
Galangin Exhibits Neuroprotective Effects in 6-OHDA-Induced Models of Parkinson’s Disease via the Nrf2/Keap1 Pathway

 and
and
Abstract
:1. Introduction
2. Results
2.1. Identification of Potential Therapeutic Targets
2.2. Topology Analysis of the PPI Network and Acquisition of Core Targets
2.3. Enrichment Analysis of the Core Targets of Galangin in PD
2.4. Molecular Docking Assessment
2.5. Galangin Decreases Cell Death in 6-OHDA-Treated HT22 Cells
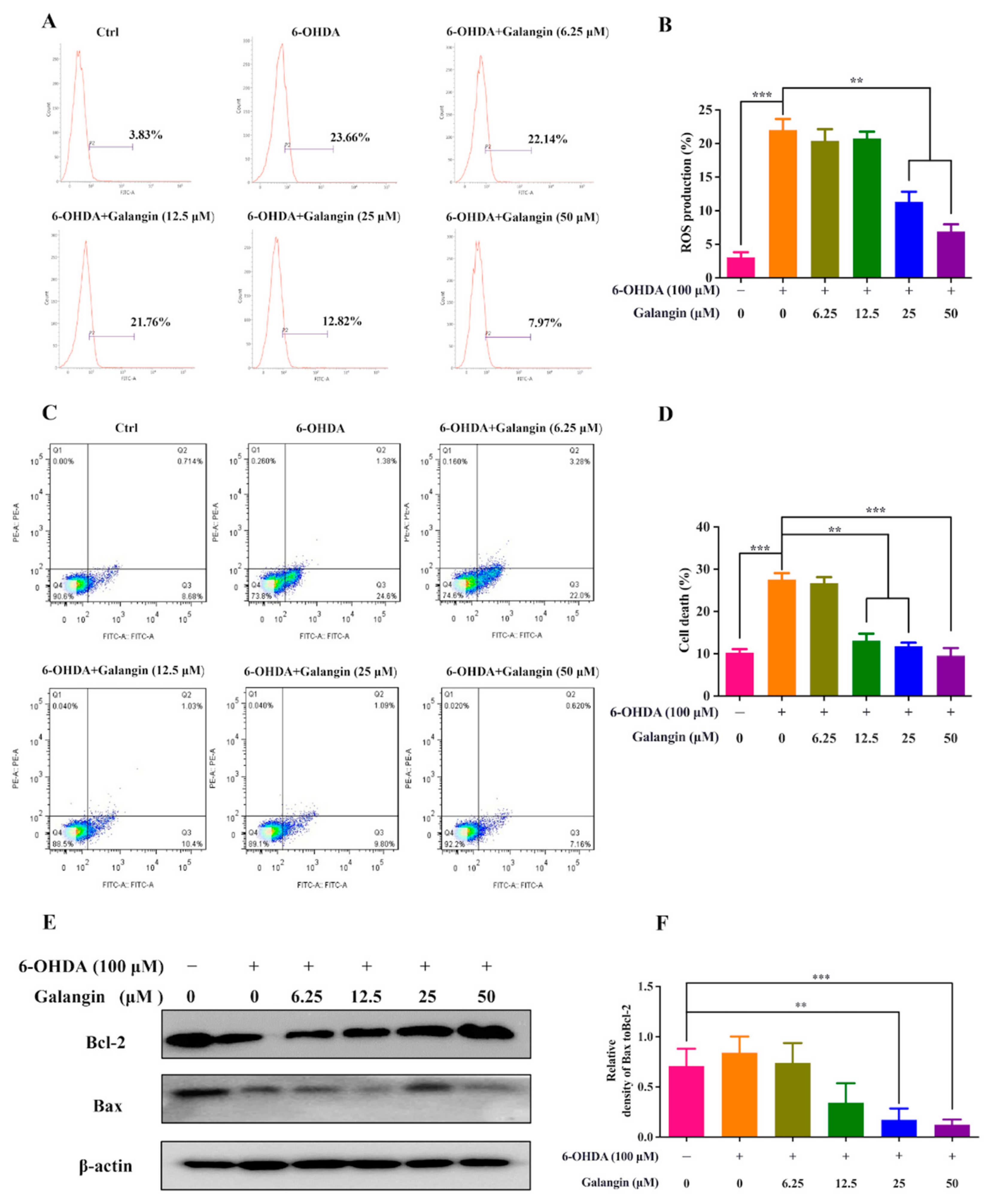
2.6. Galangin Inhibits ROS Production and Cell Apoptosis in 6-OHDA-Treated HT22 Cells
2.7. Galangin Activates the Nrf2/Keap1/HO-1 Signaling Pathway
2.8. Galangin Exhibits Neuroprotective Effects in 6-OHDA-Treated BZ555 Worms
3. Discussion
4. Materials and Methods
4.1. Acquisition of Potential Targets of Galangin against PD
4.2. Protein–Protein Interaction (PPI) Network Construction and Screening of Core Targets
4.3. GO and KEGG Pathway Enrichment Analyses of Core Targets
4.4. Molecular Docking Simulation
4.5. Materials and Reagents
4.6. Cell Culture
4.7. MTT Assay
4.8. Hoechst 33342/PI Staining
4.9. Cell Apoptosis Rate Analysis
4.10. Cellular ROS Measurement
4.11. Western Blot Analysis
4.12. C. elegans Strains, Culture, and Synchronization
4.13. Quantitative Analysis of Dopamine (DA) Neurodegeneration
4.14. Food-Sensing Behavioral Test
4.15. Measurement of ROS Levels in C. elegans
4.16. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rizek, P.; Kumar, N.; Jog, M.S. An update on the diagnosis and treatment of Parkinson disease. Can Med. Assoc. J. 2016, 188, 1157–1165. [Google Scholar] [CrossRef] [PubMed]
- Postuma, R.B.; Berg, D.; Stern, M.; Poewe, W.; Olanow, C.W.; Oertel, W.; Obeso, J.; Marek, K.; Litvan, I.; Lang, A.E.; et al. MDS clinical diagnostic criteria for Parkinson’s disease. Mov. Disord. 2015, 30, 1591–1601. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Violante, M.; Ospina-García, N.; Dávila-Avila, N.M.; Cruz-Fino, D.; Cruz-Landero, A.; Cervantes-Arriaga, A. Motor and non-motor wearing-off and its impact in the quality of life of patients with Parkinson’s disease. Arq. Neuropsiquiatr. 2018, 76, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Blesa, J.; Trigo-Damas, I.; Quiroga-Varela, A.; Jackson-Lewis, V.R. Oxidative stress and Parkinson’s disease. Front. Neuroanat. 2015, 9, 91. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Raymick, J.; Imam, S. Neuroprotective and Therapeutic Strategies against Parkinson’s Disease: Recent Perspectives. Int. J. Mol. Sci. 2016, 17, 904. [Google Scholar] [CrossRef] [PubMed]
- Archibald, N.K.; Clarke, M.P.; Mosimann, U.P.; Burn, D.J. The retina in Parkinson’s disease. Brain 2019, 132, 1128–1145. [Google Scholar] [CrossRef]
- Bodis-Wollner, I. Retinopathy in Parkinson Disease. J. Neural. Transm. 2009, 116, 1493–1501. [Google Scholar] [CrossRef]
