
Protective Effects of Naringin–Dextrin Nanoformula against Chemically Induced Hepatocellular Carcinoma in Wistar Rats: Roles of Oxidative Stress, Inflammation, Cell Apoptosis, and Proliferation

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Characterization, Entrapment Efficiency (EE), and Release Profile
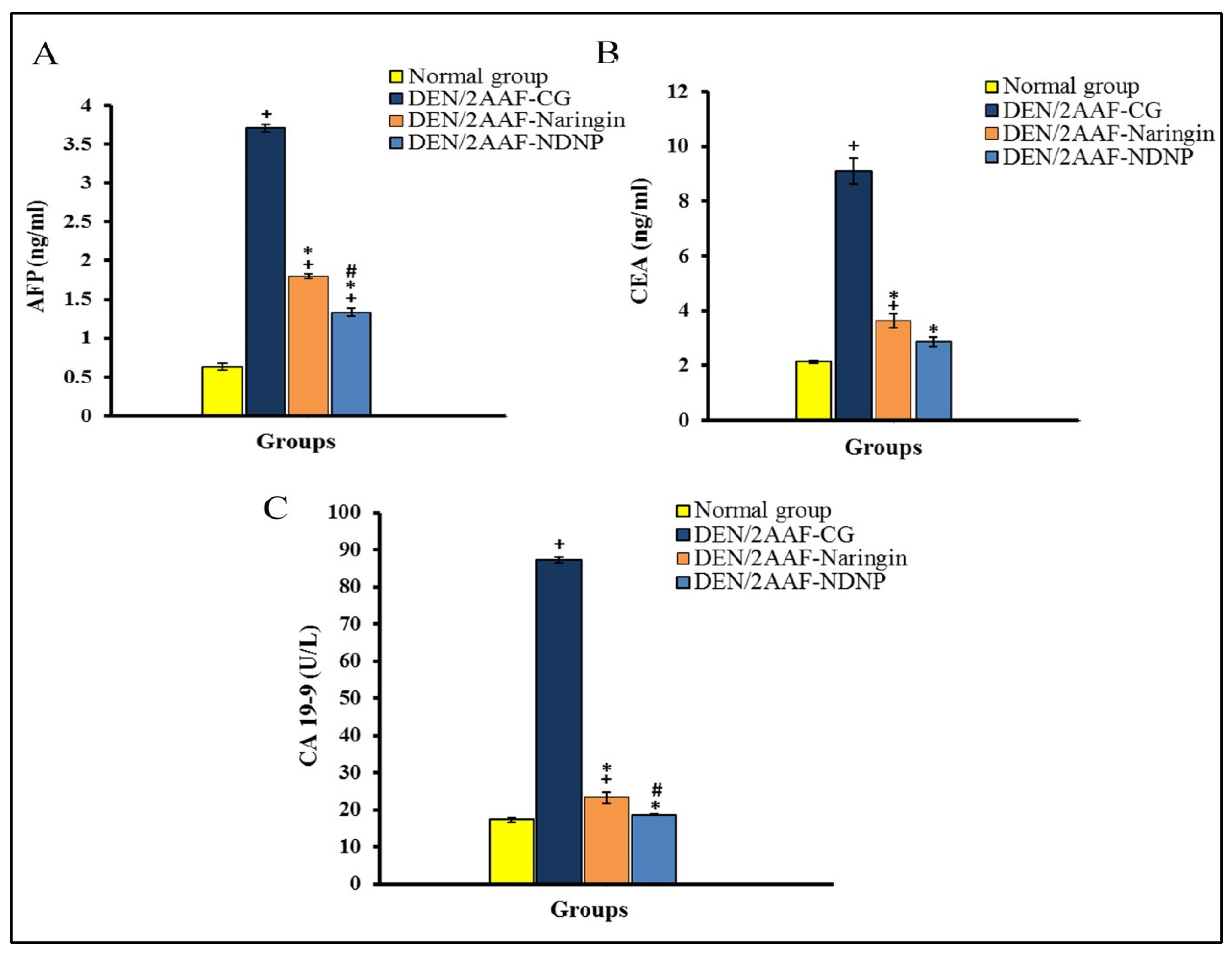
2.2. Effect of Free Naringin and NDNP on DEN/2AAF-Induced Liver Damage
2.3. Effect of Free Naringin and NDNP on DEN/2AAF-Induced Changes in Hepatic Oxidative Stress and Antioxidant Activities
2.4. Effect of Free Naringin and NDNP on DEN/2AAF-Induced Changes in Hepatic Tumor Necrosis Factor Alpha (TNF-α), Interleukin 1 Beta (IL-1β), and Nuclear Factor E2-Related Factor 2 (NRF2) Levels
2.5. Effect of Free Naringin and NDNP on DEN/2AAF-Induced Changes in Hepatic Nuclear Factor-Kappa B (NF-κB) and Interleukin-8 (IL-8) Expressions
2.6. Effects of Free Naringin and NDNP on mRNA Expression of Antiapoptotic and Proapoptotic Biomarkers
2.7. Effects of Free Naringin and NDNP on DEN/2AAF-Induced Changes in Hepatic Isoleucine–Glutamine Motif-Containing GTPase-Activating Protein 1, 2 and 3 (IQGAP1, IQGAP2, and IQGAP3) Expressions
2.8. Effects of Free Naringin and NDNP on DEN/2AAF-Induced Changes in Hepatic Harvey Rat Sarcoma Viral Oncogene Homolog (HRAS) and Kirsten Rat Sarcoma Viral Oncogene Homolog (KRAS) Expressions
2.9. Effects of Free Naringin and NDNP on DEN/2AAF-Induced Changes in Hepatic Proliferator Protein (Ki-67) Expression
2.10. Improvements in DEN/2AAF-Induced Liver Histopathology by Free Naringin and NDNP
3. Materials and Methods
3.1. Chemicals
3.2. Preparation and Characterization of Dextrin–Naringin Nanoparticles
3.3. Experimental Animals
3.4. Experimental Design and Animal Grouping
3.5. Blood and Liver Tissue Sampling
3.6. Detection of Serum Liver Function Parameters
3.7. Detection of Tumor Markers Levels in Serum
3.8. Detection of Hepatic Oxidative Stress and Antioxidant Status Estimation
3.9. Detection of Hepatic TNF-α, IL1β, and NRF2
3.10. Isolation of Total RNA and Reverse Transcription Quantitative PCR (RT-qPCR)
3.11. Western Blot Analysis
3.12. Histopathological Investigation
3.13. Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saber, S.; Mahmoud, A.; Goda, R.; Helal, N.S.; El-Ahwany, E.; Abdelghany, R.H. Perindopril, fosinopril and losartan inhibited the progression of diethylnitrosamine-induced hepatocellular carcinoma in mice via the inactivation of nuclear transcription factor kappa-B. Toxicol. Lett. 2018, 295, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Saber, S.; Khodir, A.E.; Soliman, W.E.; Salama, M.M.; Abdo, W.S.; Elsaeed, B.; Nader, K.; Abdelnasser, A.; Megahed, N.; Basuony, M.; et al. Telmisartan attenuates N-nitrosodiethylamine-induced hepatocellular carcinoma in mice by modulating the NF-κB-TAK1-ERK1/2 axis in the context of PPARγ agonistic activity. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2019, 392, 1591–1604. [Google Scholar] [CrossRef] [PubMed]
- Younis, N.S.; Ghanim, A.M.; Saber, S. Mebendazole augments sensitivity to sorafenib by targeting MAPK and BCL-2 signalling in n-nitrosodiethylamine-induced murine hepatocellular carcinoma. Sci. Rep. 2019, 9, 19095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seydi, E.; Motallebi, A.; Dastbaz, M.; Dehghan, S.G.; Salimi, A.; Nazemi, M.; Pourahmad, J. Selective toxicity of Persian Gulf sea cucumber (Holothuria parva) and sponge (Haliclona oculata) methanolic extracts on liver mitochondria isolated from an animal model of hepatocellular carcinoma. Hepat Mon. 2015, 15, e33073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asafo-Agyei, K.O.; Samant, H. Hepatocellular Carcinoma; StatPearls Publishing LLC, National Center for Biotechnology Information, U.S. National Library of Medicine: Bethesda, MD, USA, 2021. [Google Scholar]
