
Purification, Characterization and Evaluation of the Antitumoral Activity of a Phospholipase A2 from the Snake Bothrops moojeni

 , ,
, ,  , ,
, ,  and
and
Abstract
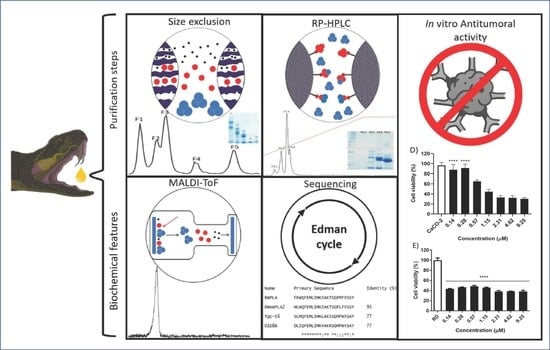
:
1. Introduction
2. Results
2.1. Purification and Biochemical Characterization
2.2. Enzymatic Activity
2.3. Hemolytic Activity
2.4. Anticancer Activity
3. Discussion
4. Materials and Methods
4.1. Bothrops Moojeni Venom, Cell Lines and Ethics Committee Guidelines
4.2. B. moojeni Venom Purification
4.3. SDS-PAGE Electrophoresis
4.4. Quantification of Proteins
4.5. Mass Spectrometry
4.6. Amino-Terminal Sequencing
4.7. Phospholipase Activity
4.8. Hemolytic Assay
4.9. Anticancer Activity
4.9.1. Cell Culture
4.9.2. Cytotoxicity by MTT Assay
4.9.3. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Redig, A.J.; Mcallister, S.S. Breast Cancer as a Systemic Disease: A View of Metastasis. J. Intern. Med. 2013, 274, 113–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mokhtari, R.B.; Homayouni, T.S.; Baluch, N.; Morgatskaya, E.; Kumar, S.; Das, B.; Yeger, H. Combination Therapy in Combating Cancer. Oncotarget 2017, 8, 38022–38043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waheed, H.; Moin, S.F.; Choudhary, M.I. Snake Venom: From Deadly Toxins to Life-Saving Therapeutics. Curr. Med. Chem. 2017, 24, 1874–1891. [Google Scholar] [CrossRef]
- Kang, T.S.; Georgieva, D.; Genov, N.; Murakami, M.T.; Sinha, M.; Kumar, R.P.; Kaur, P.; Kumar, S.; Dey, S.; Sharma, S.; et al. Enzymatic Toxins from Snake Venom: Structural Characterization and Mechanism of Catalysis. FEBS J. 2011, 278, 4544–4576. [Google Scholar] [CrossRef] [PubMed]
- Munawar, A.; Ali, S.A.; Akrem, A.; Betzel, C. Snake Venom Peptides: Tools of Biodiscovery. Toxins 2018, 10, 474. [Google Scholar] [CrossRef] [Green Version]
- Xiao, H.; Pan, H.; Liao, K.; Yang, M.; Huang, C. Snake Venom PLA2, a Promising Target for Broad-Spectrum Antivenom Drug Development. Biomed Res. Int. 2017, 2017, 6592820. [Google Scholar] [CrossRef] [Green Version]
- Quach, N.D.; Arnold, R.D.; Cummings, B.S. Secretory Phospholipase A2 Enzymes as Pharmacological Targets for Treatment of Disease. Biochem. Pharmacol. 2014, 90, 338–348. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, F.V.P.V.; Lopes, D.S.; Cirilo Gimenes, S.N.; Achê, D.C.; Vecchi, L.; Alves, P.T.; Guimarães, D.D.O.; Rodrigues, R.S.; Goulart, L.R.; Rodrigues, V.D.M.; et al. Human Breast Cancer Cell Death Induced by BnSP-6, a Lys-49 PLA2 Homologue from Bothrops Pauloensis Venom. Int. J. Biol. Macromol. 2016, 82, 671–677. [Google Scholar] [CrossRef]