- Wu, A.G.; Yong, Y.Y.; Pan, Y.R.; Zhang, L.; Wu, J.M.; Zhang, Y.; Tang, Y.; Wei, J.; Yu, L.; Law, B.Y.; et al. Targeting Nrf2-Mediated Oxidative Stress Response in Traumatic Brain Injury: Therapeutic Perspectives of Phytochemicals. Oxid Med. Cell Longev. 2022, 2022, 1015791. [Google Scholar] [CrossRef]
- Yan, L.; Guo, M.S.; Zhang, Y.; Yu, L.; Wu, J.M.; Tang, Y.; Ai, W.; Zhu, F.D.; Law, B.Y.; Chen, Q.; et al. Dietary Plant Polyphenols as the Potential Drugs in Neurodegenerative Diseases: Current Evidence, Advances, and Opportunities. Oxid Med. Cell Longev. 2022, 2022, 5288698. [Google Scholar] [CrossRef]
- Naylor, J.; Minard, A.; Gaunt, H.J.; Amer, M.S.; Wilson, L.A.; Migliore, M.; Cheung, S.Y.; Rubaiy, H.N.; Blythe, N.M.; Musialowski, K.E.; et al. Natural and synthetic flavonoid modulation of TRPC5 channels. Br. J. Pharmacol. 2016, 173, 562–574. [Google Scholar] [CrossRef]
- Zhang, H.; Li, N.; Wu, J.; Su, L.; Chen, X.; Lin, B.; Luo, H. Galangin inhibits proliferation of HepG2 cells by activating AMPK via increasing the AMP/TAN ratio in a LKB1-independent manner. Eur. J. Pharmacol. 2013, 718, 235–244. [Google Scholar] [CrossRef]
- Patel, D.K.; Patel, K.; Tahilyani, V. Barbaloin: A concise report of its pharmacological and analytical aspects. Asian Pac. J. Trop. Biomed. 2012, 2, 835–838. [Google Scholar] [CrossRef]
- Hopkins, A.L. Network pharmacology. Nat. Biotechnol. 2007, 25, 1110–1111. [Google Scholar] [CrossRef]
- Hsin, K.Y.; Ghosh, S.; Kitano, H. Combining machine learning systems and multiple docking simulation packages to improve docking prediction reliability for network pharmacology. PLoS ONE 2013, 8, e83922. [Google Scholar] [CrossRef]
- Gu, L.; Lu, J.; Li, Q.; Wu, N.; Zhang, L.; Li, H.; Xing, W.; Zhang, X. A network-based analysis of key pharmacological pathways of Andrographis paniculata acting on Alzheimer’s disease and experimental validation. J. Ethnopharmacol. 2020, 251, 112488. [Google Scholar] [CrossRef]
- Simola, N.; Morelli, M.; Carta, A.R. The 6-hydroxydopamine model of Parkinson’s disease. Neurotox. Res. 2007, 11, 151–167. [Google Scholar] [CrossRef]
- Ouyang, M.; Shen, X. Critical role of ASK1 in the 6-hydroxydopamine-induced apoptosis in human neuroblastoma SH-SY5Y cells. J. Neurochem. 2006, 97, 234–244. [Google Scholar] [CrossRef]
- Jeong, W.S.; Jun, M.; Kong, A.N. Nrf2: A potential molecular target for cancer chemoprevention by natural compounds. Antioxid. Redox Signal. 2006, 8, 99–106. [Google Scholar] [CrossRef]
- Taguchi, K.; Motohashi, H.; Yamamoto, M. Molecular mechanisms of the Keap1–Nrf2 pathway in stress response and cancer evolution. Genes Cells 2011, 16, 123–140. [Google Scholar] [CrossRef]