- Sivalingam, K.; Amirthalingam, V.; Ganasan, K.; Huang, C.Y.; Viswanadha, V.P. Neferine suppresses diethylnitrosamine-induced lung carcinogenesis in Wistar rats. Food Chem. Toxicol. 2019, 123, 385–398. [Google Scholar] [CrossRef]
- Hidajat, M.; McElvenny, D.M.; Ritchie, P.; Darnton, A.; Mueller, W.; Agius, R.M.; Cherrie, J.W.; De Vocht, F. Lifetime cumulative exposure to rubber dust, fumes and N-nitrosamines and non-cancer mortality: A 49-year follow-up of UK rubber factory workers. Occup. Environ. Med. 2020, 77, 316–323. [Google Scholar] [CrossRef] [Green Version]
- Arboatti, A.S.; Lambertucci, F.; Sedlmeier, M.G.; Pisani, G.; Monti, J.; Álvarez, M.D.; Frances, D.E.; Ronco, M.T.; Carnovale, C.E. Diethylnitrosamine increases proliferation in early stages of hepatic carcinogenesis in insulin-treated type 1 diabetic mice. BioMed Res. Inter. 2018, 2018, 9472939. [Google Scholar] [CrossRef] [Green Version]
- Alsahli, M.A.; Almatroodi, S.A.; Almatroudi, A.; Khan, A.A.; Anwar, S.; Almutary, A.G.; Alrumaihi, F.; Rahmani, A.H. 6-Gingerol, a major ingredient of ginger attenuates diethylnitrosamine-induced liver injury in rats through the modulation of oxidative stress and anti-inflammatory activity. Mediat. Inflame 2021, 2021, 6661937. [Google Scholar] [CrossRef]
- Senthil, K.; Aranganathan, S.; Nalini, N. Evidence of oxidative stress in the circulation of ovarian cancer patients. Clin. Chim. Acta 2004, 339, 27–32. [Google Scholar] [CrossRef]
- Panieri, E.; Santoro, M.M. ROS homeostasis and metabolism: A dangerous liaison in cancer cells. Cell Death Dis. 2016, 7, 2253. [Google Scholar] [CrossRef]
- Schumacker, P.T. Reactive oxygen species in cancer cells: Live by the sword, die by the sword. Cancer Cell 2006, 10, 175–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tornesello, M.L.; Buonaguro, L.; Izzo, F.; Buonaguro, F.M. Molecular alterations in hepatocellular carcinoma associated with hepatitis B and hepatitis C infections. Oncotarget 2016, 7, 25087–25102. [Google Scholar] [CrossRef] [PubMed]
- Okada, M.; Shibuya, K.; Sato, A.; Seino, S.; Watanabe, E.; Suzuki, S.; Kitanaka, C. Specific role of JNK in the maintenance of the tumor-initiating capacity of A549 human non-small cell lung cancer cells. Oncol. Rep. 2013, 30, 1957–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Wu, L.; Yan, G.; Chen, Y.; Zhou, M.; Wu, Y.; Li, Y. Inflammation and tumor progression: Signaling pathways and targeted intervention. Signal Transduct. Target Ther. 2021, 6, 1–46. [Google Scholar]
- Carneiro, B.A.; El-Deiry, W.S. Targeting apoptosis in cancer therapy. Nat. Rev. Clin. Oncol. 2020, 17, 395–417. [Google Scholar] [CrossRef]
- Ryan, M.B.; Corcoran, R.B. Therapeutic strategies to target RAS-mutant cancers. Nat. Rev. Clin. Oncol. 2018, 15, 709–720. [Google Scholar] [CrossRef]
- Diepstraten, S.T.; Anderson, M.A.; Czabotar, P.E.; Lessene, G.; Strasser, A.; Kelly, G.L. The manipulation of apoptosis for cancer therapy using BH3-mimetic drugs. Nat. Rev. Cancer 2022, 22, 45–64. [Google Scholar] [CrossRef]
- Adiban, H.; Shirazi, F.H.; Gholami, S.; Kamalinejad, M.; Hosseini, S.H.; Noubarani, M.; Eskandari, M.R. Chemopreventive effect of quince (Cydonia oblonga Mill.) fruit extract on hepatocellular carcinoma induced by diethylnitrosamine in rats. Inter. Pharm. Acta. 2019, 2, 2e2-1. [Google Scholar]
- Shirazi, F.H.; Piri, M.; Keshavarz, S.; Gholami, S.; Hosseini, S.H.; Noubarani, M.; Andalib, S.; Kamalinejad, M.; Adiban, H.; Eskandari, M.R. Olive fruit (Olea europaea L.): Chemopreventive effect in the rat model of hepatocellular carcinoma. Pharma. Nut. 2018, 6, 207–214. [Google Scholar] [CrossRef]
- Camargo, C.A.; Gomes-Marcondes, M.C.; Wutzki, N.C.; Aoyama, H. Naringin inhibits tumor growth and reduces interleukin-6 and tumor necrosis factor α levels in rats with Walker 256 carcinosarcoma. Anticancer Res. 2012, 32, 129–134. [Google Scholar]