- Cedro, R.C.A.; Menaldo, D.L.; Costa, T.R.; Zoccal, K.F.; Sartim, M.A.; Santos-Filho, N.A.; Faccioli, L.H.; Sampaio, S.V. Cytotoxic and Inflammatory Potential of a Phospholipase A2 from Bothrops Jararaca Snake Venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Daniele, J.J.; Bianco, I.D.; Delgado, C.; Carrillo, D.B.; Fidelio, G.D. A New Phospholipase A2 Isoform Isolated from Bothrops Neuwiedii (Yarara Chica) Venom with Novel Kinetic and Chromatographic Properties. Toxicon 1997, 35, 1205–1215. [Google Scholar] [CrossRef]
- De Vasconcelos Azevedo, F.V.P.; Zóia, M.A.P.; Lopes, D.S.; Gimenes, S.N.; Vecchi, L.; Alves, P.T.; Rodrigues, R.S.; Silva, A.C.A.; Yoneyama, K.A.G.; Goulart, L.R.; et al. Antitumor and Antimetastatic Effects of PLA2-BthTX-II from Bothrops Jararacussu Venom on Human Breast Cancer Cells. Int. J. Biol. Macromol. 2019, 135, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Stábeli, R.G.; Amui, S.F.; Sant’Ana, C.D.; Pires, M.G.; Nomizo, A.; Monteiro, M.C.; Romão, P.R.T.; Guerra-Sá, R.; Vieira, C.A.; Giglio, J.R.; et al. Bothrops Moojeni Myotoxin-II, a Lys49-Phospholipase A2 Homologue: An Example of Function Versatility of Snake Venom Proteins. Comp. Biochem. Physiol.-C Toxicol. Pharmacol. 2006, 142, 371–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderon, L.A.; Sobrinho, J.C.; Zaqueo, K.D.; de Moura, A.A.; Grabner, A.N.; Mazzi, M.V.; Marcussi, S.; Nomizo, A.; Fernandes, C.F.C.; Zuliani, J.P.; et al. Antitumoral Activity of Snake Venom Proteins: New Trends in Cancer Therapy. Biomed Res. Int. 2014, 2014, 203639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damico, D.C.S.; Lilla, S.; de Nucci, G.; Ponce-Soto, L.A.; Winck, F.V.; Novello, J.C.; Marangoni, S. Biochemical and Enzymatic Characterization of Two Basic Asp49 Phospholipase A2 Isoforms from Lachesis Muta Muta (Surucucu) Venom. Biochim. Biophys. Acta 2005, 1726, 75–86. [Google Scholar] [CrossRef]
- Denegri, M.E.G.; Acosta, O.C.; Huancahuire-Vega, S.; Martins-de-Souza, D.; Marangoni, S.; Maruñak, S.L.; Teibler, G.P.; Leiva, L.C.; Ponce-Soto, L.A. Isolation and Functional Characterization of a New Acidic PLA2 Ba SpII RP4 of the Bothrops Alternatus Snake Venom from Argentina. Toxicon 2010, 56, 64–74. [Google Scholar] [CrossRef]
- Menaldo, D.L.; Jacob-Ferreira, A.L.; Bernardes, C.P.; Cintra, A.C.O.; Sampaio, S.V. Purification Procedure for the Isolation of a P-I Metalloprotease and an Acidic Phospholipase A2 from Bothrops Atrox Snake Venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 28. [Google Scholar] [CrossRef] [Green Version]
- Pereañez, J.A.; Núñez, V.; Huancahuire-Vega, S.; Marangoni, S.; Ponce-Soto, L.A. Biochemical and Biological Characterization of a PLA2 from Crotoxin Complex of Crotalus Durissus Cumanensis. Toxicon 2009, 53, 534–542. [Google Scholar] [CrossRef]
- Stábeli, R.G.; Simões-Silva, R.; Kayano, A.M.; Gimenez, G.S.; Moura, A.A.; Caldeira, C.A.S.; Coutinho-Neto, A.; Zaqueo, K.D.; Zuliani, J.P.; Calderon, L.A. Purification of Phospholipases A2 from American Snake Venoms. In Chromatography—The Most Versatile Method of Chemical Analysis; Calderón, L.A., Ed.; Intechopen: London, UK, 2012; pp. 1–34. [Google Scholar]
