
From Snake Venoms to Therapeutics: A Focus on Natriuretic Peptides

Abstract
:1. Introduction
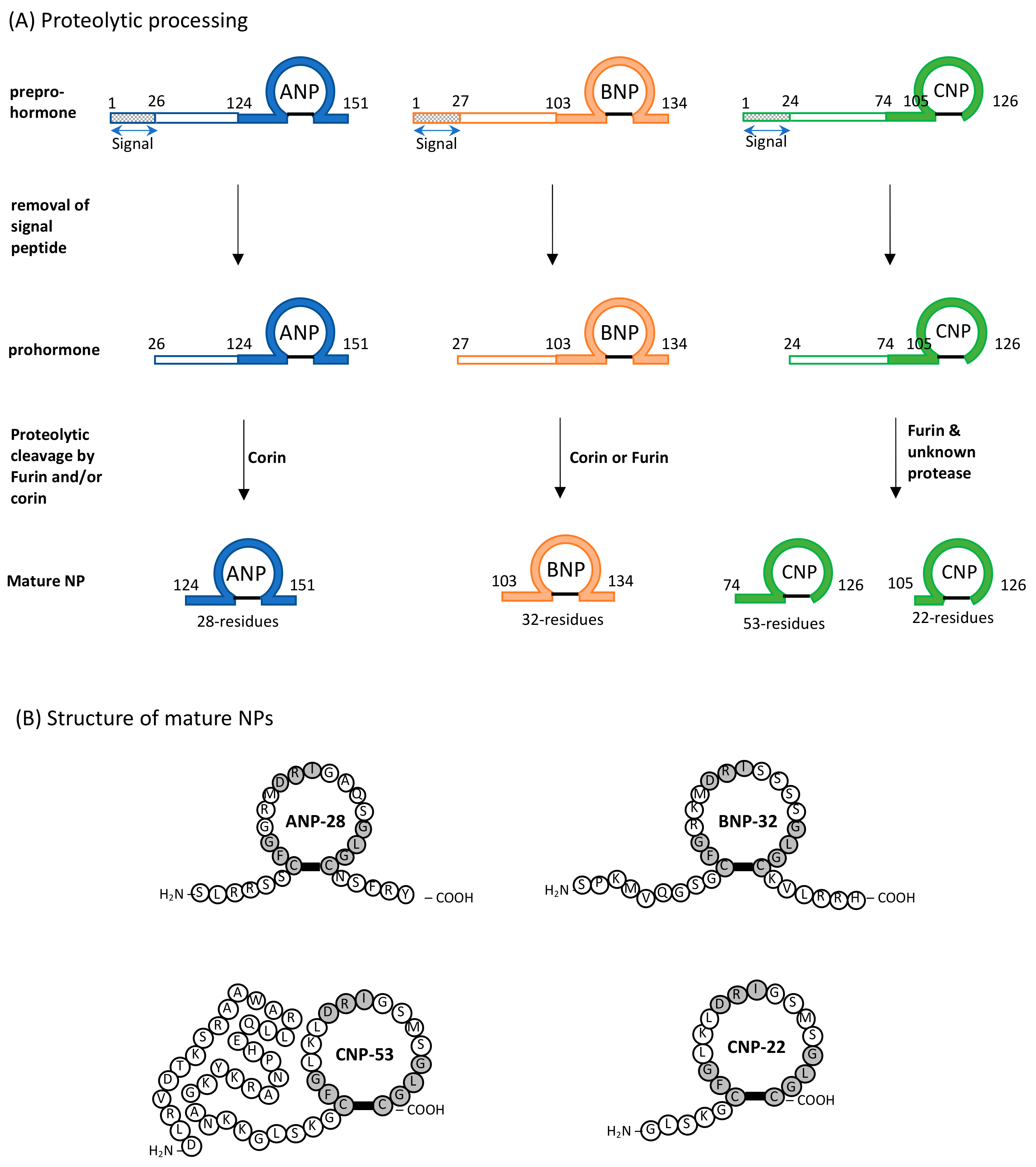
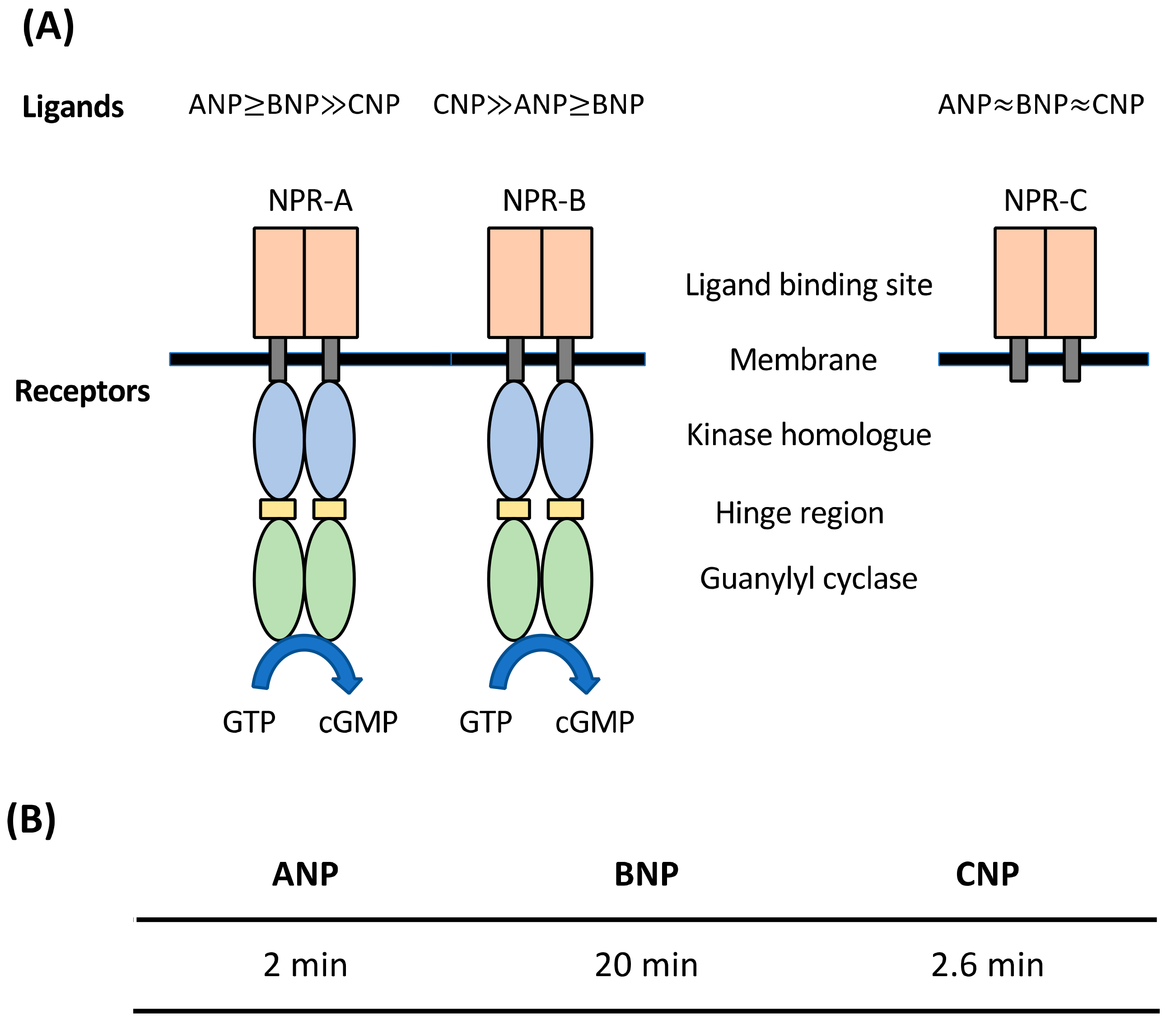
2. Natriuretic Peptides (NPs): General Considerations
3. Snake Venom Natriuretic Peptides (NPs)
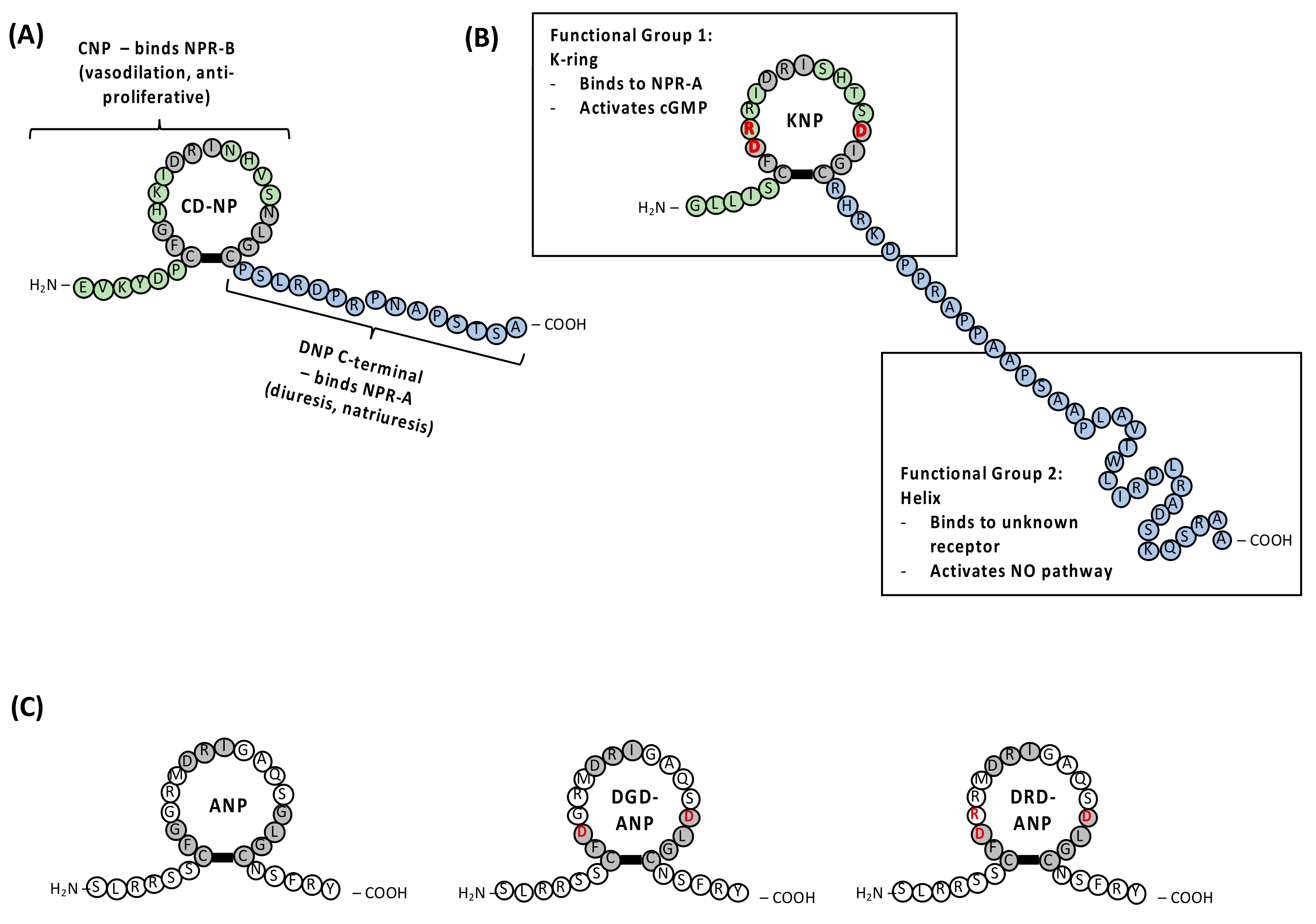
3.1. Dendroaspis NP (DNP) and Chimeric CD-NP
3.2. Krait NP (KNP), DGD-ANP and DRD-ANP
3.3. Other Snake NPs
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferraz, C.R.; Arrahman, A.; Xie, C.; Casewell, N.R.; Lewis, R.J.; Kool, J.; Cardoso, F.C. Multifunctional Toxins in Snake Venoms and Therapeutic Implications: From Pain to Hemorrhage and Necrosis. Front. Ecol. Evol. 2019, 7, 218. [Google Scholar] [CrossRef]
- Suranse, V.; Srikanthan, A.; Sunagar, K. Animal Venoms: Origin, Diversity and Evolution. In eLS; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2018; pp. 1–20. [Google Scholar]
- Fry, B.G.; Scheib, H.; van der Weerd, L.; Young, B.; McNaughtan, J.; Ramjan, S.R.; Vidal, N.; Poelmann, R.E.; Norman, J.A. Evolution of an arsenal: Structural and functional diversification of the venom system in the advanced snakes (Caenophidia). Mol. Cell. Proteom. 2008, 7, 215–246. [Google Scholar] [CrossRef] [PubMed]
- Dias, G.S.; Kitano, E.S.; Pagotto, A.H.; Sant’anna, S.V.S.; Rocha, M.M.; Zelanis, A.; Serrano, S.M. Individual variability in the venom proteome of juvenile Bothrops jararaca specimens. J. Proteome Res. 2013, 12, 4585–4598. [Google Scholar] [CrossRef] [PubMed]
- Menezes, M.C.; Furtado, M.F.; Travaglia-Cardoso, S.R.; Camargo, A.C.; Serrano, S.M. Sex-based individual variation of snake venom proteome among eighteen Bothrops jararaca siblings. Toxicon 2006, 47, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Barlow, A.; Pook, C.E.; Harrison, R.A.; Wüster, W. Coevolution of diet and prey-specific venom activity supports the role of selection in snake venom evolution. Proc. R. Soc. B Biol. Sci. 2009, 276, 2443–2449. [Google Scholar] [CrossRef] [PubMed]