- Sun, J.; Kormakov, S.; Liu, Y.; Huang, Y.; Wu, D.; Yang, Z. Recent progress in metal-based nanoparticles mediated photodynamic therapy. Molecules 2018, 23, 1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Chen, Z.; Hu, Y.; Rao, Z.; Wu, W.; Yang, Z. Nanocrystals: The preparation, precise control and application toward the pharmaceutics and food industry. Curr. Pharm. Des. 2018, 24, 2425–2431. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, D.; Tuli, H.S.; Yerer, M.B.; Sharma, A.; Sak, K.; Srivastava, S.; Pandey, A.; Garg, V.K.; Sethi, G.; Bishayee, A. Natural product-based nanoformulations for cancer therapy: Opportunities and challenges. Semin. Cancer Biol. 2021, 69, 5–23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.N.; Yu, R.S.; Xu, M.; Cheng, X.Y.; Chen, C.M.; Xu, X.L.; Lu, C.Y.; Lu, K.J.; Chen, M.J.; Zhu, M.L.; et al. Visual targeted therapy of hepatic cancer using homing peptide modified calcium phosphate nanoparticles loading doxorubicin guided by T1 weighted MRI. Nanomed. Nanotech. Biol. Med. 2018, 14, 2167–2178. [Google Scholar] [CrossRef] [PubMed]
- Shamay, Y.; Shah, J.; Işık, M.; Mizrachi, A.; Leibold, J.; Tschaharganeh, D.F.; Roxbury, D.; Budhathoki-Uprety, J.; Nawaly, K.; Sugarman, J.L.; et al. Quantitative self-assembly prediction yields targeted nanomedicines. Nat. Mater. 2018, 17, 361–368. [Google Scholar] [CrossRef]
- Mohamed, E.E.; Abdel-Moneim, A.; Ahmed, O.M.; Zoheir, K.M.; Eldin, Z.E.; El-Shahawy, A.A. Anticancer activity of a novel naringin–dextrin nanoformula: Preparation, characterization, and in vitro induction of apoptosis in human hepatocellular carcinoma cells by inducing ROS generation, DNA fragmentation, and cell cycle arrest. J. Drug Deliv. Sci. Tech. 2022, 75, 103677. [Google Scholar] [CrossRef]
- Manchun, S.; Dass, C.R.; Sriamornsak, P. Designing nanoemulsion templates for fabrication of dextrin nanoparticles via emulsion cross-linking technique. Carbohy. Polym. 2014, 101, 650–655. [Google Scholar] [CrossRef]
- Ahmed, O.M.; Fahim, H.I.; Mohamed, E.E.; Abdel-Moneim, A. Protective effects of Persea americana fruit and seed extracts against chemically induced liver cancer in rats by enhancing their antioxidant, anti-inflammatory, and apoptotic activities. Environ. Sci. Pollut. Res. 2022, 4, 1–6. [Google Scholar] [CrossRef]
- Gella, F.J.; Olivella, T.; Cruz, P.M.; Arenas, J.; Moreno, R.; Durban, R.; Gomez, J.A. A simple procedure for routine determination of aspartate aminotransferase and alanine aminotransferase with pyridoxal phosphate. Clin. Chim. Acta 1985, 153, 241–247. [Google Scholar] [CrossRef]
- Schumann, G.; Bonora, R.; Ceriotti, F.; Férard, G.; Ferrero, C.; Franck, P.F.; Gella, F.J.; Hoelzel, W.; Jørgensen, P.J.; Kanno, T.; et al. IFCC primary reference procedures for the measurement of catalytic activity concentrations of enzymes at 37 °C. Part 6: Reference procedure for the measurement of catalytic concentration of γ-Glutamyltransferase. Clin. Chem. Lab. Med. 2002, 40, 734–738. [Google Scholar]
- Jendrassik, L. Colorimetric determination of bilirubin. Biochemistry 1938, 97, 72–81. [Google Scholar]
- Doumas, B.T.; Watson, W.A.; Biggs, H.G. Albumin standards and the measurement of serum albumin with bromcresol green. Clin. Chim. Acta 1971, 31, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Sthoeger, Z.; Zinger, H.; Sharabi, A.; Asher, I.; Mozes, E. The tolerogenic peptide, hCDR1, down-regulates the expression of interferon-α in murine and human systemic lupus erythematosus. PLoS ONE 2013, 8, e60394. [Google Scholar] [CrossRef] [PubMed]
- He, F. Bradford Protein Assay. Bio-Protocol 2011, 101, e45. [Google Scholar] [CrossRef] [Green Version]
- Tawfik, N.G.; Mohamed, W.R.; Mahmoud, H.S.; Alqarni, M.A.; Naguib, I.A.; Fahmy, A.M.; Ahmed, O.M. Isatin Counteracts Diethylnitrosamine/2-Acetylaminofluorene-Induced Hepatocarcinogenesis in Male Wistar Rats by Upregulating Anti-Inflammatory, Antioxidant, and Detoxification Pathways. Antioxidant 2022, 11, 699. [Google Scholar] [CrossRef]
