
Modulation of Intestinal Corticotropin-Releasing Hormone Signaling by the Herbal Preparation STW 5-II: Possible Mechanisms for Irritable Bowel Syndrome Management




Abstract
:1. Introduction
2. Results
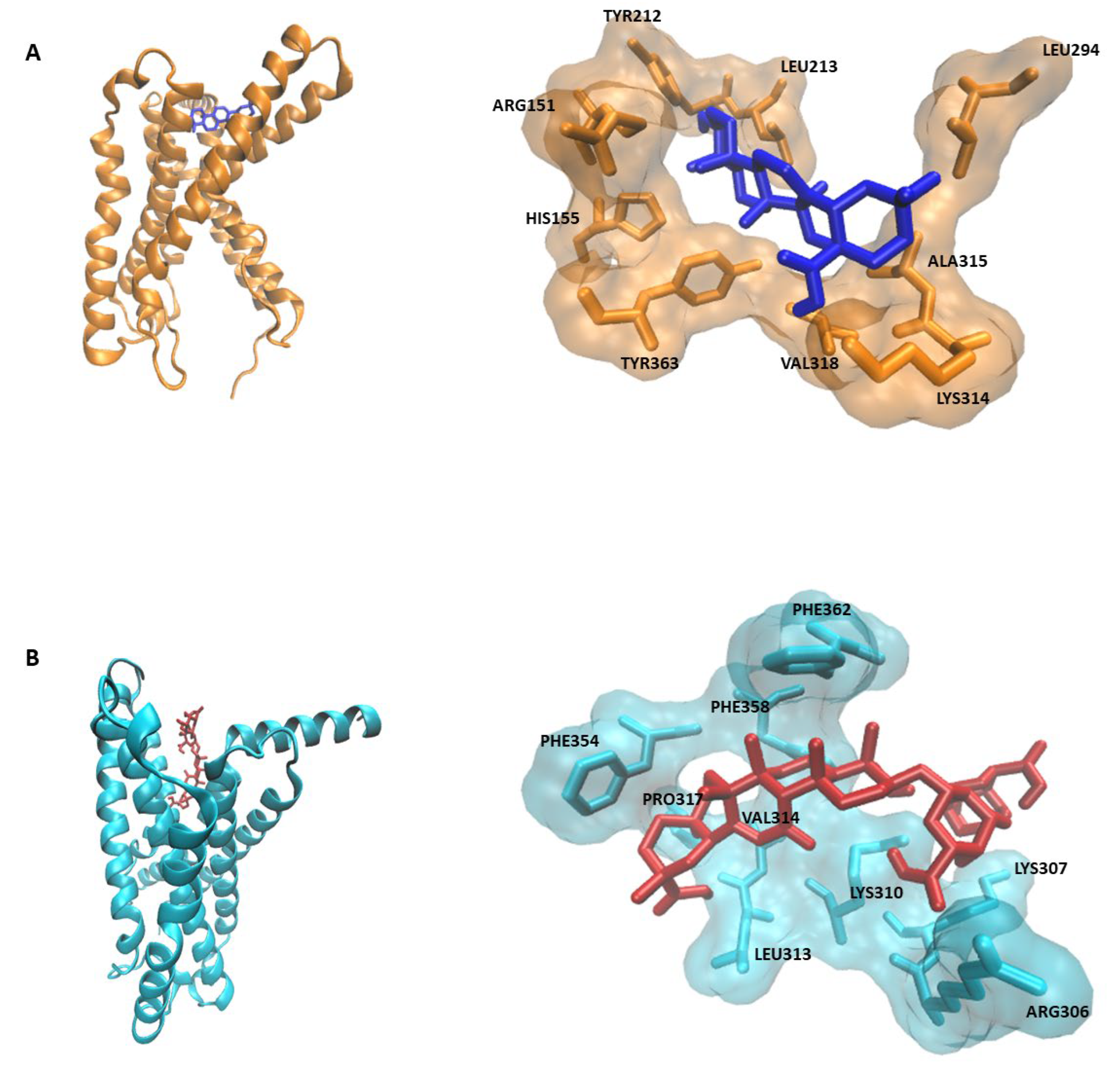
2.1. PyRx Screening and Docking
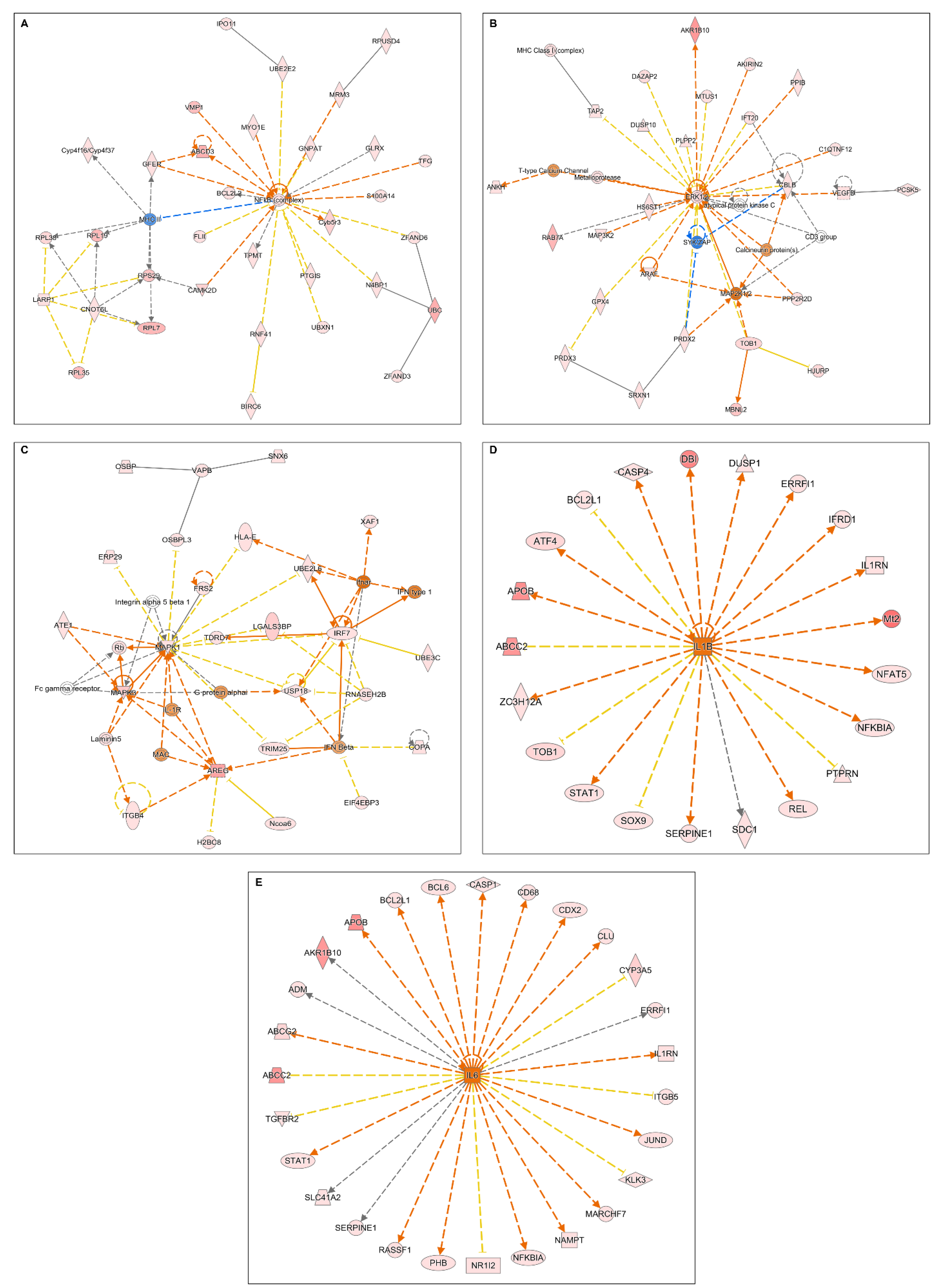
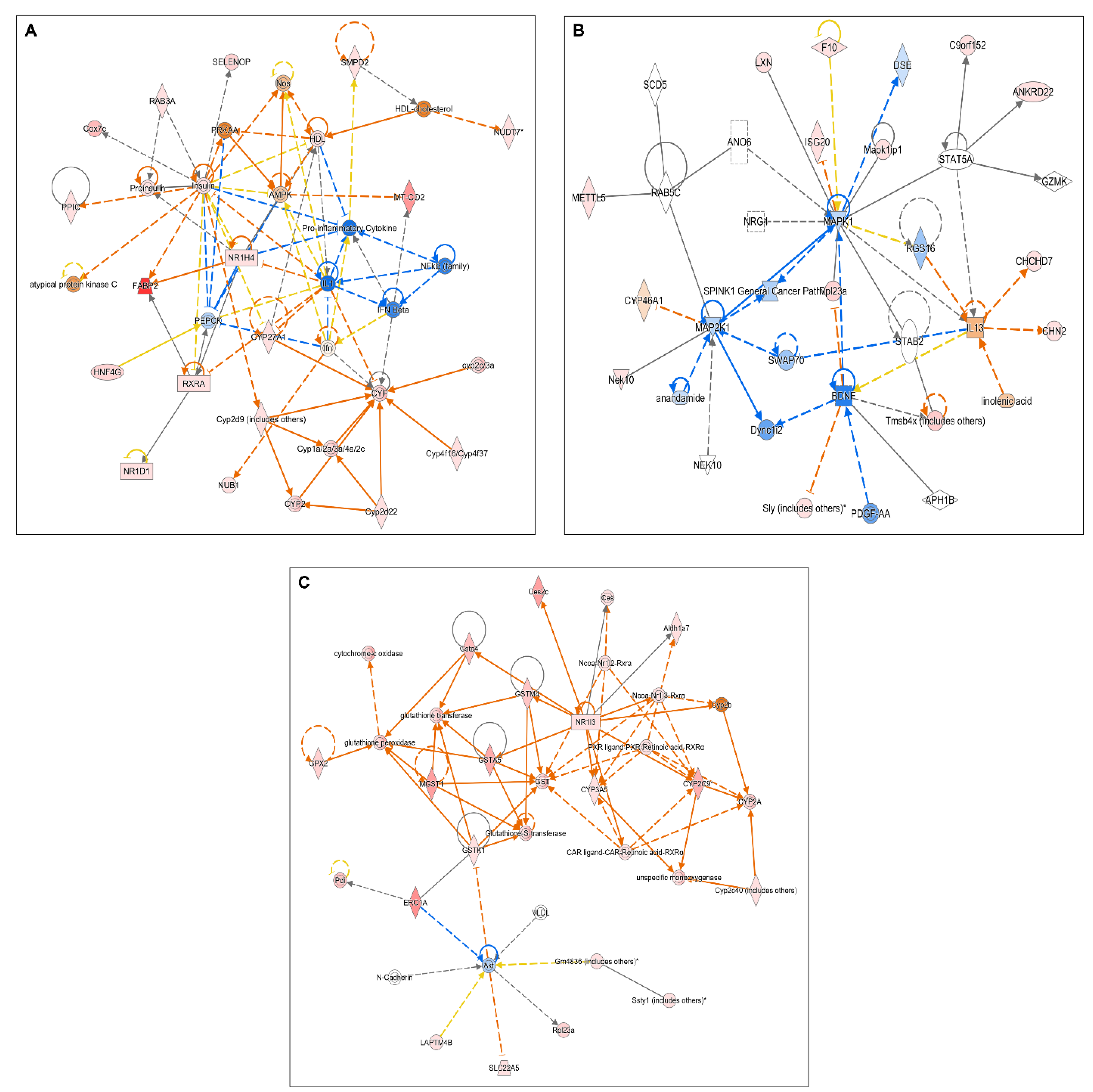
2.2. Microarray Analyses
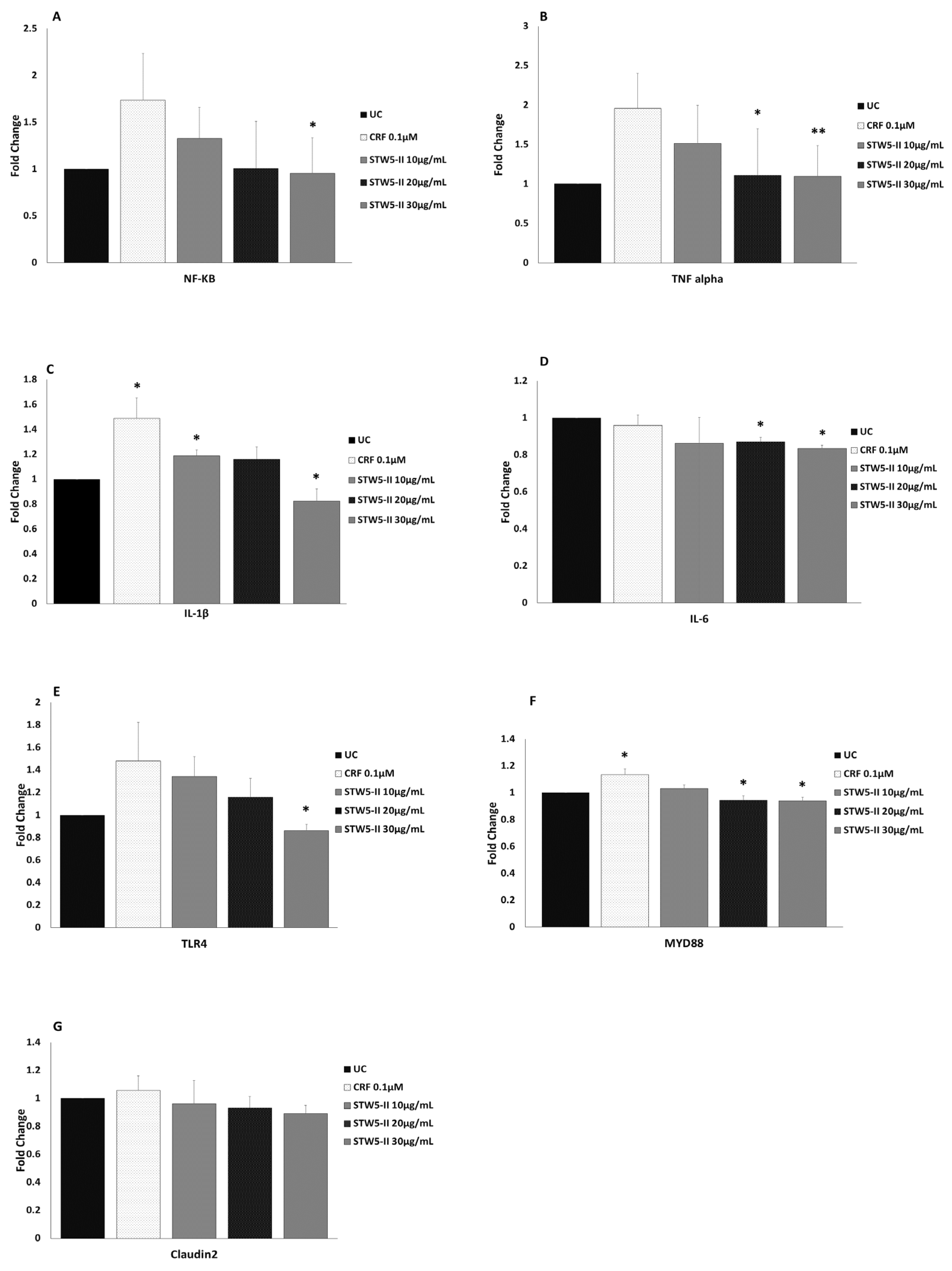
2.3. Quantitative Real-Time PCR (qPCR)
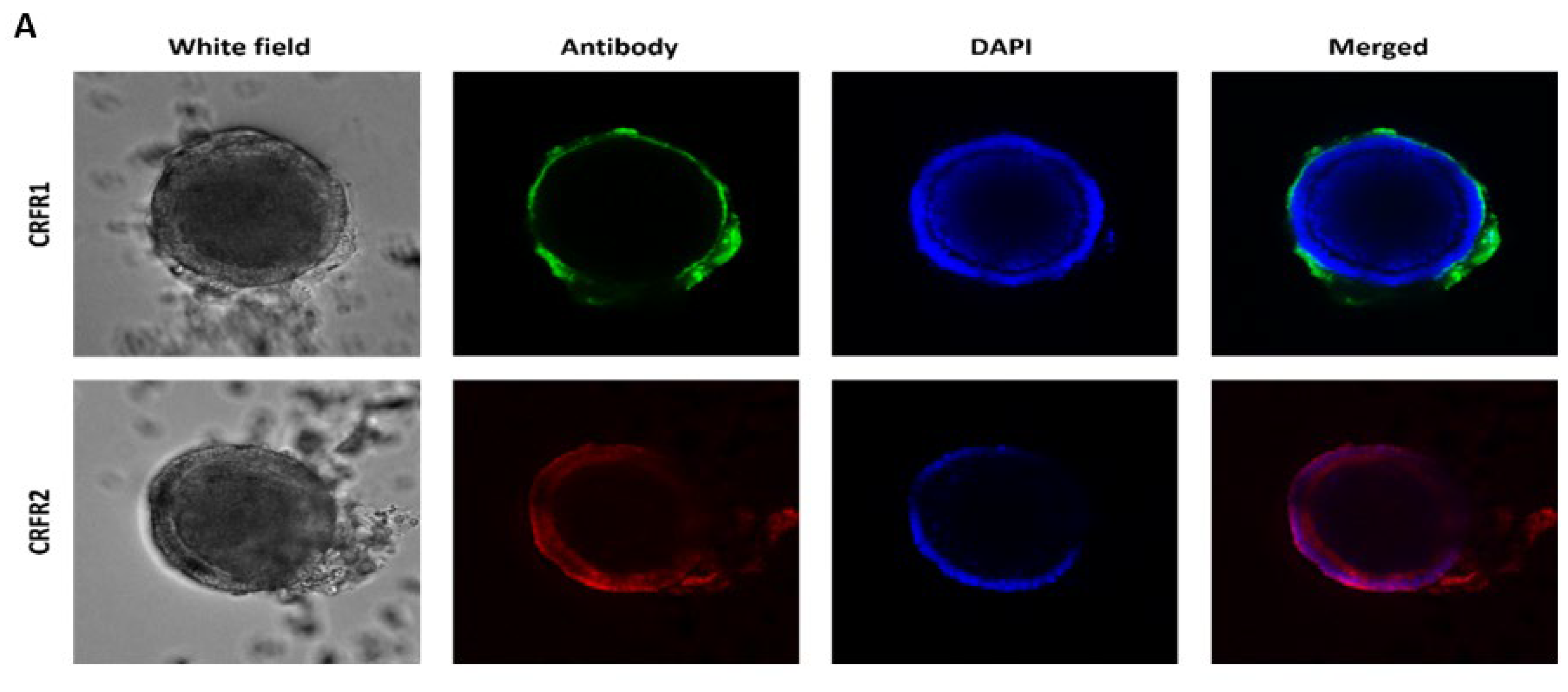
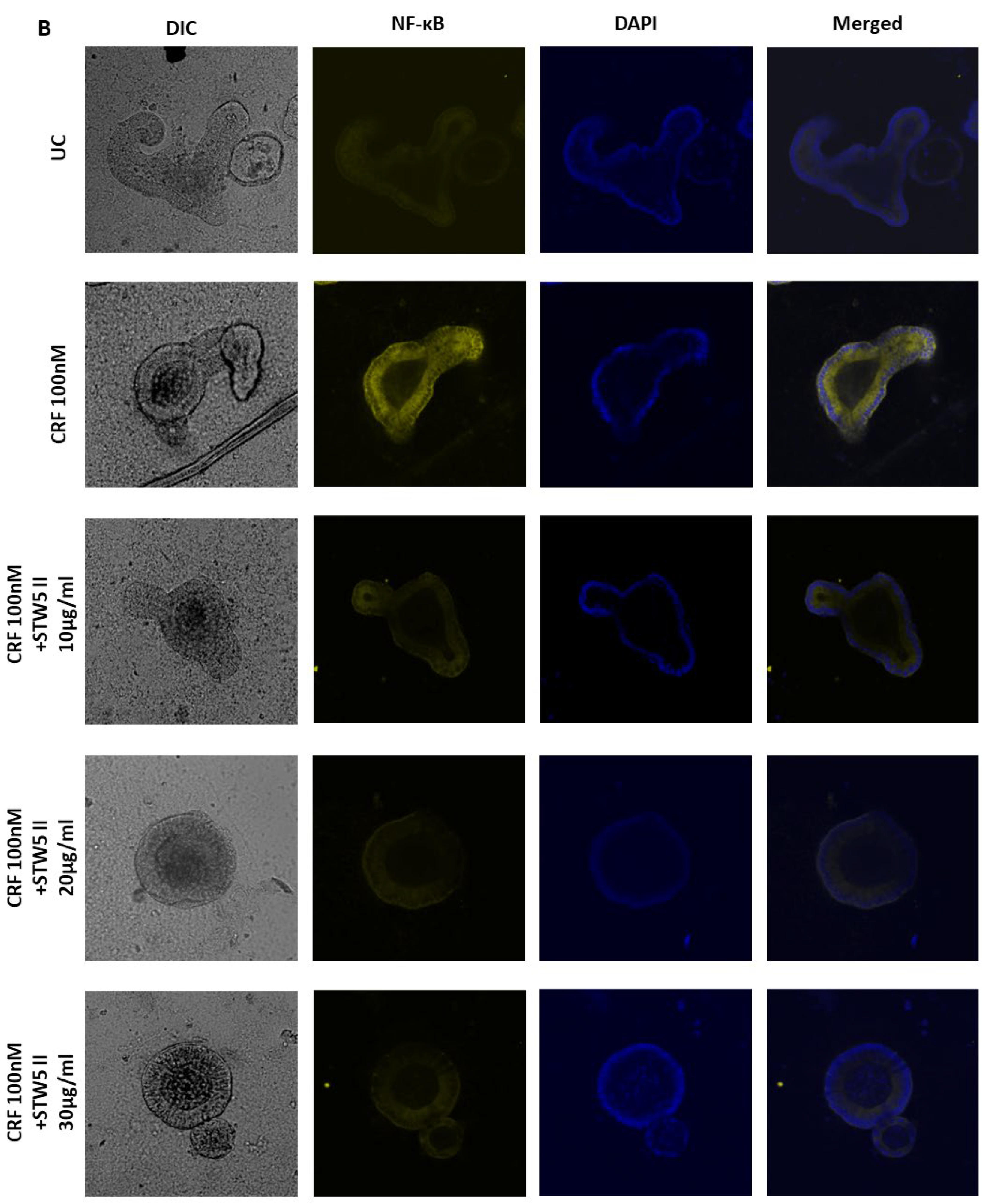
2.4. Whole-Mount Immunofluorescence Staining
2.5. Detection of Serotonin by ELISA
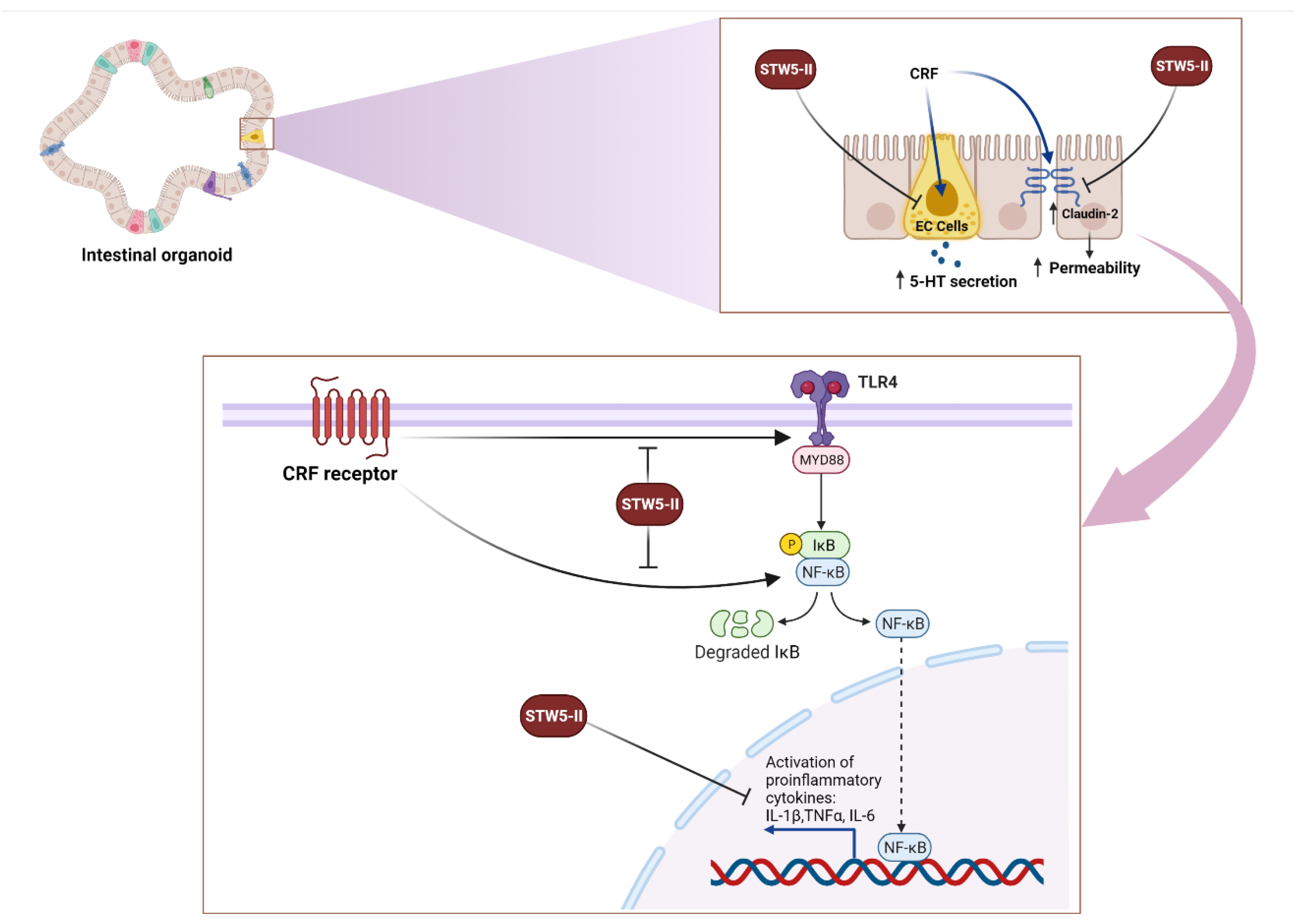
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Mouse Intestinal Organoid Culture
4.3. Stress-Induced IBS-Like Model
4.4. PyRx Screening
4.5. Microarray Analyses
4.6. Molecular Docking
4.7. Quantitative Real-Time Reverse Transcription PCR
4.8. Whole-Mount Immunofluorescent Staining
4.9. Detection of Serotonin by ELISA
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lacy, B.E.; Mearin, F.; Chang, L.; Chey, W.D.; Lembo, A.J.; Simren, M.; Spiller, R. Bowel Disorders. Gastroenterology 2016, 150, 1393–1407. [Google Scholar] [CrossRef] [PubMed]
- Drossman, D.A.; Hasler, W.L. Rome IV—Functional GI Disorders: Disorders of Gut-Brain Interaction. Gastroenterology 2016, 150, 1257–1261. [Google Scholar] [CrossRef] [PubMed]