- Casewell, N.R.; Jackson, T.N.W.; Laustsen, A.H.; Sunagar, K. Causes and Consequences of Snake Venom Variation. Trends Pharmacol. Sci. 2020, 41, 570–581. [Google Scholar] [CrossRef]
- Modahl, C.M.; Brahma, R.K.; Koh, C.Y.; Shioi, N.; Kini, R.M. Omics Technologies for Profiling Toxin Diversity and Evolution in Snake Venom: Impacts on the Discovery of Therapeutic and Diagnostic Agents. Annu. Rev. Anim. Biosci. 2020, 8, 91–116. [Google Scholar] [CrossRef]
- Koh, C.Y.; Kini, R.M. From snake venom toxins to therapeutics—Cardiovascular examples. Toxicon 2012, 59, 497–506. [Google Scholar] [CrossRef]
- de Bold, A.J.; Borenstein, H.B.; Veress, A.T.; Sonnenberg, H. A rapid and potent natriuretic response to intravenous injection of atrial myocardial extract in rats. Life Sci. 1981, 28, 89–94. [Google Scholar] [CrossRef]
- Potter, L.R.; Abbey-Hosch, S.; Dickey, D.M. Natriuretic peptides, their receptors, and cyclic guanosine monophosphate-dependent signaling functions. Endocr. Rev. 2006, 27, 47–72. [Google Scholar] [CrossRef]
- Rao, S.; Pena, C.; Shurmur, S.; Nugent, K. Atrial Natriuretic Peptide: Structure, Function, and Physiological Effects: A Narrative Review. Curr. Cardiol. Rev. 2021, 17, e051121191003. [Google Scholar] [CrossRef] [PubMed]
- Del Ry, S.; Cabiati, M.; Clerico, A. Recent advances on natriuretic peptide system: New promising therapeutic targets for the treatment of heart failure. Pharmacol. Res. 2013, 76, 190–198. [Google Scholar] [CrossRef]
- Hayek, S.; Nemer, M. Cardiac natriuretic peptides: From basic discovery to clinical practice. Cardiovasc. Ther. 2011, 29, 362–376. [Google Scholar] [CrossRef] [PubMed]
- Olins, G.M.; Patton, D.R.; Bovy, P.R.; Mehta, P.P. A linear analog of atrial natriuretic peptide (ANP) discriminates guanylate cyclase-coupled ANP receptors from non-coupled receptors. J. Biol. Chem. 1988, 263, 10989–10993. [Google Scholar] [CrossRef]
- Yoshihiro, O.; Hiroshi, I.; Yuka, Y. Molecular cloning and chromosomal assignment of the mouse C-type natriuretic peptide (CNP) gene (Nppc): Comparison with the human CNP gene (NPPC). Genomics 1994, 24, 383–387. [Google Scholar]