- Yassin, N.; AbouZid, S.F.; El-Kalaawy, A.M.; Ali, T.M.; Elesawy, B.H.; Ahmed, O.M. Tackling of renal carcinogenesis in Wistar rats by Silybum marianum total extract, silymarin, and silibinin via modulation of oxidative stress, apoptosis, Nrf2, PPAR, NF-κB, and PI3K/Akt signaling pathways. Oxidative Med. Cell. Longev. 2021, 2021, 7665169. [Google Scholar] [CrossRef]
- Bancroft, J.D.; Gamble, M. (Eds.) Theory and practice of histological techniques. In Elsevier Health Sciences, 6th ed.; Churchill Livingstone: Beijing, China, 2008. [Google Scholar]
- Sperandio, R.C.; Pestana, R.C.; Miyamura, B.V.; Kaseb, A.O. Hepatocellular carcinoma immunotherapy. Annu. Rev. Med. 2022, 73, 267–278. [Google Scholar]
- Mohanty, S.; Konkimalla, V.B.; Pal, A.; Sharma, T.; Si, S.C. Naringin as Sustained Delivery Nanoparticles Ameliorates the Anti-inflammatory Activity in a Freund’s Complete Adjuvant-Induced Arthritis Model. ACS Omega 2021, 6, 28630–28641. [Google Scholar] [CrossRef]
- Ahmed, O.M.; Ahmed, A.A.; Fahim, H.I.; Zaky, M.Y. Quercetin and naringenin abate diethylnitrosamine/acetylaminofluorene-induced hepatocarcinogenesis in Wistar rats: The roles of oxidative stress, inflammation and cell apoptosis. Drug Chem. Toxicol. 2019, 45, 262–273. [Google Scholar] [CrossRef]
- Shahin, Y.R.; Elguindy, N.M.; Abdel Bary, A.; Balbaa, M. The protective mechanism of Nigella sativa against diethylnitrosamine-induced hepatocellular carcinoma through its antioxidant effect and EGFR/ERK1/2 signaling. Environ. Toxicol. 2018, 33, 885–898. [Google Scholar] [CrossRef]
- Acharya, R.; Mishra, N.; Kumar, A.; Bose, P.; Pattnaik, A.; Mukhopadhyay, K.; Sunita, P.; Pattanayak, S.P. Naringin, a natural flavonone glycoside attenuates N-nitrosodiethylamine-induced hepatocellular carcinoma in Sprague-Dawley rats. Pharma. Mag. 2021, 17, 196–204. [Google Scholar]
- Ratheesh, G.; Tian, L.; Venugopal, J.R.; Ezhilarasu, H.; Sadiq, A.; Fan, T.P.; Ramakrishna, S. Role of medicinal plants in neurodegenerative diseases. Biomanufactur. Rev. 2017, 2, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Park, S.J.; Jang, J.Y.; Jeong, S.W.; Cho, Y.K.; Lee, S.H.; Kim, S.G.; Cha, S.W.; Kim, Y.S.; Cho, Y.D.; Kim, H.S.; et al. Usefulness of AFP, AFP-L3, and PIVKA-II, and their combinations in diagnosing hepatocellular carcinoma. Medicine 2017, 96, e5811. [Google Scholar] [CrossRef] [PubMed]
- Edoo, M.I.A.; Chutturghoon, V.K.; Wusu-Ansah, G.K.; Hai, Z.H.U.; Zhen, T.Y.; Xie, H.Y.; Zheng, S.S. Serum biomarkers AFP, CEA and CA19-9 combined detection for early diagnosis of hepatocellular carcinoma. Iran J. Public Health 2016, 48, 314. [Google Scholar]
- Mansour, D.F.; Abdallah, H.M.; Ibrahim, B.M.; Hegazy, R.R.; Esmail, R.S.; Abdel-Salam, L.O. The carcinogenic agent diethylnitrosamine induces early oxidative stress, inflammation and proliferation in rat liver, stomach and colon: Protective effect of ginger extract. Asian Pacific. J. Cancer Prev. 2019, 20, 2551–2561. [Google Scholar] [CrossRef]
- Gunasekaran, S.; Mayakrishnan, V.; Al-Ghamdi, S.; Alsaidan, M.; Geddawy, A.; Abdelaziz, M.A.; Mohideen, A.P.; Bahakim, N.O.; Ramesh, T.; Ayyakannu, U.R. Investigation of phytochemical profile and in vivo anti-proliferative effect of Laetiporus versisporus (Lloyd) Imazeki mushroom against diethylnitrosamine-induced hepatocellular carcinoma. J. King Saud Univ. Sci. 2021, 33, 101551. [Google Scholar] [CrossRef]
- Tao, L.Y.; Cai, L.; He, X.D.; Liu, W.; Qu, Q. Comparison of serum tumor markers for intrahepatic cholangiocarcinoma and hepatocellular carcinoma. Amer. Sur. 2010, 76, 1210–1213. [Google Scholar] [CrossRef]
- Oliveira, M.M.; Teixeira, J.C.; Vasconcelos-Nóbrega, C.; Felix, L.M.; Sardão, V.A.; Colaço, A.A.; Oliveira, P.A.; Peixoto, F.P. Mitochondrial and liver oxidative stress alterations induced by N-butyl-N-(4-hydroxybutyl) nitrosamine: Relevance for hepatotoxicity. J. Appl. Toxicol. 2013, 33, 434–443. [Google Scholar] [CrossRef]
