
Biological Evaluation of Valeriana Extracts from Argentina with Potent Cholinesterase Inhibition for the Treatment of Neurodegenerative Disorders and Their Comorbidities—The Case of Valeriana carnosa Sm. (Caprifoliaceae) Studied in Mice
 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
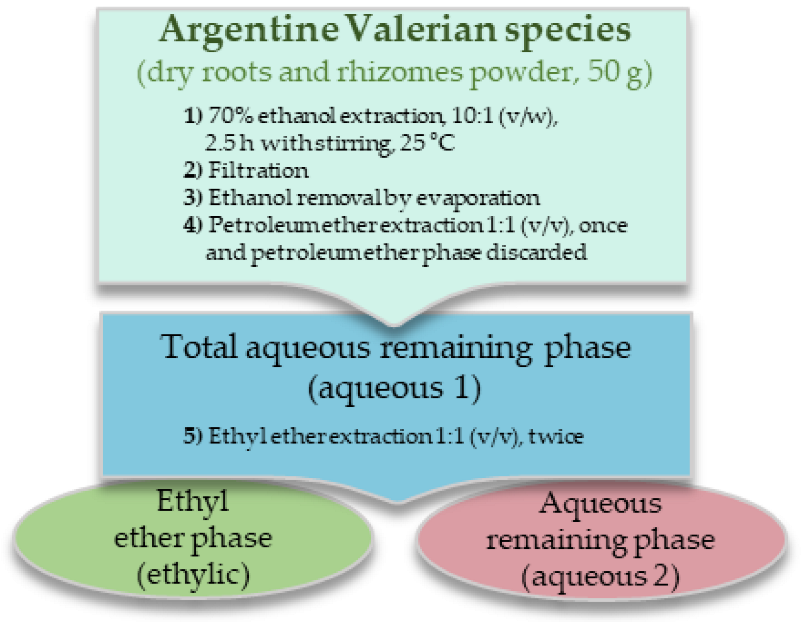
2.1. Plant Material and Extraction
2.2. Antioxidant Effect of Aqueous 1 Extracts
2.3. Inhibitory Activity of Extracts against AChE
2.4. Inhibitory Activity of the Extracts against BChE
2.5. Inhibitory Activity of Extracts against MAO
2.6. Inhibitory Activity of Extracts against Aβ1-42 Aggregation
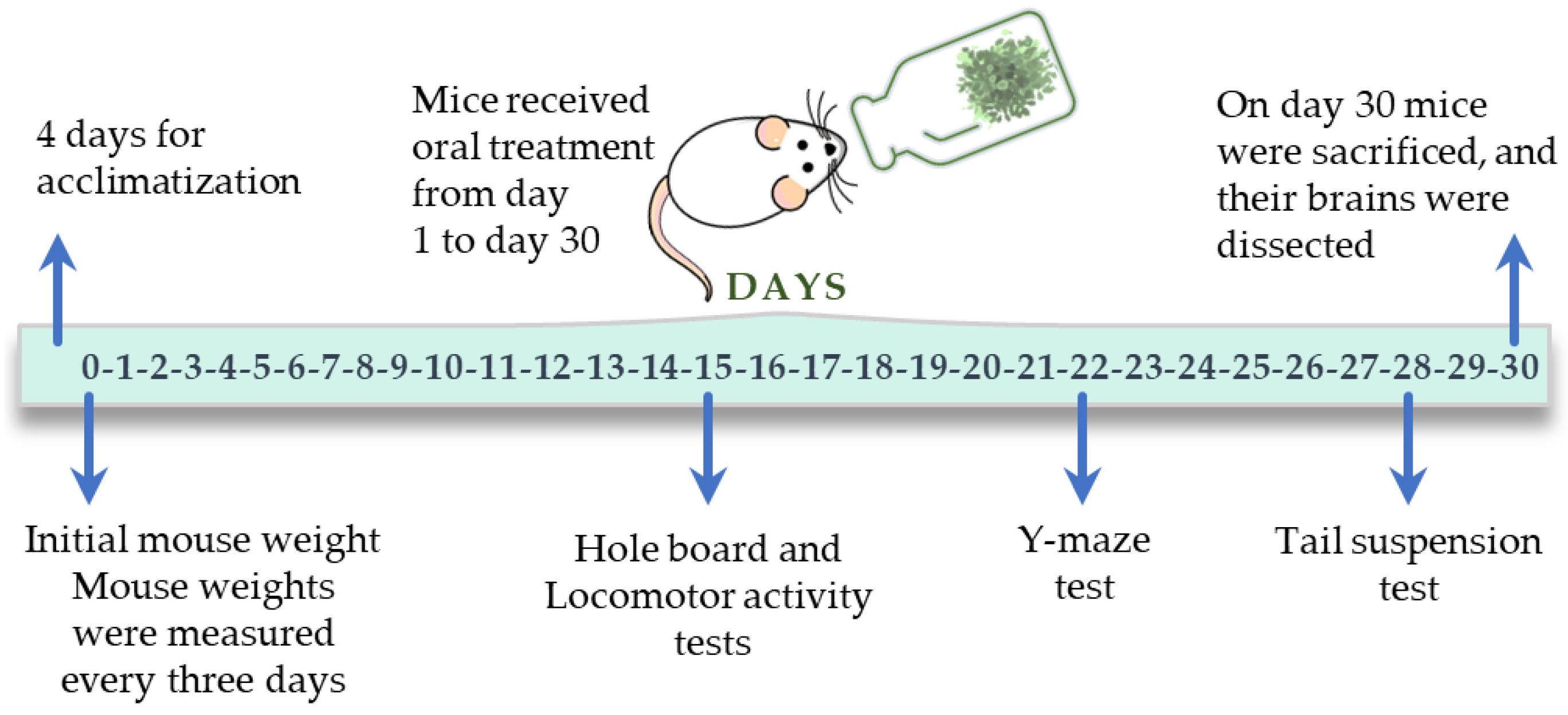
2.7. Effect of Chronic Treatments of V. Carnosa Aqueous 1 Extract in Mice
2.7.1. Stability Determination by High Performance Liquid Chromatography (HPLC) Fractionation
2.7.2. Fluid Intake and Mouse Weight
2.7.3. Effect of V. carnosa in the Hole Board and Locomotor Activity Assays
2.7.4. Effect of V. carnosa in the Y-maze Test
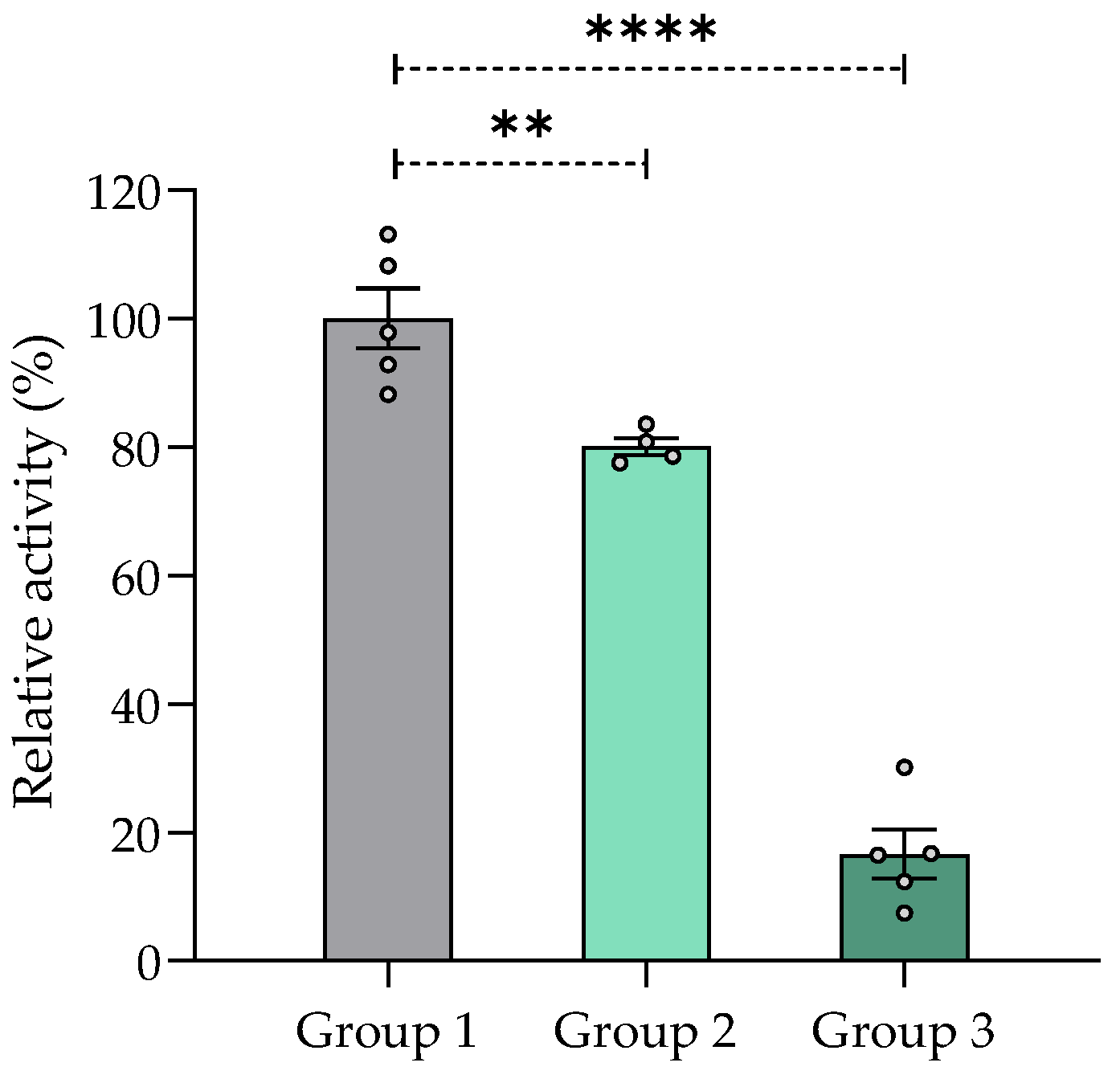
2.7.5. Effect of Chronic Treatment of V. carnosa on AChE Inhibition in the Mouse Brain
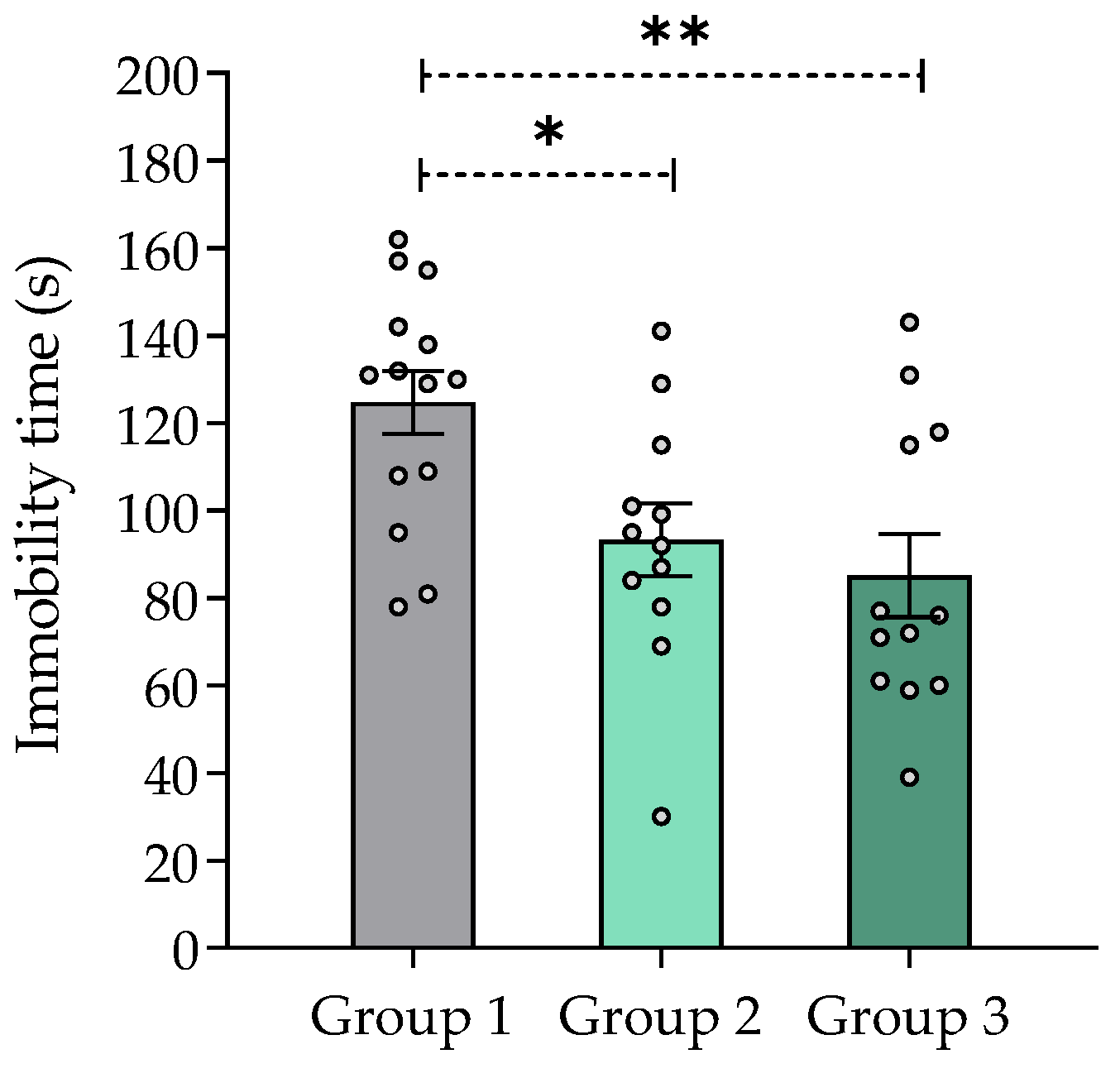
2.7.6. Effect of V. carnosa in the Tail Suspension Test
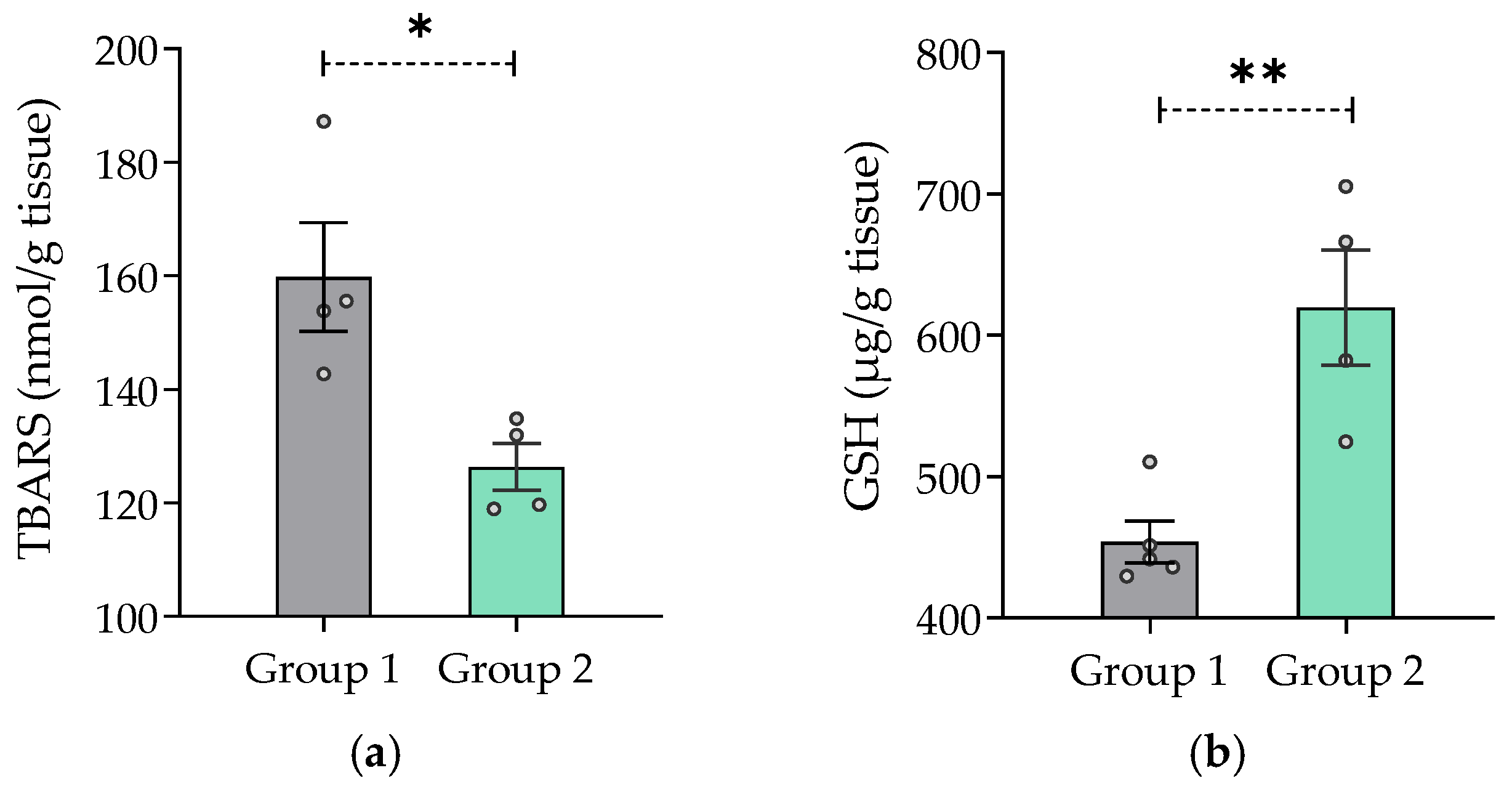
2.7.7. Effect of V. carnosa on Oxidative Stress Parameters
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. High Performance Liquid Chromatography (HPLC)
3.3. Animals
General
3.4. Chemical Assays
3.4.1. Determination of Antioxidant Capacity by the DPPH Method
3.4.2. Determination of Antioxidant Capacity by the ABTS•+ Method
3.4.3. Metal Fe2+ Chelating Ability
3.5. Biochemical Assays
3.5.1. Assessment of Lipid Damage through the Measurement of TBARS
3.5.2. Determination of Reduced GSH
3.5.3. Inhibition of Aβ1-42 Aggregation
3.5.4. ChE Inhibition
3.5.4.1. Recombinant Enzymes
3.5.4.2. AChE Inhibition in Mouse Brains
3.5.4.3. BChE Inhibition in Mouse Serum
3.5.5. In vitro hMAO Inhibition Assay
3.6. Pharmacological Studies
3.6.1. Protocol
3.6.2. Behavioral Studies
Hole Board Assay
Locomotor Activity Test
Y-Maze
Tail Suspension Test
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bisset, N.G. Herbal Drugs and Phytopharmaceuticals; Medpharm GmbH Scientific: Stuttgart, Germany, 1994. [Google Scholar]
- Houghton, P.J. The scientific basis for the reputed activity of Valerian. J. Pharm. Pharmacol. 1999, 51, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Yager, J.; Siegfreid, S.L.; DiMatteo, T.L. Use of alternative remedies by psychiatric patients: Illustrative vignettes and a discussion of the issues. Am. J. Psychiatry 1999, 156, 1432–1438. [Google Scholar] [CrossRef] [PubMed]
- Bounthanh, C.; Bergmann, C.; Beck, J.P.; Haag-Berrurier, M.; Anton, R. Valepotriates, a new class of cytotoxic and antitumor agents. Planta Med. 1981, 41, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Tortarolo, M.; Braun, R.; Hübner, G.E.; Maurer, H.R. In vitro effects of epoxide-bearing alepotriates on mouse early hematopoietic progenitor cells and human T-lymphocytes. Arch. Toxicol. 1982, 51, 37–42. [Google Scholar] [CrossRef]
- Morazzoni, P.; Bombardelli, E. Valeriana officinalis: Traditional used and recent evaluation of activity. Fitoterapia 1995, 66, 99–112. [Google Scholar]
- Hattesohl, M.; Feistel, B.; Sievers, H.; Lehnfeld, R.; Hegger, M.; Winterhoff, H. Extracts of Valeriana officinalis L. s.l. show anxiolytic and antidepressant effects but neither sedative nor myorelaxant properties. Phytomedicine 2008, 15, 2–15. [Google Scholar] [CrossRef]
- Li, J.; Li, X.; Wang, C.; Zhang, M.; Ye, M.; Wang, Q. The potential of Valeriana as a traditional Chinese medicine: Traditional clinical applications, bioactivities, and phytochemistry. Front. Pharmacol. 2022, 13, 973138. [Google Scholar] [CrossRef]
- Fernández, S.; Wasowski, C.; Paladini, A.C.; Marder, M. Sedative and sleep-enhancing properties of linarin, a flavonoid-isolated from Valeriana officinalis. Pharmacol. Biochem. Behav. 2004, 77, 399–404. [Google Scholar] [CrossRef]
- Marcucci, C.; Relats, J.M.A.; Bach, H.G.; Kamecki, F.; Varela, B.G.; Wagner, M.L.; Pastore, V.; Colettis, N.; Ricco, R.A.; Marder, M. Neurobehavioral evaluation and phytochemical characterization of a series of argentine valerian species. Heliyon 2020, 6, e05691. [Google Scholar] [CrossRef]
- Marder, M.; Viola, H.; Wasowski, C.; Fernández, S.; Medina, J.H.; Paladini, A.C. 6-methylapigenin and hesperidin: New valeriana flavonoids with activity on the CNS. Pharmacol. Biochem. Behav. 2003, 75, 537–545. [Google Scholar] [CrossRef]