- Baba, M.; Yoshida, K.; Ieda, M. Clinical Applications of Natriuretic Peptides in Heart Failure and Atrial Fibrillation. Int. J. Mol. Sci. 2019, 20, 2824. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Wu, F.; Morser, J.; Wu, Q. Corin, a transmembrane cardiac serine protease, acts as a pro-atrial natriuretic peptide-converting enzyme. Proc. Natl. Acad. Sci. USA 2000, 97, 8525–8529. [Google Scholar] [CrossRef]
- Dietz, J.R. Mechanisms of atrial natriuretic peptide secretion from the atrium. Cardiovasc. Res. 2005, 68, 8–17. [Google Scholar] [CrossRef]
- McDonagh, T.A.; Metra, M.; Adamo, M.; Gardner, R.S.; Baumbach, A.; Böhm, M.; Burri, H.; Butler, J.; Čelutkienė, J.; Chioncel, O.; et al. 2021 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure. Eur. J. Heart Fail. 2022, 24, 4–131. [Google Scholar] [CrossRef]
- Tamura, N.; Ogawa, Y.; Chusho, H.; Nakamura, K.; Nakao, K.; Suda, M.; Kasahara, M.; Hashimoto, R.; Katsuura, G.; Mukoyama, M.; et al. Cardiac fibrosis in mice lacking brain natriuretic peptide. Proc. Natl. Acad. Sci. USA 2000, 97, 4239–4244. [Google Scholar] [CrossRef]
- Komatsu, Y.; Itoh, H.; Suga, S.; Ogawa, Y.; Hama, N.; Kishimoto, I.; Nakagawa, O.; Igaki, T.; Doi, K.; Yoshimasa, T.; et al. Regulation of endothelial production of C-type natriuretic peptide in coculture with vascular smooth muscle cells. Role of the vascular natriuretic peptide system in vascular growth inhibition. Circ. Res. 1996, 78, 606–614. [Google Scholar] [CrossRef]
- Potter, L.R. Guanylyl cyclase structure, function and regulation. Cell Signal 2011, 23, 1921–1926. [Google Scholar] [CrossRef] [PubMed]
- Garbers, D.L. Guanylyl cyclase-linked receptors. Pharmacol. Ther. 1991, 50, 337–345. [Google Scholar] [CrossRef]