- Tsai, I.H.; Chen, Y.H.; Wang, Y.M.; Tu, M.C.; Tu, A.T. Purification, Sequencing, and Phylogenetic Analyses of Novel Lys-49 Phospholipases A(2) from the Venoms of Rattlesnakes and Other Pit Vipers. Arch. Biochem. Biophys. 2001, 394, 236–244. [Google Scholar] [CrossRef]
- Calgarotto, A.K.; Damico, D.C.S.; Ponce-Soto, L.A.; Baldasso, P.A.; Da Silva, S.L.; Souza, G.H.M.F.; Eberlin, M.N.; Marangoni, S. Biological and Biochemical Characterization of New Basic Phospholipase A2 BmTX-I Isolated from Bothrops Moojeni Snake Venom. Toxicon 2008, 51, 1509–1519. [Google Scholar] [CrossRef]
- Galbiatti, C.; Rocha, T.; Randazzo-Moura, P.; Ponce-Soto, L.A.; Marangoni, S.; Cruz-Höfling, M.A.; Rodrigues-Simioni, L. Pharmacological and Partial Biochemical Characterization of Bmaj-9 Isolated from Bothrops Marajoensis Snake Venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2012, 18, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Ponce-Soto, L.A.; Baldasso, P.A.; Romero-Vargas, F.F.; Winck, F.V.; Novello, J.C.; Marangoni, S. Biochemical, Pharmacological and Structural Characterization of Two PLA2 Isoforms Cdr-12 and Cdr-13 from Crotalus Durissus Ruruima Snake Venom. Protein J. 2007, 26, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Ponce-Soto, L.A.; Lomonte, B.; Gutiérrez, J.M.; Rodrigues-Simioni, L.; Novello, J.C.; Marangoni, S. Structural and Functional Properties of BaTX, a New Lys49 Phospholipase A2 Homologue Isolated from the Venom of the Snake Bothrops Alternatus. Biochim. Biophys. Acta-Gen. Subj. 2007, 1770, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Costa, T.R.; Menaldo, D.L.; Oliveira, C.Z.; Santos-Filho, N.A.; Teixeira, S.S.; Nomizo, A.; Fuly, A.L.; Monteiro, M.C.; de Souza, B.M.; Palma, M.S. Myotoxic Phospholipases A2 Isolated from Bothrops Brazili Snake Venom and Synthetic Peptides Derived from Their C-Terminal Region: Cytotoxic Effect on Microorganism and Tumor Cells. Peptides 2008, 29, 1645–1656. [Google Scholar] [CrossRef]
- Díaz Oreiro, C.; Lomonte, B.; Zamudio, F.; Gutiérrez, J.M. Purification and Characterization of Myotoxin IV, a Phospholipase A2 Variant, from Bothrops Asper Snake Venom. Nat. Toxins 1995, 3, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Nunes, E.; Frihling, B.; Barros, E.; de Oliveira, C.; Verbisck, N.; Flores, T.; de Freitas Júnior, A.; Franco, O.; de Macedo, M.; Migliolo, L.; et al. Antibiofilm Activity of Acidic Phospholipase Isoform Isolated from Bothrops Erythromelas Snake Venom. Toxins 2013, 12, 35–43. [Google Scholar] [CrossRef]
- Soares, A.M.; Andrião-Escarso, S.H.; Bortoleto, R.K.; Rodrigues-Simioni, L.; Arni, R.K.; Ward, R.J.; Gutiérrez, J.M.; Giglio, J.R. Dissociation of Enzymatic and Pharmacological Properties of Piratoxins-I and-III, Two Myotoxic Phospholipases A2 from Bothrops Pirajai Snake Venom. Arch. Biochem. Biophys. 2001, 387, 188–196. [Google Scholar] [CrossRef]
