, 14, 939793. [Google Scholar] [CrossRef] [PubMed]
- Salinas, R.D.; Connolly, D.R.; Song, H. Invited Review: Epigenetics in Neurodevelopment. Neuropathol. Appl. Neurobiol. 2020, 46, 6–27. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, H.; Zhang, Q.; Zhang, Y. The Functions of Long Non-Coding RNAs in Neural Stem Cell Proliferation and Differentiation. Cell Biosci. 2020, 10, 74. [Google Scholar] [CrossRef]
- Sauvageau, M.; Goff, L.A.; Lodato, S.; Bonev, B.; Groff, A.F.; Gerhardinger, C.; Sanchez-Gomez, D.B.; Hacisuleyman, E.; Li, E.; Spence, M.; et al. Multiple Knockout Mouse Models Reveal lincRNAs Are Required for Life and Brain Development. eLife 2013, 2, e01749. [Google Scholar] [CrossRef]
- Goff, L.A.; Groff, A.F.; Sauvageau, M.; Trayes-Gibson, Z.; Sanchez-Gomez, D.B.; Morse, M.; Martin, R.D.; Elcavage, L.E.; Liapis, S.C.; Gonzalez-Celeiro, M.; et al. Spatiotemporal Expression and Transcriptional Perturbations by Long Noncoding RNAs in the Mouse Brain. Proc. Natl. Acad. Sci. USA 2015, 112, 6855–6862. [Google Scholar] [CrossRef] [PubMed]
- Carelli, S.; Giallongo, T.; Rey, F.; Latorre, E.; Bordoni, M.; Mazzucchelli, S.; Gorio, M.C.; Pansarasa, O.; Provenzani, A.; Cereda, C.; et al. HuR Interacts with lincBRN1a and lincBRN1b during Neuronal Stem Cells Differentiation. RNA Biol. 2019, 16, 1471–1485. [Google Scholar] [CrossRef]
- Wang, C.; Li, G.; Wu, Y.; Xi, J.; Kang, J. LincRNA1230 Inhibits the Differentiation of Mouse ES Cells towards Neural Progenitors. Sci. China Life Sci. 2016, 59, 443–454. [Google Scholar] [CrossRef]
- Lin, Y.; Cai, H. Biological Functions and Therapeutic Potential of SHCBP1 in Human Cancer. Biomed. Pharmacother. 2023, 160, 114362. [Google Scholar] [CrossRef]
- Li, Y.-P.; Duan, F.-F.; Zhao, Y.-T.; Gu, K.-L.; Liao, L.-Q.; Su, H.-B.; Hao, J.; Zhang, K.; Yang, N.; Wang, Y. A TRIM71 Binding Long Noncoding RNA Trincr1 Represses FGF/ERK Signaling in Embryonic Stem Cells. Nat. Commun. 2019, 10, 1368. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.-Y.; Bogu, G.K.; Soh, B.S.; Stanton, L.W. The Long Noncoding RNA RMST Interacts with SOX2 to Regulate Neurogenesis. Mol. Cell 2013, 51, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Shen, W.; Zhang, B.; Tian, K.; Li, Y.; Mu, L.; Luo, Z.; Zhong, X.; Wu, X.; Liu, Y.; et al. Long Non-Coding RNA LncKdm2b Regulates Cortical Neuronal Differentiation by Cis-Activating Kdm2b. Protein Cell 2020, 11, 161–186. [Google Scholar] [CrossRef] [PubMed]
- Vance, K.W.; Sansom, S.N.; Lee, S.; Chalei, V.; Kong, L.; Cooper, S.E.; Oliver, P.L.; Ponting, C.P. The Long Non-Coding RNA Paupar Regulates the Expression of Both Local and Distal Genes. EMBO J. 2014, 33, 296–311. [Google Scholar] [CrossRef]
- Chen, L.; Feng, P.; Zhu, X.; He, S.; Duan, J.; Zhou, D. Long Non-coding RNA Malat1 Promotes Neurite Outgrowth through Activation of ERK/MAPK Signalling Pathway in N2a Cells. J. Cell. Mol. Med. 2016, 20, 2102–2110. [Google Scholar] [CrossRef]