- Suga, S.; Nakao, K.; Hosoda, K.; Mukoyama, M.; Ogawa, Y.; Shirakami, G.; Arai, H.; Saito, Y.; Kambayashi, Y.; Inouye, K.; et al. Receptor selectivity of natriuretic peptide family, atrial natriuretic peptide, brain natriuretic peptide, and C-type natriuretic peptide. Endocrinology 1992, 130, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Matsukawa, N.; Grzesik, W.J.; Takahashi, N.; Pandey, K.N.; Pang, S.; Yamauchi, M.; Smithies, O. The natriuretic peptide clearance receptor locally modulates the physiological effects of the natriuretic peptide system. Proc. Natl. Acad. Sci. USA 1999, 96, 7403–7408. [Google Scholar] [CrossRef]
- Abassi, Z.A.; Golomb, E.; Agbaria, R.; Roller, P.P.; Tate, J.; Keiser, H.R. Hydrolysis of iodine labelled urodilatin and ANP by recombinant neutral endopeptidase EC. 3.4.24.11. Br. J. Pharmacol. 1994, 113, 204–208. [Google Scholar] [CrossRef]
- Stephenson, S.L.; Kenny, A.J. The hydrolysis of α-human atrial natriuretic peptide by pig kidney microvillar membranes is initiated by endopeptidase-24.11. Biochem. J. 1987, 243, 183–187. [Google Scholar] [CrossRef]
- Nakao, K.; Sugawara, A.; Morii, N.; Sakamoto, M.; Yamada, T.; Itoh, H.; Shiono, S.; Saito, Y.; Nishimura, K.; Ban, T.; et al. The pharmacokinetics of alpha-human atrial natriuretic polypeptide in healthy subjects. Eur. J. Clin. Pharmacol. 1986, 31, 101–103. [Google Scholar] [CrossRef]
- Hunt, P.; Richards, A.; Espiner, E.; Nicholls, M.; Yandle, T. Bioactivity and metabolism of C-type natriuretic peptide in normal man. J. Clin. Endocrinol. Metab. 1994, 78, 1428–1435. [Google Scholar]
- Abassi, Z.; Karram, T.; Ellaham, S.; Winaver, J.; Hoffman, A. Implications of the natriuretic peptide system in the pathogenesis of heart failure: Diagnostic and therapeutic importance. Pharmacol. Ther. 2004, 102, 223–241. [Google Scholar] [CrossRef]
- Oliveira, A.L.; Viegas, M.F.; da Silva, S.L.; Soares, A.M.; Ramos, M.J.; Fernandes, P.A. The chemistry of snake venom and its medicinal potential. Nat. Rev. Chem. 2022, 6, 451–469. [Google Scholar] [CrossRef] [PubMed]
- Schweitz, H.; Vigne, P.; Moinier, D.; Frelin, C.; Lazdunski, M. A new member of the natriuretic peptide family is present in the venom of the green mamba (Dendroaspis angusticeps). J. Biol. Chem. 1992, 267, 13928–13932. [Google Scholar] [CrossRef]