- Borges, I.P.; Castanheira, L.E.; Barbosa, B.F.; de Souza, D.L.N.; da Silva, R.J.; Mineo, J.R.; Tudini, K.A.Y.; Rodrigues, R.S.; Ferro, E.A.V.; de Melo Rodrigues, V. Anti-Parasitic Effect on Toxoplasma Gondii Induced by BnSP-7, a Lys49-Phospholipase A2 Homologue from Bothrops Pauloensis Venom. Toxicon 2016, 119, 84–91. [Google Scholar] [CrossRef]
- Moreira, V.; Gutiérrez, J.M.; Soares, A.M.; Zamunér, S.R.; Purgatto, E.; Teixeira, C.D.F.P. Secretory Phospholipases A2 Isolated from Bothrops Asper and from Crotalus Durissus Terrificus Snake Venoms Induce Distinct Mechanisms for Biosynthesis of Prostaglandins E2 and D2 and Expression of Cyclooxygenases. Toxicon 2008, 52, 428–439. [Google Scholar] [CrossRef]
- Trento, E.P.; Garcia, O.S.; Rucavado, A.; França, S.C.; Batalini, C.; Arantes, E.C.; Giglio, J.R.; Soares, A.M. Inhibitory Properties of the Anti-Bothropic Complex from Didelphis Albiventris Serum on Toxic and Pharmacological Actions of Metalloproteases and Myotoxins from Bothrops Asper Venom. Biochem. Pharmacol. 2001, 62, 1521–1529. [Google Scholar] [CrossRef]
- Veronese, E.L.G.; Esmeraldino, L.E.; Trombone, A.P.F.; Santana, A.E.; Bechara, G.H.; Kettelhut, I.; Cintra, A.C.O.; Giglio, J.R.; Sampaio, S.V. Inhibition of the Myotoxic Activity of Bothrops Jararacussu Venom and Its Two Major Myotoxins, BthTX-I and BthTX-II, by the Aqueous Extract of Tabernaemontana Catharinensis A. DC.(Apocynaceae). Phytomedicine 2005, 12, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Ximenes, R.M.; Rabello, M.M.; Araújo, R.M.; Silveira, E.R.; Fagundes, F.H.R.; Diz-Filho, E.; Buzzo, S.C.; Soares, V.C.G.; Toyama, D.D.O.; Gaeta, H.H. Inhibition of Neurotoxic Secretory Phospholipases A2 Enzymatic, Edematogenic, and Myotoxic Activities by Harpalycin 2, an Isoflavone Isolated from Harpalyce Brasiliana Benth. Evid.-Based Complement. Altern. Med. 2012, 2012, 987517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Albuquerque Modesto, J.C.; Spencer, P.J.; Fritzen, M.; Valenca, R.C.; Oliva, M.L.V.; Bezerra da Silva, M.; Chudzinski-Tavassi, A.M.; Camargo Guarnieri, M. BE-I-PLA2, a Novel Acidic Phospholipase A (2) from Bothrops Erythromelas Venom: Isolation, Cloning and Characterization as Potent Anti-Platelet and Inductor of Prostaglandin I-2 Release by Endothelial Cells. Biochem. Pharmacol. 2006, 72, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Ketelhut, D.F.J.; de Mello, M.H.; Veronese, E.L.G.; Esmeraldino, L.E.; Murakami, M.T.; Arni, R.M.; Giglio, J.R.; Cintra, A.C.O.; Sampaio, S.V. Isolation, Caharacterization and Biological Activity of Acidic Phospholipase A2 Isoforms from Bothrops Jararacussu Snake Venom. Biochimie 2003, 85, 983–991. [Google Scholar] [CrossRef]