- Verma, R.S.; Verma, R.K.; Padalia, R.C.; Chauhan, A.; Singh, A.; Singh, H.P. Chemical diversity in the essential oil of Indian valerian (Valeriana jatamansi Jones). Chem. Biodivers. 2011, 8, 1921–1929. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Zhang, Z.X.; Chen, T.; Ye, J.; Dai, W.X.; Shan, L.; Su, J.; Shen, Y.H.; Li, H.L.; Liu, R.H.; et al. Characterization of chlorinated valepotriates from Valeriana jatamansi. Phytochemistry 2013, 85, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Huang, R.; Han, C.; Lv, Y.; Zhao, Y.; Chen, Y. New Iridoid Triesters from Valeriana jatamansi. Helv. Chim. Acta 2005, 88, 1059–1062. [Google Scholar] [CrossRef]
- Borsini, O.H. Valerianaceae. In Flora Patagónica; Colección Científica INTA: Buenos Aires, Argentina, 1999; Volume 8. [Google Scholar]
- Kutschker, A.; Ezcurra, C.; Balzaretti, V. Valeriana (Valerianaceae) de los Andes australes: Biodiversidad y compuestos químicos. In Tradiciones Y Transformaciones en Etnobotánica; Pochettino, M., Ladio, A., Arenas, P., Eds.; CYTED: La Plata, Argentina, 2010; pp. 219–224. [Google Scholar]
- Breteler, M.M.B. Mapping Out Biomarkers for Alzheimer Disease. JAMA 2011, 305, 304–305. [Google Scholar] [CrossRef] [PubMed]
- Hampel, H.; Mesulam, M.M.; Cuello, A.C.; Farlow, M.R.; Giacobini, E.; Grossberg, G.T.; Khachaturian, A.S.; Vergallo, A.; Cavedo, E.; Snyder, P.J.; et al. The cholinergic system in the pathophysiology and treatment of Alzheimer’s disease. Brain 2018, 141, 1917–1933. [Google Scholar] [CrossRef]
- Giacobini, E. Cholinergic function and Alzheimer’s disease. Int. J. Geriatr. Psychiatry 2003, 18, S1–S5. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Bush, A.I. Alzheimer’s amyloid β-peptide (1–42): Involvement of methionine residue 35 in the oxidative stress and neurotoxicity properties of this peptide. Neurobiol. Aging 2004, 25, 563–568. [Google Scholar] [CrossRef]
- Xing, S.; Li, Q.; Xiong, B.; Chen, Y.; Feng, F.; Liu, W.; Sun, H. Structure and therapeutic uses of butyrylcholinesterase: Application in detoxification, Alzheimer’s disease, and fat metabolism. Med. Res. Rev. 2021, 41, 858–901. [Google Scholar] [CrossRef]
- Nguyen, K.; Hoffman, H.; Chakkamparambil, B.; Grossberg, G.T. Evaluation of rivastigmine in Alzheimer’s disease. Neurodegener. Dis. Manag. 2021, 11, 35–48. [Google Scholar] [CrossRef]
- Atri, A. Current and Future Treatments in Alzheimer’s Disease. Semin. Neurol. 2019, 39, 227–240. [Google Scholar] [CrossRef]
- Anighoro, A.; Bajorath, J.; Rastelli, G. Polypharmacology: Challenges and opportunities in drug discovery. J. Med. Chem. 2014, 57, 7874–7887. [Google Scholar] [CrossRef] [PubMed]
- Santin, Y.; Resta, J.; Parini, A.; Mialet-Perez, J. Monoamine oxidases in age-associated diseases: New perspectives for old enzymes. Ageing Res. Rev. 2021, 66, 101256. [Google Scholar] [CrossRef] [PubMed]
- Bolognesi, M.L. Harnessing Polypharmacology with Medicinal Chemistry. ACS Med. Chem. Lett. 2019, 10, 273–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bawa, P.; Pradeep, P.; Kumar, P.; Choonara, Y.E.; Modi, G.; Pillay, V. Multi-target therapeutics for neuropsychiatric and neurodegenerative disorders. Drug Discov. Today 2016, 21, 1886–1914. [Google Scholar] [CrossRef]
- Murphy, K.; Kubin, Z.J.; Shepherd, J.N.; Ettinger, R.H. Valeriana officinalis root extracts have potent anxiolytic effects in laboratory rats. Phytomedicine 2010, 17, 674–678. [Google Scholar] [CrossRef]
- De Souza, M.M.; Holzmann, I.; Cechinel Filho, V.; Mora, T.C.; Cáceres, A.; Martínez, J.V.; Cruz, S.M. Evaluation of Behavioral and Pharmacological Effects of Hydroalcoholic Extract of Valeriana prionophylla Standl. from Guatemala. Evid. Based. Complement. Alternat. Med. 2011, 2011, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Sah, S.P.; Mathela, C.S.; Chopra, K. Antidepressant effect of Valeriana wallichii patchouli alcohol chemotype in mice: Behavioural and biochemical evidence. J. Ethnopharmacol. 2011, 135, 197–200. [Google Scholar] [CrossRef]
- Subhan, F.; Karim, N.; Gilani, A.H.; Sewell, R.D.E. Terpenoid content of Valeriana wallichii extracts and antidepressant-like response profiles. Phytother. Res. 2010, 24, 686–691. [Google Scholar] [CrossRef]
- Khuda, F.; Iqbal, Z.; Khan, A.; Zakiullah, R.; Shah, Y.; Khan, A. Report: Screening of selected medicinal plants for their enzyme inhibitory potential—A validation of their ethnopharmacological uses. Pak. J. Pharm. Sci. 2014, 27, 593–596. [Google Scholar]