- Sun, Z.; Zhang, S.; Chan, J.Y.; Zhang, D.D. Keap1 controls postinduction repression of the Nrf2-mediated antioxidant response by escorting nuclear export of Nrf2. Mol. Cell Biol. 2007, 27, 6334–6349. [Google Scholar] [CrossRef]
- Jadiya, P.; Khan, A.; Sammi, S.R.; Kaur, S.; Mir, S.S.; Nazir, A. Anti-Parkinsonian effects of Bacopa monnieri: Insights from transgenic and pharmacological Caenorhabditis elegans models of Parkinson’s disease. Biochem. Biophys. Res. Commun. 2011, 413, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Fu, R.H.; Wang, Y.C.; Chen, C.S.; Tsai, R.T.; Liu, S.P.; Chang, W.L.; Lin, H.L.; Lu, C.H.; Wei, J.R.; Wang, Z.W.; et al. Acetylcorynoline attenuates dopaminergic neuron degeneration and α-synuclein aggregation in animal models of Parkinson’s disease. Neuropharmacology 2014, 82, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.G.; Qiu, W.Q.; Yu, L.; Pan, R.; Teng, J.F.; Sang, Z.P.; Law, B.Y.; Zhao, Y.; Zhang, L.; Yan, L.; et al. Targeting microglial autophagic degradation of the NLRP3 inflammasome for identification of thonningianin A in Alzheimer’s disease. Inflamm. Regen. 2022, 42, 25. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.S.; Zhu, F.D.; Qiu, W.Q.; Qiao, G.; Law, B.Y.; Yu, L.; Wu, J.M.; Tang, Y.; Yu, C.L.; Qin, D.L.; et al. High-throughput screening for amyloid-β binding natural small-molecules based on the combinational use of biolayer interferometry and UHPLC-DAD-Q/TOF-MS/MS. Acta Pharm. Sin. B 2022, 12, 1723–1739. [Google Scholar] [CrossRef]
- Qiu, W.Q.; Ai, W.; Zhu, F.D.; Zhang, Y.; Guo, M.S.; Law, B.Y.; Wu, J.M.; Wong, V.K.; Tang, Y.; Yu, L.; et al. Polygala saponins inhibit NLRP3 inflammasome-mediated neuroinflammation via SHP-2-Mediated mitophagy. Free Radic. Biol. Med. 2022, 179, 76–94. [Google Scholar] [CrossRef]
- Sun, Y.S.; Jiang, X.Q.; Pan, R.; Zhou, X.G.; Qin, D.L.; Xiong, R.; Wang, Y.L.; Qiu, W.Q.; Wu, A.G.; Wu, J.M. Escins Isolated from Aesculus chinensis Bge. Promote the Autophagic Degradation of Mutant Huntingtin and Inhibit its Induced Apoptosis in HT22 cells. Front. Pharmacol. 2020, 11, 116. [Google Scholar] [CrossRef]
- Al-Numair, K.S.; Chandramohan, G.; Veeramani, C.; Alsaif, M.A. Ameliorative effect of kaempferol, a flavonoid, on oxidative stress in streptozotocin-induced diabetic rats. Redox Rep. 2015, 20, 198–209. [Google Scholar] [CrossRef]
- Zou, W.W.; Xu, S.P. Galangin inhibits the cell progression and induces cell apoptosis through activating PTEN and Caspase-3 pathways in retinoblastoma. Biomed. Pharmacother. 2018, 97, 851–863. [Google Scholar] [CrossRef]
- Aladaileh, S.H.; Abukhalil, M.H.; Saghir, S.A.M.; Hanieh, H.; Alfwuaires, M.A.; Almaiman, A.A.; Bin-Jumah, M.; Mahmoud, A.M. Galangin Activates Nrf2 Signaling and Attenuates Oxidative Damage, Inflammation, and Apoptosis in a Rat Model of Cyclophosphamide-Induced Hepatotoxicity. Biomolecules 2019, 9, 346. [Google Scholar] [CrossRef]