- Weng, R.; Lu, C.; Liu, X.; Li, G.; Lan, Y.; Qiao, J.; Bai, M.; Wang, Z.; Guo, X.; Ye, D.; et al. Long Noncoding RNA-1604 Orchestrates Neural Differentiation through the miR-200c/ZEB Axis. Stem Cells 2018, 36, 325–336. [Google Scholar] [CrossRef]
- Zhang, L.; Xue, Z.; Yan, J.; Wang, J.; Liu, Q.; Jiang, H. LncRNA Riken-201 and Riken-203 Modulates Neural Development by Regulating the Sox6 through Sequestering miRNAs. Cell Prolif. 2019, 52, e12573. [Google Scholar] [CrossRef]
- Grammatikakis, I.; Gorospe, M. Identification of Neural Stem Cell Differentiation Repressor Complex Pnky-PTBP1. Stem Cell Investig. 2016, 3, 10. [Google Scholar] [CrossRef]
- Winzi, M.; Casas Vila, N.; Paszkowski-Rogacz, M.; Ding, L.; Noack, S.; Theis, M.; Butter, F.; Buchholz, F. The Long Noncoding RNA lncR492 Inhibits Neural Differentiation of Murine Embryonic Stem Cells. PLoS ONE 2018, 13, e0191682. [Google Scholar] [CrossRef]
- Zheng, X.; Lin, C.; Li, Y.; Ye, J.; Zhou, J.; Guo, P. Long Noncoding RNA BDNF-AS Regulates Ketamine-Induced Neurotoxicity in Neural Stem Cell Derived Neurons. Biomed. Pharmacother. 2016, 82, 722–728. [Google Scholar] [CrossRef]
- Rey, F.; Pandini, C.; Barzaghini, B.; Messa, L.; Giallongo, T.; Pansarasa, O.; Gagliardi, S.; Brilli, M.; Zuccotti, G.V.; Cereda, C.; et al. Dissecting the Effect of a 3D Microscaffold on the Transcriptome of Neural Stem Cells with Computational Approaches: A Focus on Mechanotransduction. Int. J. Mol. Sci. 2020, 21, 6775. [Google Scholar] [CrossRef] [PubMed]
- Messa, L.; Barzaghini, B.; Rey, F.; Pandini, C.; Zuccotti, G.V.; Cereda, C.; Carelli, S.; Raimondi, M.T. Neural Precursor Cells Expanded Inside the 3D Micro-Scaffold Nichoid Present Different Non-Coding RNAs Profiles and Transcript Isoforms Expression: Possible Epigenetic Modulation by 3D Growth. Biomedicines 2021, 9, 1120. [Google Scholar] [CrossRef] [PubMed]
- Carelli, S.; Giallongo, T.; Rey, F.; Barzaghini, B.; Zandrini, T.; Pulcinelli, A.; Nardomarino, R.; Cerullo, G.; Osellame, R.; Cereda, C.; et al. Neural Precursors Cells Expanded in a 3D Micro-Engineered Niche Present Enhanced Therapeutic Efficacy In Vivo. Nanotheranostics 2021, 5, 8–26. [Google Scholar] [CrossRef]
- Wang, F.; Wang, Q.; Liu, B.; Mei, L.; Ma, S.; Wang, S.; Wang, R.; Zhang, Y.; Niu, C.; Xiong, Z.; et al. The Long Noncoding RNA Synage Regulates Synapse Stability and Neuronal Function in the Cerebellum. Cell Death Differ. 2021, 28, 2634–2650. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Zhang, R.; Yang, Y.; Wu, E.; Tang, Y.; Zhao, Z.; Li, C.; Yang, L.; Teng, X.; Ye, Y.; et al. Identification and Characterization of Long Non-Coding RNA Carip in Modulating Spatial Learning and Memory. Cell Rep. 2022, 38, 110398. [Google Scholar] [CrossRef]