- Wang, P.C.; Ran, X.H.; Luo, H.R.; Ma, Q.Y.; Liu, Y.Q.; Zhou, J.; Zhao, Y.X. Phenolic compounds from the roots of valeriana officinalis var. latifolia. J. Braz. Chem. Soc. 2013, 24, 1544–1548. [Google Scholar] [CrossRef]
- Wang, C.; Xiao, Y.; Yang, B.; Wang, Z.; Wu, L.; Su, X.; Brantner, A.; Kuang, H.; Wang, Q. Isolation and screened neuroprotective active constituents from the roots and rhizomes of Valeriana amurensis. Fitoterapia 2014, 96, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, C.; Shu, Z.; Chan, K.; Huang, S.; Li, Y.; Xiao, Y.; Wu, L.; Kuang, H.; Sun, X. Valeriana amurensis improves Amyloid-beta 1-42 induced cognitive deficit by enhancing cerebral cholinergic function and protecting the brain neurons from apoptosis in mice. J. Ethnopharmacol. 2014, 153, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Casamiquela, R. Proyecto etnobotánico de la Patagonia. Primer informe. Asp. Técnicos Cult. Políticos Y Leg. La Bioprospección En La Argent. 1999, 55, 91–134. [Google Scholar] [CrossRef]
- Ladio, A.; Pieroni, A.; Price, L. Gathering of Wild Plant Foods with Medicinal Use in a Mapuche Community of Northwest Patagonia. In Eating and Healing: Traditional Food as Medicine; CRC Press: Boca Raton, FL, USA, 2006; ISBN 9780429175046. [Google Scholar]
- Tepe, B.; Daferera, D.; Sokmen, A.; Sokmen, M.; Polissiou, M. Antimicrobial and antioxidant activities of the essential oil and various extracts of Salvia tomentosa Miller (Lamiaceae). Food Chem. 2005, 90, 333–340. [Google Scholar] [CrossRef]
- Duarte-Jurado, A.P.; Gopar-Cuevas, Y.; Saucedo-Cardenas, O.; Loera-Arias, M.; Montes-De-oca-luna, R.; Garcia-Garcia, A.; Rodriguez-Rocha, H. Antioxidant therapeutics in parkinson’s disease: Current challenges and opportunities. Antioxidants 2021, 10, 453. [Google Scholar] [CrossRef]
- Thusoo, S.; Gupta, S.; Sudan, R.; Kour, J.; Bhagat, S.; Hussain, R.; Bhagat, M. Antioxidant activity of essential oil and extracts of Valeriana jatamansi Roots. Biomed Res. Int. 2014, 2014, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Das, J.; Mao, A.A.; Handique, P.J. Terpenoid compositions and antioxidant activities of two Indian valerian oils from the Khasi Hills of north-east India. Nat. Prod. Commun. 2011, 6, 129–132. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.-C.; Ran, X.-H.; Chen, R.; Luo, H.-R.; Liu, Y.-Q.; Zhou, J.; Zhao, Y.-X. Germacrane-type sesquiterpenoids from the roots of Valeriana officinalis var. latifolia. J. Nat. Prod. 2010, 73, 1563–1567. [Google Scholar] [CrossRef]
- Adedapo, A.A.; Jimoh, F.O.; Koduru, S.; Afolayan, A.J.; Masika, P.J. Antibacterial and antioxidant properties of the methanol extracts of the leaves and stems of Calpurnia aurea. BMC Complement. Altern. Med. 2008, 8, 53. [Google Scholar] [CrossRef] [Green Version]
- Jain, V.; Dutta, R.; Maheshwari, D.T.; Meena, D.K.; Misra, K.; Sadashiva Yogendra Kumar, M. Valeriana wallichii extract inhibits tert-BOOH induced oxidative damage and cytotoxicity. Front. Biosci. 2018, 10, 469–480. [Google Scholar] [CrossRef]
- Sen-Utsukarci, B.; Taskin, T.; Goger, F.; Tabanca, N.; Estep, A.S.; Kessler, S.M.; Akbal-Dagistan, O.; Bardakci, H.; Kurkcuoglu, M.; Becnel, J.; et al. Chemical composition and antioxidant, cytotoxic, and insecticidal potential of Valeriana alliariifolia in Turkey. Arh. Za Hig. Rada I Toksikol. 2019, 70, 207–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhao, J.; Liu, H.; Zhou, L.; Liu, Z.; Wang, J.; Han, J.; Yu, Z.; Yang, F. Chemical analysis and biological activity of the essential oils of two valerianaceous species from China: Nardostachys chinensis and valeriana officinalis. Molecules 2010, 15, 6411–6422. [Google Scholar] [CrossRef] [PubMed]
- De Ávila, J.M.; Pereira, A.O.; Zachow, L.L.; Gehm, A.Z.; Santos, M.Z.; Mostardeiro, M.A.; Back, D.; Morel, A.F.; Dalcol, I.I. Chemical constituents from Valeriana polystachya Smith and evaluation of their effects on the acetylcholinesterase and prolyl oligopeptidase activities. Fitoterapia 2018, 131, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.W.; Yang, L.; Wu, Z.K.; Wei-Gao; Zi, C.T.; Yang, D.; Luo, H.R.; Zhou, J.; Hu, J.M. Iridoids and sesquiterpenoids from the roots of Valeriana jatamansi Jones. Fitoterapia 2015, 102, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Khan, H.U.; Khan, F.A.; Shah, A.; Wadood, A.; Ahmad, S.; Almehmadi, M.; Alsaiari, A.A.; Shah, F.U.; Kamran, N. Anti-Alzheimer and Antioxidant Effects of Nelumbo nucifera L. Alkaloids, Nuciferine and Norcoclaurine in Alloxan-Induced Diabetic Albino Rats. Pharmaceuticals 2022, 15, 1205. [Google Scholar] [CrossRef]