- Singh, G.; Maguire, J.J.; Kuc, R.E.; Skepper, J.N.; Fidock, M.; Davenport, A.P. Characterization of the snake venom ligand [125I]-DNP binding to natriuretic peptide receptor-A in human artery and potent DNP mediated vasodilatation. Br. J. Pharmacol. 2006, 149, 838–844. [Google Scholar] [CrossRef]
- Lisy, O.; Jougasaki, M.; Heublein, D.M.; Schirger, J.A.; Chen, H.H.; Wennberg, P.W.; Burnett, J.C. Renal actions of synthetic Dendroaspis natriuretic peptide. Kidney Int. 1999, 56, 502–508. [Google Scholar] [CrossRef]
- Schirger, J.A.; Heublein, D.M.; Chen, H.H.; Lisy, O.; Jougasaki, M.; Wennberg, P.W.; Burnett, J.C. Presence of Dendroaspis Natriuretic Peptide-Like Immunoreactivity in Human Plasma and Its Increase During Human Heart Failure. Mayo Clin. Proc. 1999, 74, 126–130. [Google Scholar] [CrossRef]
- Best, P.J.M.; Burnett, J.C., Jr.; Wilson, S.H.; Holmes, D.R., Jr.; Lerman, A. Dendroaspis natriuretic peptide relaxes isolated human arteries and veins. Cardiovasc. Res. 2002, 55, 375–384. [Google Scholar] [CrossRef]
- Lisy, O.; Lainchbury, J.G.; Leskinen, H.; Burnett, J.C. Therapeutic Actions of a New Synthetic Vasoactive and Natriuretic Peptide, Dendroaspis Natriuretic Peptide, in Experimental Severe Congestive Heart Failure. Hypertension 2001, 37, 1089–1094. [Google Scholar] [CrossRef]
- Ichiki, T.; Dzhoyashvili, N.; Burnett, J.C., Jr. Natriuretic peptide based therapeutics for heart failure: Cenderitide: A novel first-in-class designer natriuretic peptide. Int. J. Cardiol. 2019, 281, 166–171. [Google Scholar] [CrossRef]
- Dickey, D.M.; Burnett, J.C., Jr.; Potter, L.R. Novel Bifunctional Natriuretic Peptides as Potential Therapeutics. J. Biol. Chem. 2008, 283, 35003–35009. [Google Scholar] [CrossRef]
- Kawakami, R.; Lee, C.Y.W.; Scott, C.; Bailey, K.R.; Schirger, J.A.; Chen, H.H.; Benike, S.L.; Cannone, V.; Martin, F.L.; Sangaralingham, S.J.; et al. A Human Study to Evaluate Safety, Tolerability, and Cyclic GMP Activating Properties of Cenderitide in Subjects With Stable Chronic Heart Failure. Clin. Pharmacol. Ther. 2018, 104, 546–552. [Google Scholar] [CrossRef]