- Grinman, E.; Nakahata, Y.; Avchalumov, Y.; Espadas, I.; Swarnkar, S.; Yasuda, R.; Puthanveettil, S.V. Activity-Regulated Synaptic Targeting of lncRNA ADEPTR Mediates Structural Plasticity by Localizing Sptn1 and AnkB in Dendrites. Sci. Adv. 2021, 7, eabf0605. [Google Scholar] [CrossRef]
- Wei, A.; Wang, L. Prediction of Synaptically Localized RNAs in Human Neurons Using Developmental Brain Gene Expression Data. Genes 2022, 13, 1488. [Google Scholar] [CrossRef]
- Thapar, A.; Cooper, M.; Rutter, M. Neurodevelopmental Disorders. Lancet Psychiatry 2017, 4, 339–346. [Google Scholar] [CrossRef]
- First, M.B. Diagnostic and Statistical Manual of Mental Disorders, 5th Edition, and Clinical Utility. J. Nerv. Ment. Dis. 2013, 201, 727. [Google Scholar] [CrossRef]
- Parenti, I.; Rabaneda, L.G.; Schoen, H.; Novarino, G. Neurodevelopmental Disorders: From Genetics to Functional Pathways. Trends Neurosci 2020, 43, 608–621. [Google Scholar] [CrossRef]
- Zhang, S.F.; Gao, J.; Liu, C.M. The Role of Non-Coding RNAs in Neurodevelopmental Disorders. Front. Genet. 2019, 10, 1033. [Google Scholar] [CrossRef]
- Ziats, M.N.; Rennert, O.M. Aberrant Expression of Long Noncoding RNAs in Autistic Brain. J. Mol. Neurosci. 2013, 49, 589–593. [Google Scholar] [CrossRef]
- Luo, T.; Liu, P.; Wang, X.Y.; Li, L.Z.; Zhao, L.P.; Huang, J.; Li, Y.M.; Ou, J.L.; Peng, X.Q. Effect of the Autism-Associated lncRNA Shank2-AS on Architecture and Growth of Neurons. J. Cell Biochem. 2018, 120, 1754–1762. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, X.; Ju, W.; Flory, M.; Zhong, J.; Jiang, S.; Wang, P.; Dong, X.; Tao, X.; Chen, Q.; et al. Genome-Wide Differential Expression of Synaptic Long Noncoding RNAs in Autism Spectrum Disorder. Transl. Psychiatry 2015, 5, e660. [Google Scholar] [CrossRef] [PubMed]
- Kerin, T.; Ramanathan, A.; Rivas, K.; Grepo, N.; Coetzee, G.A.; Campbell, D.B. A Noncoding RNA Antisense to Moesin at 5p14.1 in Autism. Sci. Transl. Med. 2012, 4, 128ra40. [Google Scholar] [CrossRef] [PubMed]
- DeWitt, J.J.; Hecht, P.M.; Grepo, N.; Wilkinson, B.; Evgrafov, O.V.; Morris, K.V.; Knowles, J.A.; Campbell, D.B. Transcriptional Gene Silencing of the Autism-Associated Long Noncoding RNA MSNP1AS in Human Neural Progenitor Cells. Dev. Neurosci. 2016, 38, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Verkerk, A.J.M.H.; Pieretti, M.; Sutcliffe, J.S.; Fu, Y.-H.; Kuhl, D.P.A.; Pizzuti, A.; Reiner, O.; Richards, S.; Victoria, M.F.; Zhang, F.; et al. Identification of a Gene (FMR-1) Containing a CGG Repeat Coincident with a Breakpoint Cluster Region Exhibiting Length Variation in Fragile X Syndrome. Cell 1991, 65, 905–914. [Google Scholar] [CrossRef]
- Ladd, P.D.; Smith, L.E.; Rabaia, N.A.; Moore, J.M.; Georges, S.A.; Hansen, R.S.; Hagerman, R.J.; Tassone, F.; Tapscott, S.J.; Filippova, G.N. An Antisense Transcript Spanning the CGG Repeat Region of FMR1 Is Upregulated in Premutation Carriers but Silenced in Full Mutation Individuals. Hum. Mol. Genet. 2007, 16, 3174–3187. [Google Scholar] [CrossRef]