- Verma, A.; Singh, D.; Anwar, F.; Bhatt, P.C.; Al-Abbasi, F.; Kumar, V. Triterpenoids principle of Wedelia calendulacea attenuated diethynitrosamine-induced hepatocellular carcinoma via down-regulating oxidative stress, inflammation and pathology via NF-kB pathway. Inflammopharmacol 2018, 26, 133–146. [Google Scholar] [CrossRef]
- Ahmed, O.M. Relationships between oxidative stress, cancer development and therapeutic interventions. J. Can. Sci. Res. 2016, 1, 1e104. [Google Scholar] [CrossRef]
- Yassin, N.Y.; AbouZid, S.F.; El-Kalaawy, A.M.; Ali, T.M.; Almehmadi, M.M.; Ahmed, O.M. Silybum marianum total extract, silymarin and silibinin abate hepatocarcinogenesis and hepatocellular carcinoma growth via modulation of the HGF/c-Met, Wnt/β-catenin, and PI3K/Akt/mTOR signaling pathways. Biomed. Pharmacother. 2022, 145, 112409. [Google Scholar] [CrossRef] [PubMed]
- Mo’men, Y.S.; Hussein, R.M.; Kandeil, M.A. A novel chemoprotective effect of tiopronin against diethylnitrosamine-induced hepatocellular carcinoma in rats: Role of ASK1/P38 MAPK-P53 signalling cascade. Clin. Exp. Pharmacol. Physiol. 2020, 47, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, I.A.; Rocha, J.; Sepodes, B.; Mota-Filipe, H.; Ribeiro, M.H. Effect of naringin enzymatic hydrolysis towards naringenin on the anti-inflammatory activity of both compounds. J. Mol. Catal. B Enzym. 2008, 52, 13–18. [Google Scholar] [CrossRef]
- Thangavel, P.; Muthu, R.; Vaiyapuri, M. Antioxidant potential of naringin–A dietary flavonoid–In N-Nitrosodiethylamine induced rat liver carcinogenesis. Biomed. Prev. Nutr. 2012, 2, 193–202. [Google Scholar] [CrossRef]
- Adebiyi, O.O.; Adebiyi, O.A.; Owira, P.M. Naringin reverses hepatocyte apoptosis and oxidative stress associated with HIV-1 nucleotide reverse transcriptase inhibitors-induced metabolic complications. Nutrients 2015, 7, 10352–10368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oluwafeyisetan, A.; Olubunmi, A.; Peter, O. Naringin ameliorates HIV-1 nucleoside reverse transcriptase inhibitors-induced mitochondrial toxicity. Current HIV Res. 2016, 14, 506–516. [Google Scholar] [CrossRef]
- Ali, G.; Omar, H.; Hersi, F.; Abo-Youssef, A.; Ahmed, O.; Mohamed, W. The protective role of etoricoxib against diethylnitrosamine/2-acetylaminofluorene-induced hepatocarcinogenesis in Wistar rats: The impact of NF-κB/COX-2/PGE2 signaling. Curr. Mol. Pharmacol. 2022, 15, 252–262. [Google Scholar]
- Zoheir, K.M.; Abdelhafez, M.A.; Darwish, A.M.; Mahrous, K.F. New Approach about the Signaling Crosstalk between IQGAPs/NF-κB/IL-8 and PDCD5/p53/TRAIL Pathways that Modulate Malignant Transformation in Hepatocellular Carcinoma. Asian Pac. J. Cancer Prev. 2022, 23, 271–279. [Google Scholar]
- Uehara, T.; Pogribny, I.P.; Rusyn, I. The DEN and CCl4-induced mouse model of fbrosis and infammation-associated hepatocellular carcinoma. Curr. Protoc. Pharmacol. 2014, 66, 14–30. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Chaudhary, D.; Kumar, V.; Verma, A. Amelioration of diethylnitrosamine (DEN) induced renal oxidative stress and inflammation by Carissa carandas embedded silver nanoparticles in rodents. Toxico. Rep. 2021, 8, 636–645. [Google Scholar] [CrossRef]
- Ahmed, O.M.; Fahim, H.I.; Ahmed, H.Y.; Al-Muzafar, H.M.; Ahmed, R.R.; Amin, K.A.; El-Nahass, E.S.; Abdelazeem, W.H. The preventive effects and the mechanisms of action of navel orange peel hydroethanolic extract, naringin, and naringenin in N-acetyl-p-aminophenol-induced liver injury in Wistar rats. Oxid. Med. Cell. Longev. 2019, 2019, 2745352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, H.; Wang, H.; Wang, X.; Zhu, L.; Mao, L. The absence of Nrf2 enhances NF-κBdependent inflammation following scratch injury in mouse primary cultured astrocytes. Mediat. Inflamm. 