- Siang, A.S.; Doley, R.; Vonk, F.J.; Kini, R.M. Transcriptomic analysis of the venom gland of the red-headed krait (Bungarus flaviceps) using expressed sequence tags. BMC Mol. Biol. 2010, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Sridharan, S.; Kini, R.M. Tail wags the dog: Activity of krait natriuretic peptide is determined by its C-terminal tail in a natriuretic peptide receptor-independent manner. Biochem. J. 2015, 469, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Sridharan, S.; Kini, R.M. Decoding the molecular switches of natriuretic peptides which differentiate its vascular and renal functions. Biochem. J. 2018, 475, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Rademaker, M.T.; Scott, N.J.A.; Koh, C.Y.; Kini, R.M.; Richards, A.M. Natriuretic peptide analogues with distinct vasodilatory or renal activity: Integrated effects in health and experimental heart failure. Cardiovasc. Res. 2020, 117, 508–519. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Wickramaratana, J.C.; Lemme, S.; Beuve, A.; Garbers, D.; Hodgson, W.C.; Alewood, P. Novel natriuretic peptides from the venom of the inland taipan (Oxyuranus microlepidotus): Isolation, chemical and biological characterisation. Biochem. Biophys. Res. Commun. 2005, 327, 1011–1015. [Google Scholar] [CrossRef]
- Pierre, L.S.; Flight, S.; Masci, P.P.; Hanchard, K.J.; Lewis, R.J.; Alewood, P.F.; Jersey, J.D.; Lavin, M.F. Cloning and characterisation of natriuretic peptides from the venom glands of Australian elapids. Biochimie 2006, 88, 1923–1931. [Google Scholar] [CrossRef]
- Bazaa, A.; Marrakchi, N.; El Ayeb, M.; Sanz, L.; Calvete, J.J. Snake venomics: Comparative analysis of the venom proteomes of the Tunisian snakes Cerastes cerastes, Cerastes vipera and Macrovipera lebetina. Proteomics 2005, 5, 4223–4235. [Google Scholar] [CrossRef]