- Jung, H.A.; Karki, S.; Kim, J.H.; Choi, J.S. BACE1 and cholinesterase inhibitory activities of Nelumbo nucifera embryos. Arch. Pharm. Res. 2015, 38, 1178–1187. [Google Scholar] [CrossRef]
- Khan, F.A.; Khan, N.M.; Ahmad, S.; Nasruddin; Aziz, R.; Ullah, I.; Almehmadi, M.; Allahyani, M.; Alsaiari, A.A.; Aljuaid, A. Phytochemical Profiling, Antioxidant, Antimicrobial and Cholinesterase Inhibitory Effects of Essential Oils Isolated from the Leaves of Artemisia scoparia and Artemisia absinthium. Pharmaceuticals 2022, 15, 1221. [Google Scholar] [CrossRef]
- Li, Q.; Tu, Y.; Zhu, C.; Luo, W.; Huang, W.; Liu, W.; Li, Y. Cholinesterase, β-amyloid aggregation inhibitory and antioxidant capacities of Chinese medicinal plants. Ind. Crop. Prod. 2017, 108, 512–519. [Google Scholar] [CrossRef]
- Arendt, T.; Brückner, M.K.; Lange, M.; Bigl, V. Changes in acetylcholinesterase and butyrylcholinesterase in Alzheimer’s disease resemble embryonic development-A study of molecular forms. Neurochem. Int. 1992, 21, 381–396. [Google Scholar] [CrossRef]
- Giacobini, E. Cholinesterase inhibitors: New roles and therapeutic alternatives. Pharmacol. Res. 2004, 50, 433–440. [Google Scholar] [CrossRef]
- Finberg, J.P.M.; Rabey, J.M. Inhibitors of MAO-A and MAO-B in Psychiatry and Neurology. Front. Pharmacol. 2016, 7, 340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Y.Y.; Jenner, P.; Chen, S. Di Monoamine Oxidase-B Inhibitors for the Treatment of Parkinson’s Disease: Past, Present, and Future. J. Park. Dis. 2022, 12, 477. [Google Scholar] [CrossRef]
- Medvedev, A.; Kopylov, A.; Buneeva, O.; Kurbatov, L.; Tikhonova, O.; Ivanov, A.; Zgoda, V. A Neuroprotective Dose of Isatin Causes Multilevel Changes Involving the Brain Proteome: Prospects for Further Research. Int. J. Mol. Sci. 2020, 21, 4187. [Google Scholar] [CrossRef] [PubMed]
- Matveychuk, D.; MacKenzie, E.M.; Kumpula, D.; Song, M.S.; Holt, A.; Kar, S.; Todd, K.G.; Wood, P.L.; Baker, G.B. Overview of the Neuroprotective Effects of the MAO-Inhibiting Antidepressant Phenelzine. Cell. Mol. Neurobiol. 2022, 42, 225–242. [Google Scholar] [CrossRef]
- Cott, M. In vitro receptor binding and enzyme inhibition by hypericum perforation extract. Pharmacopsychiatry 1997, 30, 108–112. [Google Scholar] [CrossRef]
- Herraiz, T.; Guillén, H. Monoamine Oxidase-A Inhibition and Associated Antioxidant Activity in Plant Extracts with Potential Antidepressant Actions. Biomed Res. Int. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Islam, F.; Khadija, J.F.; Rashid, H.O.; Rahaman, S.; Nafady, M.H.; Islam, R.; Akter, A.; Bin Emran, T.; Wilairatana, P.; Mubarak, M.S. Bioactive Compounds and Their Derivatives: An Insight into Prospective Phytotherapeutic Approach against Alzheimer’s Disease. Oxidative Med. Cell. Longev. 2022, 2022, 1–22. [Google Scholar] [CrossRef]
- Hussain, H.; Ahmad, S.; Shah, S.W.A.; Ghias, M.; Ullah, A.; Rahman, S.U.; Kamal, Z.; Khan, F.A.; Khan, N.M.; Muhammad, J.; et al. Neuroprotective potential of synthetic mono-carbonyl curcumin analogs assessed by molecular docking studies. Molecules 2021, 26, 7168. [Google Scholar] [CrossRef]
- Velander, P.; Wu, L.; Henderson, F.; Zhang, S.; Bevan, D.R.; Xu, B. Natural product-based amyloid inhibitors. Biochem. Pharmacol. 2017, 139, 40–55. [Google Scholar] [CrossRef] [Green Version]
- Braidy, N.; Jugder, B.-E.; Poljak, A.; Jayasena, T.; Mansour, H.; Nabavi, S.M.; Sachdev, P.; Grant, R. Resveratrol as a Potential Therapeutic Candidate for the Treatment and Management of Alzheimer’s Disease. Curr. Top. Med. Chem. 2016, 16, 1951–1960. [Google Scholar] [CrossRef]
- Jia, Y.; Wang, N.; Liu, X. Resveratrol and amyloid-beta: Mechanistic insights. Nutrients 2017, 9, 1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Liao, W.; Chen, X.; Yue, H.; Li, S.; Ding, K. An arabinogalactan from fruits of Lycium barbarum L. inhibits production and aggregation of Aβ 42. Carbohydr. Polym. 2018, 195, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Okada, M. In vitro screening on amyloid beta modulation of aqueous extracts from plant seeds. J. Pharm. Bioallied Sci. 2016, 8, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Molares, S.; Ladio, A.H. Valeriana carnosa Sm. In Medicinal and Aromatic Plants of South America; Albuquerque, U., Patil, U., Máthé, Á., Eds.; Springer Nature: Dordrecht, The Netherlands, 2018. [Google Scholar]