- Tsang, S.W.; Auyeung, K.K.W.; Bian, Z.X.; Ko, J.K.S. Pathogenesis, Experimental Models and Contemporary Pharmacotherapy of Irritable Bowel Syndrome: Story About the Brain-Gut Axis. Curr. Neuropharmacol. 2016, 14, 842–856. [Google Scholar] [CrossRef] [PubMed]
- Chong, P.P.; Chin, V.K.; Looi, C.Y.; Wong, W.F.; Madhavan, P.; Yong, V.C. The Microbiome and Irritable Bowel Syndrome—A Review on the Pathophysiology, Current Research and Future Therapy. Front. Microbiol. 2019, 10, 1136. [Google Scholar] [CrossRef]
- Qin, H.Y.; Cheng, C.W.; Tang, X.D.; Bian, Z.X. Impact of Psychological Stress on Irritable Bowel Syndrome. World J. Gastroenterol. 2014, 20, 14126–14131. [Google Scholar] [CrossRef]
- Mahurkar-Joshi, S.; Chang, L. Epigenetic Mechanisms in Irritable Bowel Syndrome. Front. Psychiatry 2020, 11, 805. [Google Scholar] [CrossRef]
- Stengel, A.; Taché, Y. Corticotropin-Releasing Factor Signaling and Visceral Response to Stress. Exp. Biol. Med. 2010, 235, 1168–1178. [Google Scholar] [CrossRef]
- Buckinx, R.; Adriaensen, D.; Van Nassauw, L.; Timmermans, J.P. Corticotrophin-Releasing Factor, Related Peptides, and Receptors in the Normal and Inflamed Gastrointestinal Tract. Front. Neurosci. 2011, 5, 54. [Google Scholar] [CrossRef]
- Larauche, M.; Kiank, C.; Tache, Y. Corticotropin Releasing Factor Signaling in Colon and Ileum: Regulation by Stress and Pathophysiological Implications. J. Physiol. Pharmacol. 2009, 60 (Suppl. S7), 33–46. [Google Scholar]
- Li, B.; Lee, C.; Filler, T.; Hock, A.; Wu, R.Y.; Li, Q.; Chen, S.; Koike, Y.; Ip, W.; Chi, L.; et al. Inhibition of Corticotropin-Releasing Hormone Receptor 1 and Activation of Receptor 2 Protect against Colonic Injury and Promote Epithelium Repair. Sci. Rep. 2017, 7, 1–11. [Google Scholar]