- Pastori, C.; Peschansky, V.J.; Barbouth, D.; Mehta, A.; Silva, J.P.; Wahlestedt, C. Comprehensive Analysis of the Transcriptional Landscape of the Human FMR1 Gene Reveals Two New Long Noncoding RNAs Differentially Expressed in Fragile X Syndrome and Fragile X-Associated Tremor/Ataxia Syndrome. Hum. Genet. 2014, 133, 59–67. [Google Scholar] [CrossRef]
- Petazzi, P.; Sandoval, J.; Szczesna, K.; Jorge, O.C.; Roa, L.; Sayols, S.; Gomez, A.; Huertas, D.; Esteller, M. Dysregulation of the Long Non-Coding RNA Transcriptome in a Rett Syndrome Mouse Model. RNA Biol. 2013, 10, 1197–1203. [Google Scholar] [CrossRef]
- Meng, L.; Person, R.E.; Beaudet, A.L. Ube3a-ATS Is an Atypical RNA Polymerase II Transcript That Represses the Paternal Expression of Ube3a. Hum. Mol. Genet. 2012, 21, 3001–3012. [Google Scholar] [CrossRef] [PubMed]
- Dyken, P.; Krawiecki, N. Neurodegenerative Diseases of Infancy and Childhood. Ann. Neurol. 1983, 13, 351–364. [Google Scholar] [CrossRef] [PubMed]
- Mastrangelo, M. Clinical Approach to Neurodegenerative Disorders in Childhood: An Updated Overview. Acta Neurol. Belg. 2019, 119, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Nittari, G.; Tomassoni, D.; Roy, P.; Martinelli, I.; Tayebati, S.K.; Amenta, F. Batten Disease through Different in Vivo and in Vitro Models: A Review. J. Neurosci. Res. 2023, 101, 298–315. [Google Scholar] [CrossRef]
- Anderton, R.S.; Meloni, B.P.; Mastaglia, F.L.; Boulos, S. Spinal Muscular Atrophy and the Antiapoptotic Role of Survival of Motor Neuron (SMN) Protein. Mol. Neurobiol. 2013, 47, 821–832. [Google Scholar] [CrossRef]
- Van Rappard, D.F.; Boelens, J.J.; Wolf, N.I. Metachromatic Leukodystrophy: Disease Spectrum and Approaches for Treatment. Best. Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 261–273. [Google Scholar] [CrossRef]
- Vanier, M.T. Niemann–Pick Diseases. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2013; Volume 113, pp. 1717–1721. ISBN 978-0-444-59565-2. [Google Scholar]
- Chen, K.-W.; Chen, J.-A. Functional Roles of Long Non-Coding RNAs in Motor Neuron Development and Disease. J. Biomed. Sci. 2020, 27, 38. [Google Scholar] [CrossRef]
- Han, S.; Ren, M.; Kuang, T.; Pang, M.; Guan, D.; Liu, Y.; Wang, Y.; Zhang, W.; Ye, Z. Cerebellar Long Noncoding RNA Expression Profile in a Niemann-Pick C Disease Mouse Model. Mol. Neurobiol. 2021, 58, 5826–5836. [Google Scholar] [CrossRef]
- Lunn, M.R.; Wang, C.H. Spinal Muscular Atrophy. The Lancet 2008, 371, 2120–2133. [Google Scholar] [CrossRef]
- Woo, C.J.; Maier, V.K.; Davey, R.; Brennan, J.; Li, G.; Brothers, J.; Schwartz, B.; Gordo, S.; Kasper, A.; Okamoto, T.R.; et al. Gene Activation of SMN by Selective Disruption of lncRNA-Mediated Recruitment of PRC2 for the Treatment of Spinal Muscular Atrophy. Proc. Natl. Acad. Sci. USA 2017, 114, E1509–E1518. [Google Scholar] [CrossRef]