- Amininasab, M.; Elmi, M.M.; Endlich, N.; Endlich, K.; Parekh, N.; Naderi-Manesh, H.; Schaller, J.; Mostafavi, H.; Sattler, M.; Sarbolouki, M.N.; et al. Functional and structural characterization of a novel member of the natriuretic family of peptides from the venom of Pseudocerastes persicus. FEBS Lett. 2004, 557, 104–108. [Google Scholar] [CrossRef]
- Tourki, B.; Dumesnil, A.; Belaidi, E.; Ghrir, S.; Godin-Ribuot, D.; Marrakchi, N.; Richard, V.; Mulder, P.; Messadi, E. Lebetin 2, a Snake Venom-Derived B-Type Natriuretic Peptide, Provides Immediate and Prolonged Protection against Myocardial Ischemia-Reperfusion Injury via Modulation of Post-Ischemic Inflammatory Response. Toxins 2019, 11, 524. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| NPs | Snakes | Functions | Applications |
|---|---|---|---|
| DNP | Green mamba (Dendroaspis angusticeps) | Vasodilation, natriuresis, diuresis | C-terminal tail creates CD-NP with CNP |
| KNP | Red-headed krait (Bungarus flaviceps) | Vasodilation | Create DGD-ANP and DRD-ANP |
| TNP-a, TNP-b and TNP-c | Taipan snake (Oxyranus microlepidotus) | Vasodilation (TNP-c) | - |
| Pt-NP-a, Pt-NP-c | Eastern brown snake (Pseudonaja textilis) King brown snake (Pseudechis australis) | Inhibit ACE 1 | - |
| TcNP-a | Rough scaled snake (Tropidechis carinatus) | Activate both NPR-A and NPR-B | - |
| Lebetin 1 and 2 | Blunt-nosed viper (Macrovipera lebetina) | Anti-platelet, reduce inflammation | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ang, W.F.; Koh, C.Y.; Kini, R.M. From Snake Venoms to Therapeutics: A Focus on Natriuretic Peptides. Pharmaceuticals 2022, 15, 1153. https://doi.org/10.3390/ph15091153
Ang WF, Koh CY, Kini RM. From Snake Venoms to Therapeutics: A Focus on Natriuretic Peptides. Pharmaceuticals. 2022; 15(9):1153. https://doi.org/10.3390/ph15091153
Chicago/Turabian StyleAng, Wei Fong, Cho Yeow Koh, and R. Manjunatha Kini. 2022. "From Snake Venoms to Therapeutics: A Focus on Natriuretic Peptides" Pharmaceuticals 15, no. 9: 1153. https://doi.org/10.3390/ph15091153