- Nozu, T.; Miyagishi, S.; Nozu, R.; Takakusaki, K.; Okumura, T. Altered Colonic Sensory and Barrier Functions by CRF: Roles of TLR4 and IL-1. J. Endocrinol. 2018, 239, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.; Efferth, T.; Abdel-Aziz, H. Anti-Inflammatory Effects of Herbal Preparations STW5 and STW5-II in Cytokine-Challenged Normal Human Colon Cells. Front. Pharmacol. 2016, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rösch, W.; Vinson, B.; Sassin, I. A Randomised Clinical Trial Comparing the Efficacy of a Herbal Preparation STW 5 with the Prokinetic Drug Cisapride in Patients with Dysmotility Type of Functional Dyspepsia. Z. Gastroenterol. 2002, 40, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Madisch, A.; Holtmann, G.; Mayr, G.; Vinson, B.; Hotz, J. Treatment of Functional Dyspepsia with a Herbal Preparation: A Double-Blind, Randomized, Placebo-Controlled, Multicenter Trial. Digestion 2004, 69, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Elbadawi, M.; Ammar, R.M.; Aziz-Kalbhenn, H.; Rabini, S.; Klauck, S.M.; Dawood, M.; Saeed, M.E.M.; Kampf, C.J.; Efferth, T. Anti-Inflammatory and Tight Junction Protective Activity of the Herbal Preparation STW 5-II on Mouse Intestinal Organoids. Phytomedicine 2021, 88, 153589. [Google Scholar] [CrossRef]