- Bernabò, P.; Tebaldi, T.; Groen, E.J.N.; Lane, F.M.; Perenthaler, E.; Mattedi, F.; Newbery, H.J.; Zhou, H.; Zuccotti, P.; Potrich, V.; et al. In Vivo Translatome Profiling in Spinal Muscular Atrophy Reveals a Role for SMN Protein in Ribosome Biology. Cell Rep. 2017, 21, 953–965. [Google Scholar] [CrossRef] [PubMed]
- Vangoor, V.R.; Gomes-Duarte, A.; Pasterkamp, R.J. Long Non-coding RNAs in Motor Neuron Development and Disease. J. Neurochem. 2021, 156, 777–801. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Abak, A.; Khademi, S.; Shoorei, H.; Bahroudi, Z.; Taheri, M.; Akbari Dilmaghani, N. Functional Roles of Non-Coding RNAs in Atrophy. Biomed. Pharmacother. 2021, 141, 111820. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.J.; Cullen, J.; Barnholtz-Sloan, J.S.; Ostrom, Q.T.; Langer, C.E.; Turner, M.C.; McKean-Cowdin, R.; Fisher, J.L.; Lupo, P.J.; Partap, S.; et al. Childhood Brain Tumor Epidemiology: A Brain Tumor Epidemiology Consortium Review. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2014, 23, 2716–2736. [Google Scholar] [CrossRef]
- Northcott, P.A.; Robinson, G.W.; Kratz, C.P.; Mabbott, D.J.; Pomeroy, S.L.; Clifford, S.C.; Rutkowski, S.; Ellison, D.W.; Malkin, D.; Taylor, M.D.; et al. Medulloblastoma. Nat. Rev. Dis. Primer 2019, 5, 11. [Google Scholar] [CrossRef]
- Song, H.; Han, L.-M.; Gao, Q.; Sun, Y. Long Non-Coding RNA CRNDE Promotes Tumor Growth in Medulloblastoma. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 2588–2597. [Google Scholar]
- Sun, X.; Fan, W.; An, Z.; Sun, Y. Inhibition of Long Noncoding RNA CRNDE Increases Chemosensitivity of Medulloblastoma Cells by Targeting miR-29c-3p. Oncol. Res. 2020, 28, 95–102. [Google Scholar] [CrossRef]
- Dalan, A.B.; Gulluoglu, S.; Tuysuz, E.C.; Kuskucu, A.; Yaltirik, C.K.; Ozturk, O.; Ture, U.; Bayrak, O.F. Simultaneous Analysis of miRNA-mRNA in Human Meningiomas by Integrating Transcriptome: A Relationship between PTX3 and miR-29c. BMC Cancer 2017, 17, 207. [Google Scholar] [CrossRef]
- Li, W.; Yi, J.; Zheng, X.; Liu, S.; Fu, W.; Ren, L.; Li, L.; Hoon, D.S.B.; Wang, J.; Du, G. miR-29c Plays a Suppressive Role in Breast Cancer by Targeting the TIMP3/STAT1/FOXO1 Pathway. Clin. Epigenetics 2018, 10, 64. [Google Scholar] [CrossRef]
- Zhang, J.X.; Mai, S.J.; Huang, X.X.; Wang, F.W.; Liao, Y.J.; Lin, M.C.; Kung, H.F.; Zeng, Y.X.; Xie, D. MiR-29c Mediates Epithelial-to-Mesenchymal Transition in Human Colorectal Carcinoma Metastasis via PTP4A and GNA13 Regulation of β-Catenin Signaling. Ann. Oncol. 2014, 25, 2196–2204. [Google Scholar] [CrossRef]
- Laneve, P.; Po, A.; Favia, A.; Legnini, I.; Alfano, V.; Rea, J.; Carlo, V.D.; Bevilacqua, V.; Miele, E.; Mastronuzzi, A.; et al. The Long Noncoding RNA Linc-NeD125 Controls the Expression of Medulloblastoma Driver Genes by microRNA Sponge Activity. Oncotarget 2017, 8, 31003–31015. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, T.; Wang, S.; Xiong, Y.; Zhang, R.; Zhang, X.; Zhao, J.; Yang, A.-G.; Wang, L.; Jia, L. Nkx2-2as Suppression Contributes to the Pathogenesis of Sonic Hedgehog Medulloblastoma. Cancer Res. 2018, 78, 962–973. [Google Scholar] [CrossRef] [PubMed]