2012, 2012, 217580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.L.; Tsai, J.R.; Hwang, J.J.; Chou, S.H.; Cheng, Y.J.; Lin, F.Y.; Chen, Y.L.; Hung, C.Y.; Chen, W.C.; Chen, Y.H.; et al. High-Mobility Group Box 1–Mediated Matrix Metalloproteinase-9 Expression in Non–Small Cell Lung Cancer Contributes to Tumor Cell Invasiveness. Am. J. Resp. Cell Mol. Biol. 2010, 43, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Caglayan, C.; Temel, Y.; Kandemir, F.M.; Yildirim, S.; Kucukler, S. Naringin protects against cyclophosphamide-induced hepatotoxicity and nephrotoxicity through modulation of oxidative stress, inflammation, apoptosis, autophagy, and DNA damage. Environ. Sci. Pollut. Res. Int. 2018, 25, 20968–20984. [Google Scholar] [CrossRef]
- Wu, D.; Wu, P.; Zhao, L.; Huang, L.; Zhang, Z.; Zhao, S.; Huang, J. NF-κB expression and outcomes in solid tumors: A systematic review and meta-analysis. Medicine 2015, 94, e1687. [Google Scholar] [CrossRef]
- Shirani, K.; Yousefsani, B.S.; Shirani, M.; Karimi, G. Protective effects of naringin against drugs and chemical toxins induced hepatotoxicity: A review. Phytothe. Res. 2020, 34, 1734–1744. [Google Scholar] [CrossRef]
- Lv, Z.; Wu, W.; Ge, S.; Jia, R.; Lin, T.; Yuan, Y.; Kuang, H.; Yang, B.; Wu, L.; Wei, J.; et al. Naringin protects against perfluorooctane sulfonate-induced liver injury by modulating NRF2 and NF-κB in mice. Intern. Immunoph. 2018, 65, 140–147. [Google Scholar] [CrossRef]
- Dong, D.; Xu, L.; Yin, L.; Qi, Y.; Peng, L. Naringin prevents carbon tetrachloride-induced acute liver injury in mice. J. Funct. Foods 2015, 12, 179–191. [Google Scholar] [CrossRef]
- Manna, K.; Khan, A.; Biswas, S.; Das, U.; Sengupta, A.; Mukherjee, D.; Chakraborty, A.; Dey, S. Naringin ameliorates radiation-induced hepatic damage through modulation of Nrf2 and NF-κB pathways. RSC. Adv. 2016, 6, 23058–23073. [Google Scholar] [CrossRef]
- Saraf, S. Applications of novel drug delivery system for herbal formulations. Fitoterapia 2010, 81, 680–689. [Google Scholar]
- Nair, H.B.; Sung, B.; Yadav, V.R.; Kannappan, R.; Chaturvedi, M.M.; Aggarwal, B.B. Delivery of antiinflammatorynutraceuticals by nanoparticles for the prevention and treatment of cancer. Biochem. Pharmacol. 2010, 80, 1833–1843. [Google Scholar] [CrossRef] [PubMed]
- XinYou, S.; JianQiang, Z.; Nan, L.; Kumar, M.; AiMeiOu, Y. Chemoprotective effects of resveratrol against diethylnitrosamine induced hepatocellular carcinoma in Wistar rats. Inter. J. Pharma. 2019, 15, 549–559. [Google Scholar]
- Medhat, A.; Mansour, S.; El-Sonbaty, S.; Kandil, E.; Mahmoud, M. Evaluation of the antitumor activity of platinum nanoparticles in the treatment of hepatocellular carcinoma induced in rats. Tumor Biol. 2017, 39, 1010428317717259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramaniam, N.; Kannan, P.; Thiruvengadam, D. Hepatoprotective effect of boldine against diethylnitrosamine-induced hepatocarcinogenesis in wistar rats. J. Biochem. Mol. Toxicol. 2019, 33, e22404. [Google Scholar]
- Kang, M.H.; Reynolds, C.P. Bcl-2 inhibitors: Targeting mitochondrial apoptotic pathways in cancer therapy. Clin. Cancer Res. 2009, 15, 1126–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rückert, F.; Samm, N.; Lehner, A.K.; Saeger, H.D.; Grützmann, R.; Pilarsky, C. Simultaneous gene silencing of Bcl-2, XIAP and Survivin re-sensitizes pancreatic cancer cells towards apoptosis. BMC Cancer 2020, 10, 379–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, K.J.; Tait, S.W. Targeting BCL-2 regulated apoptosis in cancer. Open Biol. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Marquez, R.T.; Tsao, B.W.; Faust, N.F.; Xu, L. Drug resistance and molecular cancer therapy: Apoptosis versus autophagy. Apoptosis 2013, 2013, 155–177. [Google Scholar]
- Ou, X.H.; Lu, Y.; Liao, L.F.; Li, D.N.; Liu, L.M.; Liu, H.G.; Xu, H. Nitidine chloride induces apoptosis in human hepatocellular carcinoma cells through a pathway involving p53, p21, Bax and Bcl-2. Oncol. Rep. 2015, 33, 1264–1274. [Google Scholar]