- Abukhalil, M.H.; Althunibat, O.Y.; Aladaileh, S.H.; Al-Amarat, W.; Obeidat, H.M.; Al-Khawalde, A.A.A.; Hussein, O.E.; Alfwuaires, M.A.; Algefare, A.I.; Alanazi, K.M.; et al. Galangin attenuates diabetic cardiomyopathy through modulating oxidative stress, inflammation and apoptosis in rats. Biomed. Pharmacother. 2021, 138, 111410. [Google Scholar] [CrossRef]
- Abd El-Aal, S.A.; AbdElrahman, M.; Reda, A.M.; Afify, H.; Ragab, G.M.; El-Gazar, A.A.; Ibrahim, S.S.A. Galangin mitigates DOX-induced cognitive impairment in rats: Implication of NOX-1/Nrf-2/HMGB1/TLR4 and TNF-α/MAPKs/RIPK/MLKL/BDNF. Neurotoxicology 2022, 92, 77–90. [Google Scholar] [CrossRef]
- Li, S.J.; Wu, C.H.; Zhu, L.; Gao, J.; Fang, J.; Li, D.F.; Fu, M.H.; Liang, R.X.; Wang, L.; Cheng, M. By improving regional cortical blood flow, attenuating mitochondrial dysfunction and sequential apoptosis galangin acts as a potential neuroprotective agent after acute ischemic stroke. Molecules 2012, 17, 13403–13423. [Google Scholar] [CrossRef]
- Wu, A.G.; Zhou, X.G.; Qiao, G.; Yu, L.; Tang, Y.; Yan, L.; Qiu, W.Q.; Pan, R.; Yu, C.L.; Law, B.Y.; et al. Targeting microglial autophagic degradation in NLRP3 inflammasome-mediated neurodegenerative diseases. Ageing Res. Rev. 2021, 65, 101202. [Google Scholar] [CrossRef]
- Chen, G.; Liu, J.; Jiang, L.; Ran, X.; He, D.; Li, Y.; Huang, B.; Wang, W.; Fu, S. Galangin Reduces the Loss of Dopaminergic Neurons in an LPS-Evoked Model of Parkinson’s Disease in Rats. Int. J. Mol. Sci. 2017, 19, 12. [Google Scholar] [CrossRef]
- Nogales, C.; Mamdouh, Z.M.; List, M.; Kiel, C.; Casas, A.I.; Schmidt, H. Network pharmacology: Curing causal mechanisms instead of treating symptoms. Trends Pharmacol. Sci. 2022, 43, 136–150. [Google Scholar] [CrossRef]
- Pinzi, L.; Rastelli, G. Molecular Docking: Shifting Paradigms in Drug Discovery. Int. J. Mol. Sci. 2019, 20, 4331. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Zou, L.; Zhang, X.; Branco, V.; Wang, J.; Carvalho, C.; Holmgren, A.; Lu, J. Redox Signaling Mediated by Thioredoxin and Glutathione Systems in the Central Nervous System. Antioxid Redox Signal. 2017, 27, 989–1010. [Google Scholar] [CrossRef] [PubMed]
- Hybertson, B.M.; Gao, B.; Bose, S.K.; McCord, J.M. Oxidative stress in health and disease: The therapeutic potential of Nrf2 activation. Mol. Asp. Med. 2011, 32, 234–246. [Google Scholar] [CrossRef]
- Parga, J.A.; Rodriguez-Perez, A.I.; Garcia-Garrote, M.; Rodriguez-Pallares, J.; Labandeira-Garcia, J.L. NRF2 Activation and Downstream Effects: Focus on Parkinson’s Disease and Brain Angiotensin. Antioxidants 2021, 10, 1649. [Google Scholar] [CrossRef]