- Zhengyuan, X.; Hu, X.; Qiang, W.; Nanxiang, L.; Junbin, C.; Wangming, Z. Silencing of Urothelial Carcinoma Associated 1 Inhibits the Proliferation and Migration of Medulloblastoma Cells. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2017, 23, 4454–4461. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Zhang, R.; Zhang, C.; Zhao, L.; Zhang, Y. Long Noncoding RNA CCAT1 Promotes Cell Proliferation and Metastasis in Human Medulloblastoma via MAPK Pathway. Tumori J. 2018, 104, 43–50. [Google Scholar] [CrossRef]
- Gao, R.; Zhang, R.; Zhang, C.; Liang, Y.; Tang, W. LncRNA LOXL1-AS1 Promotes the Proliferation and Metastasis of Medulloblastoma by Activating the PI3K/AKT Pathway. Anal. Cell. Pathol. Amst. 2018, 2018, 9275685. [Google Scholar] [CrossRef]
- Varon, M.; Levy, T.; Mazor, G.; Ben David, H.; Marciano, R.; Krelin, Y.; Prasad, M.; Elkabets, M.; Pauck, D.; Ahmadov, U.; et al. The Long Noncoding RNA TP73-AS1 Promotes Tumorigenicity of Medulloblastoma Cells. Int. J. Cancer 2019, 145, 3402–3413. [Google Scholar] [CrossRef]
- Li, B.; Shen, M.; Yao, H.; Chen, X.; Xiao, Z. Long Noncoding RNA TP73-AS1 Modulates Medulloblastoma Progression In Vitro And In Vivo By Sponging miR-494-3p And Targeting EIF5A2. OncoTargets Ther. 2019, 12, 9873–9885. [Google Scholar] [CrossRef]
- Zhang, J.; Li, N.; Fu, J.; Zhou, W. Long Noncoding RNA HOTAIR Promotes Medulloblastoma Growth, Migration and Invasion by Sponging miR-1/miR-206 and Targeting YY1. Biomed. Pharmacother. 2020, 124, 109887. [Google Scholar] [CrossRef]
- Wu, J.; Armstrong, T.S.; Gilbert, M.R. Biology and Management of Ependymomas. Neuro-Oncology 2016, 18, 902–913. [Google Scholar] [CrossRef]
- Pajtler, K.W.; Mack, S.C.; Ramaswamy, V.; Smith, C.A.; Witt, H.; Smith, A.; Hansford, J.R.; von Hoff, K.; Wright, K.D.; Hwang, E.; et al. The Current Consensus on the Clinical Management of Intracranial Ependymoma and Its Distinct Molecular Variants. Acta Neuropathol. 2017, 133, 5–12. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, C.; Xie, Y.; Jiang, W.; Huang, J.; Guo, S.; Xu, F.; Wang, J. Detecting the Long Non-Coding RNA Signature Related to Spinal Cord Ependymal Tumor Subtype Using a Genome-Wide Methylome Analysis Approach. Mol. Med. Rep. 2019, 20, 1531–1540. [Google Scholar] [PubMed]
- Malgulwar, P.B.; Nambirajan, A.; Singh, M.; Suri, V.; Sarkar, C.; Sharma, M.C. Expression and Clinical Significance of Translation Regulatory Long Non-Coding RNA 1 (TRERNA1) in Ependymomas. Pathol. Oncol. Res. 2020, 26, 1975–1981. [Google Scholar] [CrossRef]
- Chakravadhanula, M.; Ozols, V.V.; Hampton, C.N.; Zhou, L.; Catchpoole, D.; Bhardwaj, R.D. Expression of the HOX Genes and HOTAIR in Atypical Teratoid Rhabdoid Tumors and Other Pediatric Brain Tumors. Cancer Genet. 2014, 207, 425–428. [Google Scholar] [CrossRef] [PubMed]