- Cui, X.; Choi, H.K.; Choi, Y.S.; Park, S.Y.; Sung, G.J.; Lee, Y.H.; Lee, J.; Jun, W.J.; Kim, K.; Choi, K.C.; et al. DNAJB1 destabilizes PDCD5 to suppress p53-mediated apoptosis. Cancer Lett. 2015, 357, 307–315. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, N.; Ye, X.; Li, H.; Feng, Y.; Cheung, F.; Nagamatsu, T. Hepatoprotective effect and its possible mechanism of Coptidis rhizoma aqueous extract on carbon tetrachloride-induced chronic liver hepatotoxicity in rats. J. Ethnopharmacol. 2011, 138, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.; Yuan, P.; Wang, D.; Jin, H.; Chen, H. Effects of naringin on the expression of miR-19b and cell apoptosis in human hepatocellular carcinoma. Oncol. Lett. 2017, 14, 1455–1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuge, C.; Sun, X.; Chen, Y.; Lei, J. PDCD5 functions as a regulator of p53 dynamics in the DNA damage response. J. Theoret. Biol. 2016, 388, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Borude, P.; Bhushan, B.; Chavan, H.; Weemhoff, J.L.; Jaeschke, H.; Krishnamurthy, P.; Apte, U. P53 regulates progression of injury and liver regeneration after acetaminophen overdose. FASEB J. 2017, 31, 531–538. [Google Scholar]
- Ghanbari-Movahed, M.; Jackson, G.; Farzaei, M.H.; Bishayee, A. A systematic review of the preventive and therapeutic effects of naringin against human malignancies. Front. Pharmacol. 2021, 12, 639840. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, S.; Marais, R.; Zhu, A.X. The role of signaling pathways in the development and treatment of hepatocellular carcinoma. Oncogene 2010, 29, 4989–5005. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, V.A. Watch the GAP: Emerging Roles for IQ Motif-Containing GTPase-Activating Proteins IQGAPs in Hepatocellular Carcinoma. Int. J. Hepatol. 2012, 2012, 958673. [Google Scholar] [CrossRef] [Green Version]
- Zoheir, K.; Abd-Rabou, A.A.; Harisa, G.I.; Kumar, A.; Ahmad, S.F.; Ansari, M.A.; Abd-Allah, A.R. IQGAP1 gene silencing induces apoptosis and decreases the invasive capacity of human hepatocellular carcinoma cells. Tumor Biol. 2016, 37, 13927–13939. [Google Scholar] [CrossRef]
- Johnson, M.; Sharma, M.; Henderson, B.R. IQGAP1 regulation and roles in cancer. Cell Signal. 2009, 21, 1471–1478. [Google Scholar]
- White, M.J.; McArthur, K.; Metcalf, D.; Lane, R.M.; Cambier, J.C.; Herold, M.J.; Van Delft, M.F.; Bedoui, S.; Lessene, G.; Ritchie, M.E.; et al. Apoptotic caspases suppress mtDNA-induced STING-mediated type I IFN production. Cell 2014, 159, 1549–1562. [Google Scholar] [CrossRef] [Green Version]
- Gnatenko, D.V.; Xu, X.; Zhu, W.; Schmidt, V.A. Transcript profiling identifies iqgap2−/− mouse as a model for advanced human hepatocellular carcinoma. PLoS ONE 2013, 8, e71826. [Google Scholar] [CrossRef] [PubMed]
- Skawran, B.; Steinemann, D.; Weigmann, A.; Flemming, P.; Becker, T.; Flik, J.; Kreipe, H.; Schlegelberger, B.; Wilkens, L. Gene expression profiling in hepa- tocellular carcinoma: Upregulation of genes in amplified chromosome regions. Mod. Pathol. 2008, 21, 505–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Zhao, W.; Xu, Q.W.; Wang, X.S.; Zhang, Y.; Zhang, J. IQGAP3 promotes EGFR-ERK signaling and the growth and metastasis of lung cancer cells. PLoS ONE 2014, 9, e97578. [Google Scholar] [CrossRef] [Green Version]
- Qian, E.N.; Han, S.Y.; Ding, S.Z.; Lv, X. Expression and diagnostic value of CCT3 and IQGAP3 in hepatocellular carcinoma. Cancer Cell Inter. 2016, 16, 55–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Qin, N.; Zhou, Q.; Chen, Y.; Huang, S.; Chen, B.; Shen, G.; Jia, H. Role of IQGAP3 in metastasis and epithelial–mesenchymal transition in human hepatocellular carcinoma. J. Transl. Med. 2017, 15, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Chen, Z.; Cao, H.; Yu, Z.; Feng, J.; Wang, K.; Lu, Q.; Wu, Y. High expression of IQGAP3 indicates poor prognosis in colorectal cancer patients. Inter. J. Bio. Markers 2019, 34, 348–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahat, A.A.; Alsaid, M.S.; Kotob, S.E.; Ahmed, H.H. Significance of Rumex vesicarius as anticancer remedy against hepatocellular carcinoma: A proposal-based on experimental animal studies. Asian Pac. J. Cancer Prev. 2015, 16, 4303–4310. [Google Scholar] [CrossRef]