- Guajardo, J.; Gastaldi, B.; González, S.; Nagahama, N. Variability of phenolic compounds at different phenological stages in two populations of Valeriana carnosa Sm. (Valerianoideae, Caprifoliaceae) in Patagonia. Boletín Latinoam. Y Del Caribe De Plantas Med. Y Aromáticas 2018, 17, 381–393. [Google Scholar]
- Taslimi, P.; Caglayan, C.; Gulcin, İ. The impact of some natural phenolic compounds on carbonic anhydrase, acetylcholinesterase, butyrylcholinesterase, and α-glycosidase enzymes: An antidiabetic, anticholinergic, and antiepileptic study. J. Biochem. Mol. Toxicol. 2017, 31, e21995. [Google Scholar] [CrossRef] [PubMed]
- Carradori, S.; Gidaro, M.C.; Petzer, A.; Costa, G.; Guglielmi, P.; Chimenti, P.; Alcaro, S.; Petzer, J.P. Inhibition of Human Monoamine Oxidase: Biological and Molecular Modeling Studies on Selected Natural Flavonoids. J. Agric. Food Chem. 2016, 64, 9004–9011. [Google Scholar] [CrossRef]
- Lee, B.; Choi, G.M.; Sur, B. Antidepressant-Like Effects of Hesperidin in Animal Model of Post-Traumatic Stress Disorder. Chin. J. Integr. Med. 2021, 27, 39–46. [Google Scholar] [CrossRef]
- Chakraborty, S.; Bandyopadhyay, J.; Chakraborty, S.; Basu, S. Multi-target screening mines hesperidin as a multi-potent inhibitor: Implication in Alzheimer’s disease therapeutics. Eur. J. Med. Chem. 2016, 121, 810–822. [Google Scholar] [CrossRef]
- Zheng, H.; Zhen, X.-T.; Chen, Y.; Zhu, S.-C.; Ye, L.-H.; Yang, S.-W.; Wang, Q.-Y.; Cao, J. In situ antioxidation-assisted matrix solid-phase dispersion microextraction and discrimination of chiral flavonoids from citrus fruit via ion mobility quadrupole time-of-flight high-resolution mass spectrometry. Food Chem. 2021, 343, 128422. [Google Scholar] [CrossRef]
- Said, U.Z.; Saada, H.N.; Abd-Alla, M.S.; Elsayed, M.E.; Amin, A.M. Hesperidin attenuates brain biochemical changes of irradiated rats. Int. J. Radiat. Biol. 2012, 88, 613–618. [Google Scholar] [CrossRef]
- Donato, F.; de Gomes, M.G.; Goes, A.T.R.; Filho, C.B.; Del Fabbro, L.; Antunes, M.S.; Souza, L.C.; Boeira, S.P.; Jesse, C.R. Hesperidin exerts antidepressant-like effects in acute and chronic treatments in mice: Possible role of l-arginine-NO-cGMP pathway and BDNF levels. Brain Res. Bull. 2014, 104, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Hemanth Kumar, B.; Dinesh Kumar, B.; Diwan, P.V. Hesperidin, a citrus flavonoid, protects against l-methionine-induced hyperhomocysteinemia by abrogation of oxidative stress, endothelial dysfunction and neurotoxicity in Wistar rats. Pharm. Biol. 2017, 55, 146–155. [Google Scholar] [CrossRef] [Green Version]
- Oboh, G.; Agunloye, O.M.; Akinyemi, A.J.; Ademiluyi, A.O.; Adefegha, S.A. Comparative study on the inhibitory effect of caffeic and chlorogenic acids on key enzymes linked to Alzheimer’s disease and some pro-oxidant induced oxidative stress in rats’ brain-in vitro. Neurochem. Res. 2013, 38, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Engelbrecht, I.; Petzer, J.P.; Petzer, A. Evaluation of Selected Natural Compounds as Dual Inhibitors of Catechol-O-Methyltransferase and Monoamine Oxidase. Cent. Nerv. Syst. Agents Med. Chem. 2019, 19, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.-H.; Lee, H.-K.; Kim, J.-A.; Hong, S.-I.; Kim, H.-C.; Jo, T.-H.; Park, Y.-I.; Lee, C.-K.; Kim, Y.-B.; Lee, S.-Y.; et al. Neuroprotective effects of chlorogenic acid on scopolamine-induced amnesia via anti-acetylcholinesterase and anti-oxidative activities in mice. Eur. J. Pharmacol. 2010, 649, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Nazir, N.; Zahoor, M.; Nisar, M.; Karim, N.; Latif, A.; Ahmad, S.; Uddin, Z. Evaluation of neuroprotective and anti-amnesic effects of Elaeagnus umbellata Thunb. On scopolamine-induced memory impairment in mice. BMC Complement. Med. Ther. 2020, 20, 1–17. [Google Scholar] [CrossRef]
- Wu, J.; Chen, H.; Li, H.; Tang, Y.; Yang, L.; Cao, S.; Qin, D. Antidepressant Potential of Chlorogenic Acid-Enriched Extract from Eucommia ulmoides Oliver Bark with Neuron Protection and Promotion of Serotonin Release through Enhancing Synapsin I Expression. Molecules 2016, 21, 260. [Google Scholar] [CrossRef] [Green Version]
- Wasowski, C.; Loscalzo, L.M.; Higgs, J.; Marder, M. Chronic Intraperitoneal and Oral Treatments with Hesperidin Induce Central Nervous System Effects in Mice. Phytother. Res. 2012, 26, 308–312. [Google Scholar] [CrossRef]
- Abu Almaaty, A.H.; Mosaad, R.M.; Hassan, M.K.; Ali, E.H.A.; Mahmoud, G.A.; Ahmed, H.; Anber, N.; Alkahtani, S.; Abdel-Daim, M.M.; Aleya, L.; et al. Urtica dioica extracts abolish scopolamine-induced neuropathies in rats. Environ. Sci. Pollut. Res. Int. 2021, 28, 18134–18145. [Google Scholar] [CrossRef]