- Silveira, L.B.; Marchi-Salvador, D.P.; Santos-Filho, N.A.; Silva Jr, F.P.; Marcussi, S.; Fuly, A.L.; Nomizo, A.; da Silva, S.L.; Stábeli, R.G.; Arantes, E.C. Isolation and Expression of a Hypotensive and Anti-Platelet Acidic Phospholipase A2 from Bothrops Moojeni Snake Venom. J. Pharm. Biomed. Anal. 2013, 73, 35–43. [Google Scholar] [CrossRef]
- Sudharshan, S.; Dhananjaya, B.L. Antibacterial Potential of a Basic Phospholipase A2 (VRV-PL-VIIIa) from Daboia Russelii Pulchella (Russell’s Viper) Venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 17. [Google Scholar] [CrossRef] [Green Version]
- Vitorino, K.A.; Alfonso, J.J.; Gómez, A.F.; Santos, A.P.A.; Antunes, Y.R.; Caldeira, C.A.D.S.; Gómez, C.V.; Teles, C.B.G.; Soares, A.M.; Calderon, L.A. Antimalarial Activity of Basic Phospholipases A2 Isolated from Paraguayan Bothrops Diporus Venom against Plasmodium Falciparum. Toxicon X 2020, 8, 100056. [Google Scholar] [CrossRef]
- Pizzo, S.V.; Schwartz, M.L.; Hill, R.L.; McKee, P.A. Mechanism of Ancrod Anticoagulation. A Direct Proteolytic Effect on Fibrin. J. Clin. Investig. 1972, 51, 2841–2850. [Google Scholar] [CrossRef]
- Gebrim, L.C.; Marcussi, S.; Menaldo, D.L.; de Menezes, C.S.R.; Nomizo, A.; Hamaguchi, A.; Silveira-Lacerda, E.P.; Homsi-Brandeburgo, M.I.; Sampaio, S.V.; Soares, A.M. Antitumor Effects of Snake Venom Chemically Modified Lys49 Phospholipase A2-like BthTX-I and a Synthetic Peptide Derived from Its C-Terminal Region. Biologicals 2009, 37, 222–229. [Google Scholar] [CrossRef]
- Da Silva, C.P.; Costa, T.R.; Paiva, R.M.A.; Cintra, A.C.O.; Menaldo, D.L.; Antunes, L.M.G.; Sampaio, S.V. Antitumor Potential of the Myotoxin BthTX-I from Bothrops Jararacussu Snake Venom: Evaluation of Cell Cycle Alterations and Death Mechanisms Induced in Tumor Cell Lines. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 44. [Google Scholar]
- Moura, A.A.D.; Kayano, A.M.; Oliveira, G.A.; Setúbal, S.S.; Ribeiro, J.G.; Barros, N.B.; Nicolete, R.; Moura, L.A.; Fuly, A.L.; Nomizo, A.; et al. Purification and Biochemical Characterization of Three Myotoxins from Bothrops Mattogrossensis Snake Venom with Toxicity against Leishmania and Tumor Cells. Biomed Res. Int. 2014, 2014, 195356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khunsap, S.; Pakmanee, N.; Khow, O.; Chanhome, L.; Sitprija, V.; Suntravat, M.; Lucena, S.E.; Perez, J.C.; Sánchez, E.E. Purification of a Phospholipase A2 from Daboia Russelii Siamensis Venom with Anticancer Effects. J. Venom Res. 2011, 2, 42. [Google Scholar] [PubMed]
- Cummings, B.S. Phospholipase A2 as Targets for Anti-Cancer Drugs. Biochem. Pharmacol. 2007, 74, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.A.; Kalatardi, S.; Dohse, J.; Sadaria, M.R.; Meng, X.; Fullerton, D.A.; Weyant, M.J. Group IIa SPLA2 Inhibition Attenuates NF-ΚB Activity and Promotes Apoptosis of Lung Cancer Cells. Anticancer Res. 2012, 32, 3601–3607. [Google Scholar] [PubMed]