- Martins, I.K.; de Carvalho, N.R.; Macedo, G.E.; Rodrigues, N.R.; Ziech, C.C.; Vinadé, L.; Filho, V.M.B.; Menezes, I.A.; Franco, J.; Posser, T. Anacardium microcarpum Promotes Neuroprotection Dependently of AKT and ERK Phosphorylation but Does Not Prevent Mitochondrial Damage by 6-OHDA. Oxid. Med. Cell Longev. 2018, 2018, 2131895. [Google Scholar] [CrossRef]
- Bernstein, A.I.; Garrison, S.P.; Zambetti, G.P.; O’Malley, K.L. 6-OHDA generated ROS induces DNA damage and p53- and PUMA-dependent cell death. Mol. Neurodegener. 2011, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Alexander, A.G.; Marfil, V.; Li, C. Use of Caenorhabditis elegans as a model to study Alzheimer’s disease and other neurodegenerative diseases. Front. Genet. 2014, 5, 279. [Google Scholar] [CrossRef] [PubMed]
- Bian, Y.; Chen, Y.; Wang, X.; Cui, G.; Ung, C.O.L.; Lu, J.H.; Cong, W.; Tang, B.; Lee, S.M. Oxyphylla A ameliorates cognitive deficits and alleviates neuropathology via the Akt-GSK3β and Nrf2-Keap1-HO-1 pathways in vitro and in vivo murine models of Alzheimer’s disease. J. Adv. Res. 2021, 34, 1–12. [Google Scholar] [CrossRef]
- Ragunathan, A.; Malathi, K.; Ramaiah, S.; Anbarasu, A. FtsA as a cidal target for Staphylococcus aureus: Molecular docking and dynamics studies. J. Cell Biochem. 2018, 120, 7751–7758. [Google Scholar] [CrossRef]
- Davis, J.B.; Maher, P. Protein kinase C activation inhibits glutamate-induced cytotoxicity in a neuronal cell line. Brain Res. 1994, 652, 169–173. [Google Scholar] [CrossRef]
- Chang, X.; Wang, X.; Li, J.; Shang, M.; Niu, S.; Zhang, W.; Li, Y.; Sun, Z.; Gan, J.; Li, W.; et al. Silver nanoparticles induced cytotoxicity in HT22 cells through autophagy and apoptosis via PI3K/AKT/mTOR signaling pathway. Ecotoxicol. Environ. Saf. 2021, 208, 111696. [Google Scholar] [CrossRef]
- He, C.L.; Tang, Y.; Wu, J.M.; Long, T.; Yu, L.; Teng, J.F.; Qiu, W.Q.; Pan, R.; Yu, C.L.; Qin, D.L.; et al. Chlorogenic acid delays the progression of Parkinson’s disease via autophagy induction in Caenorhabditis elegans. Nutr. Neurosci. 2021, 1–14, ahead of print. [Google Scholar] [CrossRef]
- Wu, A.G.; Pan, R.; Law, B.Y.; Qiu, W.Q.; Wu, J.M.; He, C.L.; Wong, V.K.; Yu, C.L.; Zhou, X.G.; Qin, D.L.; et al. Targeting autophagy as a therapeutic strategy for identification of liganans from Peristrophe japonica in Parkinson’s disease. Signal. Transduct. Target Ther. 2021, 6, 67. [Google Scholar] [CrossRef]
- Kamireddy, K.; Chinnu, S.; Priyanka, P.S.; Rajini, P.S.; Giridhar, P. Neuroprotective effect of Decalepis hamiltonii aqueous root extract and purified 2-hydroxy-4-methoxy benzaldehyde on 6-OHDA induced neurotoxicity in Caenorhabditis elegans. Biomed. Pharmacother. 2018, 105, 997–1005. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | URL | Access Date |
|---|---|---|
| Pubchem | https://pubchem.ncbi.nlm.nih.gov/ | 21 January 2022 |