- Deutsch, J.A.; Rocklin, K.W. Amnesia induced by Scopolamine and its Temporal Variations. Nature 1967, 216, 89–90. [Google Scholar] [CrossRef]
- Huang, Z.-B.; Wang, H.; Rao, X.-R.; Zhong, G.-F.; Hu, W.-H.; Sheng, G.-Q. Different effects of scopolamine on the retrieval of spatial memory and fear memory. Behav. Brain Res. 2011, 221, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Sarter, M.; Bruno, J.P. Cortical cholinergic inputs mediating arousal, attentional processing and dreaming: Differential afferent regulation of the basal forebrain by telencephalic and brainstem afferents. Neuroscience 1999, 95, 933–952. [Google Scholar] [CrossRef] [PubMed]
- Sarter, M.; Bruno, J.P.; Givens, B. Attentional functions of cortical cholinergic inputs: What does it mean for learning and memory? Neurobiol. Learn. Mem. 2003, 80, 245–256. [Google Scholar] [CrossRef]
- Konrath, E.L.; Neves, B.M.; Lunardi, P.S.; Passos, C.D.S.; Simões-Pires, A.; Ortega, M.G.; Gonçalves, C.A.; Cabrera, J.L.; Moreira, J.C.F.; Henriques, A.T. Investigation of the in vitro and ex vivo acetylcholinesterase and antioxidant activities of traditionally used Lycopodium species from South America on alkaloid extracts. J. Ethnopharmacol. 2012, 139, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Santos, W.P.; Carvalho, A.C.D.S.; Estevam, C.D.S.; Santana, A.E.G.; Marçal, R.M. In vitro and ex vivo anticholinesterase activities of Erythrina velutina leaf extracts. Pharm. Biol. 2012, 50, 919–924. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Wang, X.; Liu, Y.; Di, X. Linarin Inhibits the Acetylcholinesterase Activity In-vitro and Ex-vivo. Iran. J. Pharm. Res. 2015, 14, 949–954. [Google Scholar] [PubMed]
- Müller, L.G.; Salles, L.A.; Stein, A.C.; Betti, A.H.; Sakamoto, S.; Cassel, E.; Vargas, R.F.; Von Poser, G.L.; Rates, S.M.K. Antidepressant-like effect of Valeriana glechomifolia Meyer (Valerianaceae) in mice. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2012, 36, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Contador-Castillo, I.; Fernández-Calvo, B.; Cacho-Gutiérrez, L.J.; Ramos-Campos, F.; Hernández-Martín, L. Depression in Alzheimer type-dementia: Is there any effect on memory performance. Rev. Neurol. 2009, 49, 505–510. [Google Scholar] [CrossRef]
- Chen, Z.; Zhong, C. Oxidative stress in Alzheimer’s disease. Neurosci. Bull. 2014, 30, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Ayyathan, D.M.; Chandrasekaran, R.; Thiagarajan, K. Neuroprotective effect of Tagara, an Ayurvedic drug against methyl mercury induced oxidative stress using rat brain mitochondrial fractions. BMC Complement. Altern. Med. 2015, 15, 268. [Google Scholar] [CrossRef] [Green Version]
- Sudati, J.H.; Fachinetto, R.; Pereira, R.P.; Boligon, A.A.; Athayde, M.L.; Soares, F.A.; Barbosa, N.B.D.V.; Rocha, J.B.T. In vitro antioxidant activity of Valeriana officinalis against different neurotoxic agents. Neurochem. Res. 2009, 34, 1372–1379. [Google Scholar] [CrossRef] [PubMed]
- Malva, J.O.; Santos, S.; Macedo, T. Neuroprotective properties of Valeriana officinalis extracts. Neurotox. Res. 2004, 6, 131–140. [Google Scholar] [CrossRef]
- Von der Eltz, H.; Guder, H.-J.; Miihlegger, K. New Hydrolase Substrates. U.S. Patent 4,900,822, 13 February 1990. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Aarland, R.C.; Bañuelos-Hernández, A.E.; Fragoso-Serrano, M.; Sierra-Palacios, E.D.C.; De León-Sánchez, F.D.; Pérez-Flores, L.J.; Rivera-Cabrera, F.; Mendoza-Espinoza, J.A. Studies on phytochemical, antioxidant, anti-inflammatory, hypoglycaemic and antiproliferative activities of Echinacea purpurea and Echinacea angustifolia extracts. Pharm. Biol. 2017, 55, 649–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everett, J.; Lermyte, F.; Brooks, J.; Tjendana-Tjhin, V.; Plascencia-Villa, G.; Hands-Portman, I.; Donnelly, J.M.; Billimoria, K.; Perry, G.; Zhu, X.; et al. Biogenic metallic elements in the human brain? Sci. Adv. 2021, 7, 6707–6716. [Google Scholar] [CrossRef] [PubMed]
- Repetto, M.; Semprine, J.; Boveris, A. Lipid Peroxidation: Chemical Mechanism, Biological Implications and Analytical Determination. In Lipid Peroxidation; Catala, A., Ed.; IntechOpen: London, UK, 2012; pp. 3–30. ISBN 978-953-51-0716-3. [Google Scholar]