- Collins, V.P.; Jones, D.T.W.; Giannini, C. Pilocytic Astrocytoma: Pathology, Molecular Mechanisms and Markers. Acta Neuropathol. 2015, 129, 775–788. [Google Scholar] [CrossRef] [PubMed]
- Paassen, I.; Williams, J.; Ríos Arceo, C.; Ringnalda, F.; Mercer, K.S.; Buhl, J.L.; Moreno, N.; Federico, A.; Franke, N.E.; Kranendonk, M.; et al. Atypical Teratoid/Rhabdoid Tumoroids Reveal Subgroup-Specific Drug Vulnerabilities. Oncogene 2023, 42, 1661–1671. [Google Scholar] [CrossRef]
- Cohen, K.J.; Jabado, N.; Grill, J. Diffuse Intrinsic Pontine Gliomas-Current Management and New Biologic Insights. Is There a Glimmer of Hope? Neuro-Oncology 2017, 19, 1025–1034. [Google Scholar] [CrossRef]
- Rashed, W.M.; Maher, E.; Adel, M.; Saber, O.; Zaghloul, M.S. Pediatric Diffuse Intrinsic Pontine Glioma: Where Do We Stand? Cancer Metastasis Rev. 2019, 38, 759–770. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, H.; Zhang, D. Identification of Novel Long Non-Coding RNA in Diffuse Intrinsic Pontine Gliomas by Expression Profile Analysis. Oncol. Lett. 2018, 16, 6401–6406. [Google Scholar] [CrossRef]
- Cao, S.; Wang, Y.; Li, J.; Lv, M.; Niu, H.; Tian, Y. Tumor-Suppressive Function of Long Noncoding RNA MALAT1 in Glioma Cells by Suppressing miR-155 Expression and Activating FBXW7 Function. Am. J. Cancer Res. 2016, 6, 2561–2574. [Google Scholar]
- Liao, K.; Lin, Y.; Gao, W.; Xiao, Z.; Medina, R.; Dmitriev, P.; Cui, J.; Zhuang, Z.; Zhao, X.; Qiu, Y.; et al. Blocking lncRNA MALAT1/miR-199a/ZHX1 Axis Inhibits Glioblastoma Proliferation and Progression. Mol. Ther. Nucleic Acids 2019, 18, 388–399. [Google Scholar] [CrossRef]
- Li, H.; Guan, C. HOTAIR Inhibits the Proliferation of Glioblastoma Cells by Targeting miR-219. Cancer Biomark. Sect. Dis. Markers 2020, 28, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, G.; Gao, Y.; Liang, H. HOTAIR/miR-125 Axis-mediated Hexokinase 2 Expression Promotes Chemoresistance in Human Glioblastoma. J. Cell. Mol. Med. 2020, 24, 5707–5717. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Huang, Y.-L. DGCR5 Suppresses the EMT of Pediatric Primary Glioblastoma Multiforme Cell and Serves as a Prognostic Biomarker. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 10024–10034. [Google Scholar] [CrossRef]
- Jing, S.-Y.; Lu, Y.-Y.; Yang, J.-K.; Deng, W.-Y.; Zhou, Q.; Jiao, B.-H. Expression of Long Non-Coding RNA CRNDE in Glioma and Its Correlation with Tumor Progression and Patient Survival. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 3992–3996. [Google Scholar] [PubMed]
- Zhang, X.; Sun, S.; Pu, J.K.S.; Tsang, A.C.O.; Lee, D.; Man, V.O.Y.; Lui, W.M.; Wong, S.T.S.; Leung, G.K.K. Long Non-Coding RNA Expression Profiles Predict Clinical Phenotypes in Glioma. Neurobiol. Dis. 2012, 48, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.K.; Pastori, C.; Penas, C.; Komotar, R.J.; Ivan, M.E.; Wahlestedt, C.; Ayad, N.G. Serum Long Noncoding RNA HOTAIR as a Novel Diagnostic and Prognostic Biomarker in Glioblastoma Multiforme. Mol. Cancer 2018, 17, 74. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-S.; Mendell, J.T. Antisense-Mediated Transcript Knockdown Triggers Premature Transcription Termination. Mol. Cell 2020, 77, 1044–1054.e3. [Google Scholar] [CrossRef]