| Swiss Target Prediction | http://swisstargetprediction.ch/ | 21 January 2022 |
| DisGeNET | http://www.disgenet.org/ | 21 January 2022 |
| OMIM | https://omim.org | 21 January 2022 |
| GeneCards | https://www.genecards.org/ | 21 January 2022 |
| STRING 11.0 | https://string-db.org/ | 24 January 2022 |
| DAVID 6.8 | https://david.ncifcrf.gov/ | 24 January 2022 |
| OmicShare Tools | https://www.omicshare.com/tools/ | 24 January 2022 |
| UniProt | https://www.uniprot.org | 25 January 2022 |
| Cytoscape 3.7.1 | https://cytoscape.org/ | 12 August 2022 |
| Sybyl-X 2.0 | https://sybyl.com | 25 January 2022 |
| PharmMapper | http://www.lilab-ecust.cn/pharmmapper/ | 21 January 2022 |
| Reagents and Antibodiese | Source | Identifier |
|---|---|---|
| 6-OHDA | Sigma–Aldrich, St. Louis, MO, USA | H4381 |
| MTT | Sigma–Aldrich, St. Louis, MO, USA | MKCD4805 |
| 5-Fluoro-2′-deoxyuridine (FUDR) | Sigma–Aldrich, St. Louis, USA | F0503 |
| Galangin | Chengdu Desite Biological Technology Co., Ltd., Chengdu, China | DG0020-0020 |
| Levodopa | Topscience Company, Ltd., Shanghai, China | T0848 |
| Dulbecco’s Modified Eagle Medium (DMEM) | Gibico, Carlsbad, CA, USA | 10741574 |
| Fetal bovine serum (FBS) | Gibico, Carlsbad, CA, USA | 10099141 |
| Trypsin-EDTA solution | Gibico, Carlsbad, CA, USA | 11590626 |
| Penicillin–streptomycin solution (100×) | Gibico, Carlsbad, CA, USA | 15140122 |
| H2DCFDA fluorescence probe | Invitrogen, Carlsbad, CA, USA | D399 |
| Annexin V-FITC/PI Apoptosis detection Kit | 4A Biotech Co., Ltd., Beijing, China | 20190221 |
| Bax | CST, Beverly, MA, USA | 2772 |
| Bcl-2 | CST, Beverly, MA, USA | 15071S |
| Keap1 | CST, Beverly, MA, USA | 4678S |
| Nrf2 | CST, Beverly, MA, USA | 12721S |
| HO-1 | CST, Beverly, MA, USA | 70081S |
| β-actin | Santa Cruz Biotechnology, Dallas, TX, USA | sc-47778 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Q.-X.; Zhou, L.; Long, T.; Qin, D.-L.; Wang, Y.-L.; Ye, Y.; Zhou, X.-G.; Wu, J.-M.; Wu, A.-G. Galangin Exhibits Neuroprotective Effects in 6-OHDA-Induced Models of Parkinson’s Disease via the Nrf2/Keap1 Pathway. Pharmaceuticals 2022, 15, 1014. https://doi.org/10.3390/ph15081014
Chen Q-X, Zhou L, Long T, Qin D-L, Wang Y-L, Ye Y, Zhou X-G, Wu J-M, Wu A-G. Galangin Exhibits Neuroprotective Effects in 6-OHDA-Induced Models of Parkinson’s Disease via the Nrf2/Keap1 Pathway. Pharmaceuticals. 2022; 15(8):1014. https://doi.org/10.3390/ph15081014
Chicago/Turabian StyleChen, Qiu-Xu, Ling Zhou, Tao Long, Da-Lian Qin, Yi-Ling Wang, Yun Ye, Xiao-Gang Zhou, Jian-Ming Wu, and An-Guo Wu. 2022. "Galangin Exhibits Neuroprotective Effects in 6-OHDA-Induced Models of Parkinson’s Disease via the Nrf2/Keap1 Pathway" Pharmaceuticals 15, no. 8: 1014. https://doi.org/10.3390/ph15081014