- Pagliosa, C.M.; Vieira, F.G.K.; Silveira, T.T.; Krieguer, J.R.; Medeiros, M.F.; da Silva, E.L. Elevated Iron Chelating Activity of Ilex paraguariensis Leaf Infusion: In vitro and in vivo Pilot Studies. Braz. Arch. Biol. Technol. 2021, 64, 1–13. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Abdelghany, A.K.; El-Nahass, E.S.; Ibrahim, M.A.; El-Kashlan, A.M.; Emeash, H.H.; Khalil, F. Neuroprotective role of medicinal plant extracts evaluated in a scopolamine-induced rat model of Alzheimer’s disease. Biomarkers 2022, 27, 773–783. [Google Scholar] [CrossRef]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A New and Rapid Colorimetric Determination of Acetylcholinesterase Activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Košak, U.; Knez, D.; Coquelle, N.; Brus, B.; Pišlar, A.; Nachon, F.; Brazzolotto, X.; Kos, J.; Colletier, J.-P.; Gobec, S. N-Propargylpiperidines with naphthalene-2-carboxamide or naphthalene-2-sulfonamide moieties: Potential multifunctional anti-Alzheimer’s agents. Bioorg. Med. Chem. 2017, 25, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Wasowski, C.; Marder, M.; Viola, H.; Medina, J.H.; Paladini, A.C. Isolation and Identification of 6-Methylapigenin, a Competitive Ligand for the Brain GABA A Receptors from Valeriana wallichii. Planta Med. 2002, 68, 934–936. [Google Scholar] [CrossRef] [PubMed]
- Fernández, S.P.; Wasowski, C.; Loscalzo, L.M.; Granger, R.E.; Johnston, G.A.R.; Paladini, A.C.; Marder, M.; Fernandez, S.P.; Wasowski, C.; Loscalzo, L.M.; et al. Central Nervous System Depressant Action of Flavonoid Glycosides. Eur. J. Pharmacol. 2006, 539, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Walrave, L.; Vinken, M.; Albertini, G.; de Bundel, D.; Leybaert, L.; Smolders, I.J. Inhibition of connexin43 hemichannels impairs spatial short-term memory without affecting spatial working memory. Front. Cell. Neurosci. 2016, 10, 288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Can, A.; Dao, D.T.; Terrillion, C.E.; Piantadosi, S.C.; Bhat, S.; Gould, T.D. The tail suspension test. J. Vis. Exp. 2012, 59, e3769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cryan, J.F.; Mombereau, C.; Vassout, A. The tail suspension test as a model for assessing antidepressant activity: Review of pharmacological and genetic studies in mice. Neurosci. Biobehav. Rev. 2005, 29, 571–625. [Google Scholar] [CrossRef]
- Committee for the Update of the Guide for the Care Animals and Use of Laboratory. Guide for the Care and Use of Laboratory Animals; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]

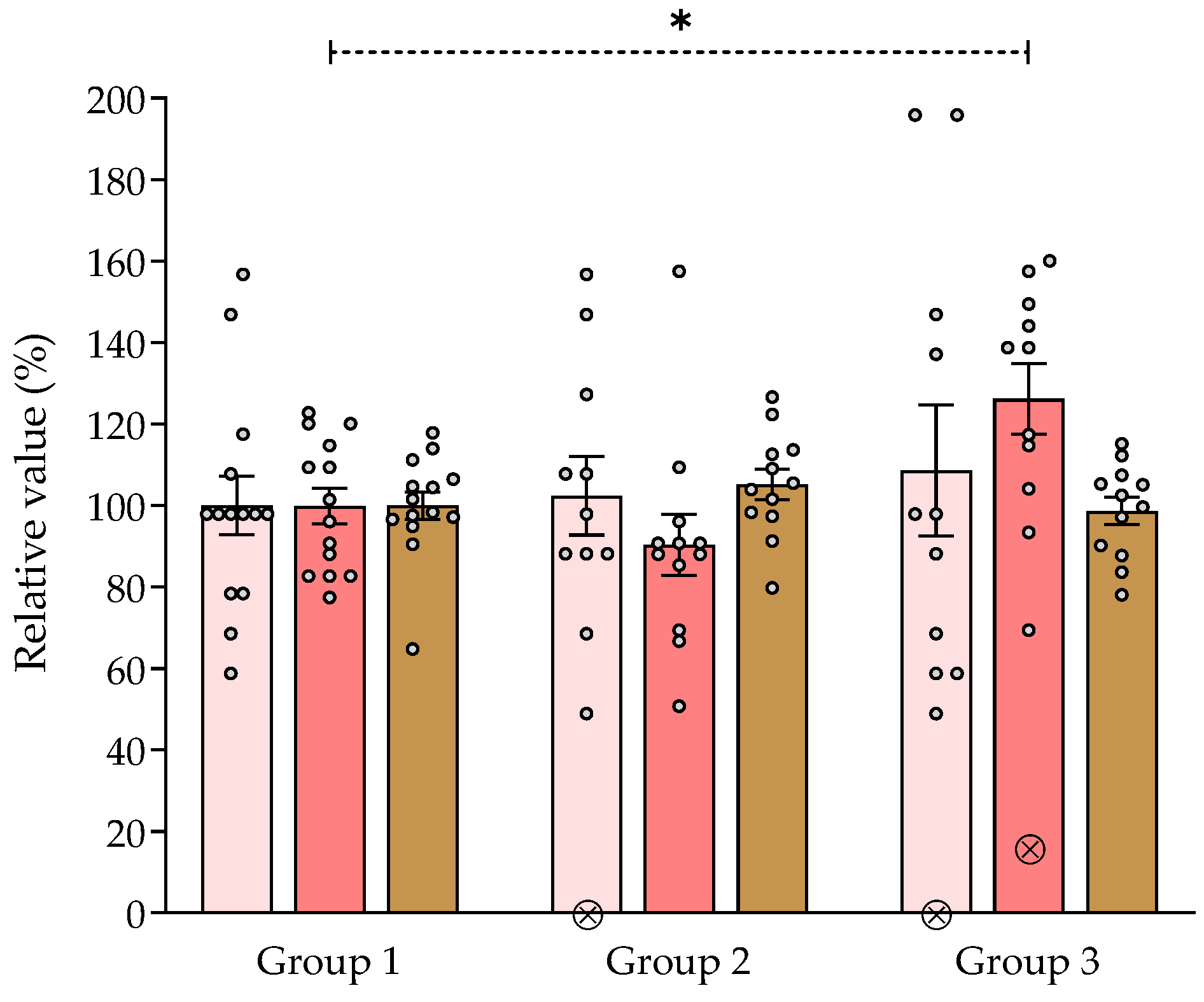
 ), percent spatial exploration (rearings) (
), percent spatial exploration (rearings) ( ) and percent distance traveled in the locomotor activity assay (
) and percent distance traveled in the locomotor activity assay ( ) recorded in 5-min sessions. * p < 0.05, significantly different from group 1. Dunnett’s multiple comparison test after ANOVA. ⊗: discarded values. ngroup 1 = 14, ngroup 2 = 12, ngroup 3 = 12.
), percent spatial exploration (rearings) () and percent distance traveled in the locomotor activity assay () recorded in 5-min sessions. * p < 0.05, significantly different from group 1. Dunnett’s multiple comparison test after ANOVA. ⊗: discarded values. ngroup 1 = 14, ngroup 2 = 12, ngroup 3 = 12.
) recorded in 5-min sessions. * p < 0.05, significantly different from group 1. Dunnett’s multiple comparison test after ANOVA. ⊗: discarded values. ngroup 1 = 14, ngroup 2 = 12, ngroup 3 = 12.
), percent spatial exploration (rearings) () and percent distance traveled in the locomotor activity assay () recorded in 5-min sessions. * p < 0.05, significantly different from group 1. Dunnett’s multiple comparison test after ANOVA. ⊗: discarded values. ngroup 1 = 14, ngroup 2 = 12, ngroup 3 = 12.
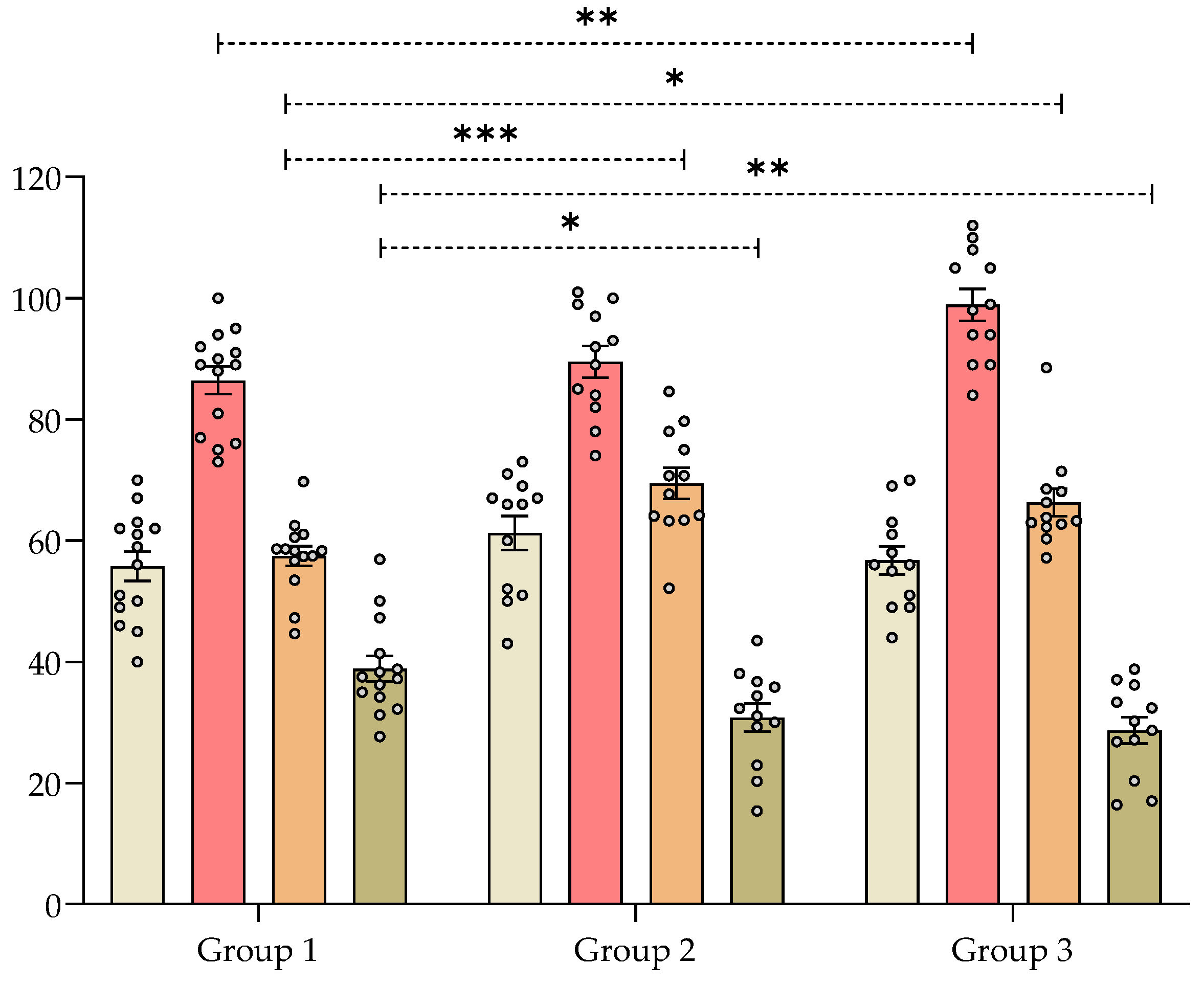
 ), number of rearings (
), number of rearings ( ), index of alternations (
), index of alternations ( ) and returns (
) and returns ( ) recorded in 8-min sessions. * p < 0.05, ** p < 0.008, *** p < 0.005 significantly different from group 1 (control). Dunnett’s multiple comparison test after ANOVA. ngroup 1 = 14, ngroup 2 = 12, ngroup 3 = 12.
), number of rearings (), index of alternations () and returns () recorded in 8-min sessions. * p < 0.05, ** p < 0.008, *** p < 0.005 significantly different from group 1 (control). Dunnett’s multiple comparison test after ANOVA. ngroup 1 = 14, ngroup 2 = 12, ngroup 3 = 12.
) recorded in 8-min sessions. * p < 0.05, ** p < 0.008, *** p < 0.005 significantly different from group 1 (control). Dunnett’s multiple comparison test after ANOVA. ngroup 1 = 14, ngroup 2 = 12, ngroup 3 = 12.