- Lai, F.; Damle, S.S.; Ling, K.K.; Rigo, F. Directed RNase H Cleavage of Nascent Transcripts Causes Transcription Termination. Mol. Cell 2020, 77, 1032–1043.e4. [Google Scholar] [CrossRef]
- Dhuri, K.; Bechtold, C.; Quijano, E.; Pham, H.; Gupta, A.; Vikram, A.; Bahal, R. Antisense Oligonucleotides: An Emerging Area in Drug Discovery and Development. J. Clin. Med. 2020, 9, 2004. [Google Scholar] [CrossRef]
- Padmakumar, S.; Jones, G.; Pawar, G.; Khorkova, O.; Hsiao, J.; Kim, J.; Amiji, M.M.; Bleier, B.S. Minimally Invasive Nasal Depot (MIND) Technique for Direct BDNF AntagoNAT Delivery to the Brain. J. Control. Release 2021, 331, 176–186. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Khoshbakht, T.; Taheri, M.; Ghanbari, M. A Concise Review on the Role of BDNF-AS in Human Disorders. Biomed. Pharmacother. 2021, 142, 112051. [Google Scholar] [CrossRef] [PubMed]
- Pruunsild, P.; Kazantseva, A.; Aid, T.; Palm, K.; Timmusk, T. Dissecting the Human BDNF Locus: Bidirectional Transcription, Complex Splicing, and Multiple Promoters. Genomics 2007, 90, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Warner, K.D.; Hajdin, C.E.; Weeks, K.M. Principles for Targeting RNA with Drug-like Small Molecules. Nat. Rev. Drug Discov. 2018, 17, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Behr, M.; Zhou, J.; Xu, B.; Zhang, H. In Vivo Delivery of CRISPR-Cas9 Therapeutics: Progress and Challenges. Acta Pharm. Sin. B 2021, 11, 2150–2171. [Google Scholar] [CrossRef]
- Khatoon, Z.; Figler, B.; Zhang, H.; Cheng, F. Introduction to RNA-Seq and Its Applications to Drug Discovery and Development: RNA-Seq in Drug Discovery and Development. Drug Dev. Res. 2014, 75, 324–330. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pandini, C.; Rey, F.; Cereda, C.; Carelli, S.; Gandellini, P. Study of lncRNAs in Pediatric Neurological Diseases: Methods, Analysis of the State-of-Art and Possible Therapeutic Implications. Pharmaceuticals 2023, 16, 1616. https://doi.org/10.3390/ph16111616
Pandini C, Rey F, Cereda C, Carelli S, Gandellini P. Study of lncRNAs in Pediatric Neurological Diseases: Methods, Analysis of the State-of-Art and Possible Therapeutic Implications. Pharmaceuticals. 2023; 16(11):1616. https://doi.org/10.3390/ph16111616
Chicago/Turabian StylePandini, Cecilia, Federica Rey, Cristina Cereda, Stephana Carelli, and Paolo Gandellini. 2023. "Study of lncRNAs in Pediatric Neurological Diseases: Methods, Analysis of the State-of-Art and Possible Therapeutic Implications" Pharmaceuticals 16, no. 11: 1616. https://doi.org/10.3390/ph16111616