- Chatoo, M.; Li, Y.; Ma, Z.; Coote, J.; Du, J.; Chen, X. Involvement of Corticotropin-Releasing Factor and Receptors in Immune Cells in Irritable Bowel Syndrome. Front. Endocrinol. 2018, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- La Fleur, S.E.; Wick, E.C.; Idumalla, P.S.; Grady, E.F.; Bhargava, A. Role of Peripheral Corticotropin-Releasing Factor and Urocortin II in Intestinal Inflammation and Motility in Terminal Ileum. Proc. Natl. Acad. Sci. USA 2005, 102, 7647–7652. [Google Scholar] [CrossRef]
- Merker, S.R.; Weitz, J.; Stange, D.E. Gastrointestinal Organoids: How They Gut It Out. Dev. Biol. 2016, 420, 239–250. [Google Scholar] [CrossRef]
- Tache, Y.; Larauche, M.; Yuan, P.-Q.; Million, M. Brain and Gut CRF Signaling: Biological Actions and Role in the Gastrointestinal Tract. Curr. Mol. Pharmacol. 2018, 11, 51–71. [Google Scholar] [CrossRef]
- Moss, A.C.; Anton, P.; Savidge, T.; Newman, P.; Cheifelz, A.S.; Gay, J.; Paraschos, S.; Winter, M.W.; Moyer, M.P.; Karalis, K.; et al. Urocortin II Mediates Pro-Inflammatory Effects in Human Colonocytes via Corticotropin-Releasing Hormone Receptor. Gut 2007, 56, 1210–1217. [Google Scholar] [CrossRef]
- He, X.; Cui, L.-H.; Wang, X.-H.; Yan, Z.-H.; Li, C.; Gong, S.-D.; Zheng, Y.; Luo, Z.; Wang, Y. Modulation of Inflammation by Toll-like Receptor 4/Nuclear Factor-Kappa B in Diarrhea-Predominant Irritable Bowel Syndrome. Oncotarget 2017, 8, 113957–113965. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Shi, H.; Liu, Y.; Ren, X.; He, S.; Chang, X.; Yin, Y. Activation of Corticotropin-Releasing Factor Receptor 1 Aggravates Dextran Sodium Sulphate-Induced Colitis in Mice by Promoting M1 Macrophage Polarization. Mol. Med. Rep. 2018, 17, 234–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Fandiño, O.; Hernández-Ruiz, J.; Schmulson, M. From Cytokines to Toll-Like Receptors and Beyond—Current Knowledge and Future Research Needs in Irritable Bowel Syndrome. J. Neurogastroenterol. Motil. 2010, 16, 363–373. [Google Scholar] [CrossRef]
- Kiank, C.; Taché, Y.; Larauche, M. Stress-Related Modulation of Inflammation in Experimental Models of Bowel Disease and Post-Infectious Irritable Bowel Syndrome: Role of Corticotropin Releasing Factor Receptors. Brain Behav. Immun. 2010, 24, 41–48. [Google Scholar] [CrossRef]
- Uh, A.; Simmons, C.F.; Bresee, C.; Khoury, N.; Gombart, A.F.; Nicholson, R.C.; Kocak, H.; Equils, O. MyD88 and TRIF Mediate the Cyclic Adenosine Monophosphate (CAMP). Induced Corticotropin Releasing Hormone (CRH) Expression in JEG3 Choriocarcinoma Cell Line. Reprod. Biol. Endocrinol. 2009, 7, 1–7. [Google Scholar] [CrossRef]
- Piche, T. Tight Junctions and IBS—The Link between Epithelial Permeability, Low-Grade Inflammation, and Symptom Generation? Neurogastroenterol. Motil. 2014, 26, 296–302. [Google Scholar] [CrossRef]
- Martínez, C.; Vicario, M.; Ramos, L.; Lobo, B.; Mosquera, J.L.; Alonso, C.; Sánchez, A.; Guilarte, M.; Antolín, M.; De Torres, I.; et al. The Jejunum of Diarrhea-Predominant Irritable Bowel Syndrome Shows Molecular Alterations in the Tight Junction Signaling Pathway That Are Associated with Mucosal Pathobiology and Clinical Manifestations. Am. J. Gastroenterol. 