), number of rearings (), index of alternations () and returns () recorded in 8-min sessions. * p < 0.05, ** p < 0.008, *** p < 0.005 significantly different from group 1 (control). Dunnett’s multiple comparison test after ANOVA. ngroup 1 = 14, ngroup 2 = 12, ngroup 3 = 12.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Phenols 1 | DPPH | ABTS | Ferrozine | TBARS |
|---|---|---|---|---|---|
| mg Galic Acid/ 1 g Plant | EC50 (mg/mL) | EC50 (mg/mL) | IC50 (mg/mL) | IC50 (mg/mL) | |
| V. effusa | 125.52 ± 6.80 | 0.56 ± 0.04 | 0.41 ± 0.02 | 2.59 ± 0.08 | 25.04 ± 0.79 2 |
| V. ferax | 191.56 ± 8.99 | 0.57 ± 0.02 | 0.21 ± 0.13 | 2.10 ± 0.04 | 5.00 (2.38–10.51) 3 |
| V. macrorhiza | 252.85 ± 12.36 | 0.43 ± 0.02 | 0.13 ± 0.01 | 5.13 ± 0.09 | 0.20 (0.13–0.30) 3 |
| V. clarionifolia | 257.36 ± 19.55 | 0.30 ± 0.01 | 0.10 ± 0.01 | 1.3 ± 0.11 | 1.29 (0.81–2.05) 3 |
| V. carnosa | 890.25 ± 156.05 | 0.12 ± 0.02 | 0.04 ± 0.01 | 1.71 ± 0.10 | 0.18 (0.11–0.30) 3,4 |
| V. officinalis | 736.19 ± 34.27 | 0.32 ± 0.01 | 0.51 ± 0.06 | 4.76 ± 0.28 | 1.76 (1.25–2.48) 3 |
| Trolox 5 | ND | 0.016 ± 0.006 | 0.013 ± 0.004 | ND | ND |
| EDTA 5 | ND | ND | ND | 0.006 ± 0.001 | ND |
| Vitamin E 5 | ND | ND | ND | ND | 37.73 ± 0.41 2 |
| Sample | Extract | AChE IC50 (mg/mL) 1 | mAChE Inhibition (%) 2 | BChE IC50 (mg/mL) 1 | hBChE Inhibition (%) 2 |
|---|---|---|---|---|---|
| Mice Brain | Recombinant | Murine Plasma | Recombinant | ||
| V. effusa | Aqueous 1 | 12.69 (5.8–27.7) | NA | 0.86 (0.61–1.22) | 17.2 ± 3.1 |
| Aqueous 2 | ND | ND | 0.65 (0.45–0.92) | ND | |
| Ethylic | ND | 23.5 ± 3.4 | 0.41 (0.28–0.62) | 37.3 ± 3.1 | |
| V. ferax | Aqueous 1 | 5.45 (3.69–8.05) | ND | 0.53 (0.43–0.65) | 18.7 ± 3.1 |
| Aqueous 2 | ND | ND | 0.15 (0.11–0.2) | ND | |
| Ethylic | ND | 31.6 ± 4.1 | 0.025 (0.017–0.038) | 69.5 ± 7.2 | |
| V. macrorhiza | Aqueous 1 | 1.08 (0.49–2.37) | NA | 0.082 (0.07–0.10) | 24.4 ± 2.4 |
| Aqueous 2 | ND | ND | 0.95 (0.57–1.58) | ND | |
| Ethylic | ND | 29.9 ± 5.3 | 0.045 (0.004–0.005) | 78.7 ± 3.9 | |
| V. clarionifolia | Aqueous 1 | 1.29 (0.81–2.05) | 26.4 ± 2.7 | 0.0019 (0.0014–0.0024) | 64.3 ± 1.1 IC50 (mg/mL): 0.190 (0.120–0.300) |
| Aqueous 2 | ND | ND | 0.039 (0.030–0.050) | ND | |
| Ethylic | ND | 32.2 ± 4.4 | 0.00057 (0.00041–0.00081) | 67.9 ± 9.4 | |
| V. carnosa | Aqueous 1 | 6.71 (2.86–15.77) | NA | 1.46 (0.99–1.14) | NA |
| Aqueous 2 | ND | ND | 2.57 (1.11–6.0) | ND | |
| Ethylic | 6.92 (2.14–22.41) | 37.7 ± 5.8 | 0.26 (0.16–0.42) | 52.6 ± 9.5 | |
| V. officinalis | Aqueous 1 | 3.42 (1.11–10.6) | NA | 0.15 (0.13–0.18) | 88.7 ± 1.4 IC50 (mg/mL): 0.140 (0.110–0.180) |
| Aqueous 2 | ND | ND | 0.50 (0.33–0.75) | ND | |
| Ethylic | ND | ND | 0.0041 (0.0026–0.0067) | ND | |
| Tacrine 3 | ND | IC50 (µM): 0.140 ± 0.008 | ND | IC50 (µM): 0.023 ± 0.003 |
| Sample | Extract 1 | hMAO-A 2 | hMAO-B 2 |
|---|---|---|---|
| Inhibition (%) | Inhibition (%) | ||
| V. effusa | Aqueous 1 | 37.2 ± 1.1 | 23.5 ± 1.3 |
| Ethylic | 45.3 ± 0.9 | 14.2 ± 4.9 | |
| V. ferax | Aqueous 1 | 38.7 ± 0.1 | 22.1 ± 0.8 |
| Ethylic | 57.9 ± 0.7 | NA | |
| V. macrorhiza | Aqueous 1 | 24.7 ± 3.7 | 62.7 ± 2.5 |
| Ethylic | 68.6 ± 0.1 | 50.6 ± 3.6 | |
| V. clarionifolia | Aqueous 1 | 39.5 ± 0.6 | 39.9 ± 1.8 |
| Ethylic | 0.4 ± 0.7 | NA | |
| V. carnosa | Aqueous 1 | 41.6 ± 0.1 | 57.6 ± 5.4 |
| Ethylic | 69.5 ± 1.4 IC50 (mg/mL): 0.286 (0.213–0.384) 3 | 49.4 ± 4.2 | |
| V. officinalis | Aqueous 1 | 62.8 ± 0.4 | 57.5 ± 3.2 |
| Ethylic | ND | ND | |
| Clorgyline 4 | - | IC50 (µM): 0.00335 ± 0.00031 3 | IC50 (µM): 13.568 ± 1.157 3 |
| Pargyline 4 | - | IC50 (µM): 3.968 ± 0.275 3 | IC50 (µM): 0.195 ± 0.019 3 |
| L-deprenyl 4 | - | IC50 (µM): 62.664 ± 0.411 3 | IC50 (µM): 0.012 ± 0.004 3 |
| Isatin 4 | - | ND | IC50 (µM): 19.778 ± 1.108 3 |
| Sample | Aβ1–42 Aggregation 1 |
|---|---|
| Inhibition (%) | |
| V. effusa | 93.4% ± 4.1 |
| V. ferax | 47.8% ± 13.4 |
| V. macrorhiza | 32.0% ± 11.5 |
| V. clarionifolia | 81.6% ± 13.6 |
| V. carnosa | 65.7% ±12.2 |
| V. officinalis | 59.7% ± 2.6 |
| Resveratrol 3 | 93.9% ± 4.8 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marcucci, C.; Rademacher, M.; Kamecki, F.; Pastore, V.; Bach, H.G.; Ricco, R.A.; Wagner, M.L.; Knez, D.; Gobec, S.; Colettis, N.; et al. Biological Evaluation of Valeriana Extracts from Argentina with Potent Cholinesterase Inhibition for the Treatment of Neurodegenerative Disorders and Their Comorbidities—The Case of Valeriana carnosa Sm. (Caprifoliaceae) Studied in Mice. Pharmaceuticals 2023, 16, 129. https://doi.org/10.3390/ph16010129
Marcucci C, Rademacher M, Kamecki F, Pastore V, Bach HG, Ricco RA, Wagner ML, Knez D, Gobec S, Colettis N, et al. Biological Evaluation of Valeriana Extracts from Argentina with Potent Cholinesterase Inhibition for the Treatment of Neurodegenerative Disorders and Their Comorbidities—The Case of Valeriana carnosa Sm. (Caprifoliaceae) Studied in Mice. Pharmaceuticals. 2023; 16(1):129. https://doi.org/10.3390/ph16010129
Chicago/Turabian StyleMarcucci, Carolina, Marina Rademacher, Fabiola Kamecki, Valentina Pastore, Hernán Gerónimo Bach, Rafael Alejandro Ricco, Marcelo Luis Wagner, Damijan Knez, Stanislav Gobec, Natalia Colettis, and et al. 2023. "Biological Evaluation of Valeriana Extracts from Argentina with Potent Cholinesterase Inhibition for the Treatment of Neurodegenerative Disorders and Their Comorbidities—The Case of Valeriana carnosa Sm. (Caprifoliaceae) Studied in Mice" Pharmaceuticals 16, no. 1: 129. https://doi.org/10.3390/ph16010129