- Tokunaga, Y.; Yamazaki, Y.; Morita, T. Identification and Localization of Heparin-Binding Region of Snake Venom VEGF and Its Blocking of VEGF-A165. Pathophysiol. Haemost. Thromb. 2005, 34, 194–196. [Google Scholar]
- Montecucco, C.; Gutiérrez, J.M.; Lomonte, B. Cellular Pathology Induced by Snake Venom Phospholipase A2 Myotoxins and Neurotoxins: Common Aspects of Their Mechanisms of Action. Cell. Mol. Life Sci. 2008, 65, 2897–2912. [Google Scholar] [CrossRef]
- Serino-Silva, C.; Morais-Zani, K.; Hikari Toyama, M.; Toyama, D.D.O.; Gaeta, H.H.; Rodrigues, C.F.B.; Aguiar, W.D.S.; Tashima, A.K.; Grego, K.F.; Tanaka-Azevedo, A.M. Purification and Characterization of the First γ-Phospholipase Inhibitor (ΓPLI) from Bothrops Jararaca Snake Serum. PLoS ONE 2018, 13, e0193105. [Google Scholar] [CrossRef] [Green Version]
- Holzer, M.; Mackessy, S.P. An Aqueous Endpoint Assay of Snake Venom Phospholipase A2. Toxicon 1996, 34, 1149–1155. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | PLA2 | Sequence | Identity (%) |
|---|---|---|---|
| B. moojeni | BmPLA | FKWQFEMLIMKIAKTSGFMFYSSY | - |
| B. moojeni | BMOOPLA2 | NLWQFEMLIMKIAKTSGFLFYSSY | 95 |
| T. gracilis | TGC-E6 | SLMQFEMLIMKLAKSSGMFWYSAY | 77 |
| C. godmani | D1E6b | DLIQFEMLIMKVAKRSGMFWYSAY | 77 |
| ********:** **:::**:* |
| Cell Line | IC50 (µM) | IS * (MRC-5) | IS * (BV-2) |
|---|---|---|---|
| MRC-5 | 1.1 | - | - |
| BV-2 | 3.8 | - | - |
| Caco-2 | 0.6 | 1.85 | 6.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frihling, B.E.F.; Boleti, A.P.d.A.; de Oliveira, C.F.R.; Sanches, S.C.; Cardoso, P.H.d.O.; Verbisck, N.; Macedo, M.L.R.; Rita, P.H.S.; Carvalho, C.M.E.; Migliolo, L. Purification, Characterization and Evaluation of the Antitumoral Activity of a Phospholipase A2 from the Snake Bothrops moojeni. Pharmaceuticals 2022, 15, 724. https://doi.org/10.3390/ph15060724
Frihling BEF, Boleti APdA, de Oliveira CFR, Sanches SC, Cardoso PHdO, Verbisck N, Macedo MLR, Rita PHS, Carvalho CME, Migliolo L. Purification, Characterization and Evaluation of the Antitumoral Activity of a Phospholipase A2 from the Snake Bothrops moojeni. Pharmaceuticals. 2022; 15(6):724. https://doi.org/10.3390/ph15060724
Chicago/Turabian StyleFrihling, Breno Emanuel Farias, Ana Paula de Araújo Boleti, Caio Fernando Ramalho de Oliveira, Simone Camargo Sanches, Pedro Henrique de Oliveira Cardoso, Newton Verbisck, Maria Lígia Rodrigues Macedo, Paula Helena Santa Rita, Cristiano Marcelo Espinola Carvalho, and Ludovico Migliolo. 2022. "Purification, Characterization and Evaluation of the Antitumoral Activity of a Phospholipase A2 from the Snake Bothrops moojeni" Pharmaceuticals 15, no. 6: 724. https://doi.org/10.3390/ph15060724