2012, 107, 736–746. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, Z.Q.; Liu, X.Y.; Yang, L.; Geng, X.R.; Yang, G.; Liu, Z.G.; Zheng, P.Y.; Yang, P.C. Stress-Derived Corticotropin Releasing Factor Breaches Epithelial Endotoxin Tolerance. PLoS ONE 2013, 8, e65760. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Bin, L.; Chaoying, C.; Meng, Z.; Meng, L.; Xi, W. Potential Regulatory Effects of Corticotropin-Releasing Factor on Tight Junction-Related Intestinal Epithelial Permeability Are Partially Mediated by CK8 Upregulation. Cell. Physiol. Biochem. 2017, 44, 1161–1173. [Google Scholar] [CrossRef]
- Garvin, B.; Wiley, J.W. The Role of Serotonin in Irritable Bowel Syndrome: Implications for Management. Curr. Gastroenterol. Rep. 2008, 10, 363–368. [Google Scholar] [CrossRef]
- Panda, B. Happiness through the Eye of a Chemist Disorders. Int. J. HIT Trans. ECCN 2018, 4, 15–19. [Google Scholar]
- Gershon, M.D.; Tack, J. The Serotonin Signaling System: From Basic Understanding To Drug Development for Functional GI Disorders. Gastroenterology 2007, 132, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Costedio, M.M.; Hyman, N.; Mawe, G.M. Serotonin and Its Role in Colonic Function and in Gastrointestinal Disorders. Dis. Colon. Rectum. 2007, 50, 376–388. [Google Scholar] [CrossRef]
- Crowell, M.D. Role of Serotonin in the Pathophysiology of the Irritable Bowel Syndrome. Brit. J. Pharmacol. 2004, 141, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Hadjivasilis, A.; Tsioutis, C.; Michalinos, A.; Ntourakis, D. New Insights into Irritable Bowel Syndrome: From Pathophysiology to Treatment. Ann. Gastroenterol. 2019, 32, 1–11. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand | Binding Energy kcal/mol |
|---|---|
| Oleanolic acid | −8.9 |
| Licorice glucoside C1 | −8.9 |
| Licorice saponin E2 | −8.8 |
| Yunganoside L2 | −8.6 |
| Glycyrrhetinic acid | −8.4 |
| Rosmarinic acid dimer | −8.4 |
| Glycyrrhizinic acid | −8.3 |
| Licorice saponin G2 | −8.3 |
| Licorice saponin B2 | −8.2 |
| Licorice saponin B2 isomer | −8.2 |
| Ligand | Binding Energy kcal/mol |
|---|---|
| Licorice saponin G2 | −9.2 |
| Yunganoside L2 | −9.2 |
| Glycyrrhetinic acid | −9.1 |
| Oleanolic acid | −9.0 |
| Licorice saponin B2 isomer | −9.0 |
| Licorice saponin_B2 | −8.7 |
| Rosmarinic acid dimer | −8.6 |
| Cucurbitacin I | −8.5 |
| Macedonoside-A-acetate | −8.5 |
| Kaempferol-3,3″′,4′-tri-O-glucoside-7-O-rhamnoside | −8.5 |
| Ligand | Target | Binding Energy kcal/mol | pKi (nM) | Amino Acids |
|---|---|---|---|---|
| Oleanolic acid | CRFR1 | −8.6 ± 0.01 | 496.6 ± 9.1 | LEU294, LEU213, TYR212, ALA315, LYS314, VAL318, TYR363, ARG151, HIS155 |
| Licorice saponin G2 | CRFR2 | −10.07 ± ≤0.001 | 41.36 ± ≤0.001 | LYS307, ARG306, LYS310, LEU313, VAL314, PRO317, PHE354, PHE358, PHE362 |
| Gene | Gene Symbol | Forward Primer | Reverse Primer |
|---|---|---|---|
| Glyceraldehyde-3-phosphate dehydrogenase | GAPDH | GAGGGATGCTGCCCTTACC | CAAATCCGTTCACACCGACC |
| Tumor necrosis factor-alpha | TNFα | TAGCCCACGTCGTAGCAAAC | ACAAGGTACAACCCATCGGC |
| Nuclear factor kappa B Subunit 1 | NF-κB | CTCTGGCGAATGGCTTTACT | GAGGGGAAACAGATCGTCCA |
| Interleukin-1 beta | IL-1β | TGCCACCTTTTGACAGTGATG | AAAGGTTTGGAAGCAGCCCT |
| Interleukin-6 | IL-6 | ACCAAGAGATAAGCTGGAGTCA | TAACGCACTAGGTTTGCCGA |
| Claudin-2 | CLDN2 | ATGCCTTCTTGAGCCTGCTT | GCTGCTGCTCTTGCTTCTTG |
| Myeloid differentiation Primary response protein 88 | MyD88 | AGAGCTGCTGGCCTTGTTAG | GACTCCTGGTTCTGCTGCTT |
| Toll-like receptor 4 | TLR4 | TTCTTCTCCTGCCTGACACC | GTCATCAGGGACTTTGCTGAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elbadawi, M.; Ammar, R.M.; Rabini, S.; Klauck, S.M.; Efferth, T. Modulation of Intestinal Corticotropin-Releasing Hormone Signaling by the Herbal Preparation STW 5-II: Possible Mechanisms for Irritable Bowel Syndrome Management. Pharmaceuticals 2022, 15, 1121. https://doi.org/10.3390/ph15091121
Elbadawi M, Ammar RM, Rabini S, Klauck SM, Efferth T. Modulation of Intestinal Corticotropin-Releasing Hormone Signaling by the Herbal Preparation STW 5-II: Possible Mechanisms for Irritable Bowel Syndrome Management. Pharmaceuticals. 2022; 15(9):1121. https://doi.org/10.3390/ph15091121
Chicago/Turabian StyleElbadawi, Mohamed, Ramy M. Ammar, Sabine Rabini, Sabine M. Klauck, and Thomas Efferth. 2022. "Modulation of Intestinal Corticotropin-Releasing Hormone Signaling by the Herbal Preparation STW 5-II: Possible Mechanisms for Irritable Bowel Syndrome Management" Pharmaceuticals 15, no. 9: 1121. https://doi.org/10.3390/ph15091121