- Newell, P.; Villanueva, A.; Friedman, S.L.; Koike, K.; Llovet, J.M. Experimental models of hepatocellular carcinoma. J. Hepatol. 2008, 48, 858–879. [Google Scholar] [CrossRef] [Green Version]
- Dhillon, A.S.; Hagan, S.; Rath, O.; Kolch, W. MAP kinase signalling pathways in cancer. Oncogene 2007, 26, 3279–3290. [Google Scholar] [CrossRef] [Green Version]
- Castellano, E.; Downward, J. RAS interaction with PI3K: More than just another effector pathway. Genes Cancer 2011, 2, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Zoheir, K.M.; Abd-Rabou, A.A.; Harisa, G.I.; Ashour, A.E.; Ahmad, S.F.; Attia, S.M.; Bakheet, S.A.; Abdel-Hamied, H.E.; Abd-Allah, A.R.; Kumar, A. Gene expression of IQGAPs and Ras families in an experimental mouse model for hepatocellular carcinoma: A mechanistic study of cancer progression. Int. J. Clin. Exp. Pathol. 2015, 8, 8821–8831. [Google Scholar] [PubMed]
- Cao, Y.; Ke, R.; Wang, S.; Zhu, X.; Chen, J.; Huang, C.; Jiang, Y.; Lv, L. EDNA topoisomerase IIα and Ki67 are prognostic factors in patients with hepatocellular carcinoma. Oncol. Lett. 2017, 13, 4109–4116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jagetia, G.C.; Reddy, T.K. Modulation of radiationinduced alteration in the antioxidant status of mice by naringin. Life Sci. 2005, 77, 780–794. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Prakash, A.; Dogra, S. Naringin alleviates cognitive impairment, mitochondrial dysfunction and oxidative stress induced by D-galactose in mice. Food Chem. Toxicol. 2010, 48, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Thangavel, P.; Vaiyapuri, M. Antiproliferative and apoptotic effects of naringin on diethylnitrosamine induced hepatocellular carcinoma in rats. Biomed. Aging Pathol. 2013, 3, 59–64. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | ALT | AST | ALP | Total Bilirubin | Albumin |
|---|---|---|---|---|---|
| (U/L) | (U/L) | (U/L) | (mg/dL) | (g/dL) | |
| Normal group | 40.40 ± 3.05 | 97.00 ± 1.52 | 206.08 ± 6.45 | 0.27 ± 0.02 | 3.75 ± 0.11 |
| DEN/2AAF-CG | 70.18 ± 2.68 + | 161.66 ± 2.96 + | 506.28 ± 11.46 + | 0.90 ± 0.07 + | 2.88 ± 0.05 + |
| DEN/2AAF + naringin | 50.31 ± 1.31 +* | 111.16 ± 4.24 +* | 398.66 ± 10.59 +* | 0.48 ± 0.03 +* | 3.32 ± 0.07 +* |
| DEN/2AAF + NDNP | 41.36 ± 1.73 *# | 100.50 ± 2.51 *# | 338.6 ± 10.69 +*# | 0.40 ± 0.04 * | 3.55 ± 0.04 *# |
| Groups | TNF-α | IL-1β | NRF2 |
|---|---|---|---|
| (Pg/mg Tissue) | (Pg/mg Tissue) | (Pg/mg Tissue) | |
| Normal group | 22.53 ± 1.04 | 35.64 ± 1.07 | 259.41 ± 2.81 |
| DEN/2AAF- CG | 126.66 ± 0.93 + | 165.78 ± 0.80 + | 93.80 ± 0.77 + |
| DEN/2AAF + naringin | 55.23 ± 0.78 +* | 74.13 ± 0.74 +* | 219.41 ± 1.2 +* |
| DEN/2AAF + NDNP | 34.00 ± 0.77 +*# | 55.41 ± 0.59 +*# | 236.51 ± 3.12 +*# |
| Histopathological Changes | Score | Normal | DEN/2AAF | DEN/2AAF+ Naringin | DEN/2AAF+ NDNP |
|---|---|---|---|---|---|
| Karyomegalic hepatocytic nuclei with more than one prominent nucleoli | 0 | 6(100%) | - | 5(83.3%) | 6(100%) |
| I | - | - | 1(16.7%) | - | |
| II | - | 1(16.7%) | - | - | |
| III | - | 5(83.3%) | - | - | |
| Steatosis and cytoplasmic vacuolization of hepatocyte | 0 | 6(100%) | - | 2(33.3%) | 4(66.6%) |
| I | - | - | 3 (50%) | 1(16.7%) | |
| II | - | 2(33.3%) | 1(16.7%) | 1(16.7%) | |
| III | - | 4 (66.6%) | - | - | |
| Dysplastic hepatocytes with mitotic figures | 0 | 6(100%) | - | 4(66.6%) | 6(100%) |
| I | - | - | 2(33.3%) | - | |
| II | - | 3(50%) | - | - | |
| III | - | 3(50%) | - | - | |
| Clear hepatocellular focus | 0 | 6(100%) | - | 1(16.7%) | 6(100%) |
| I | - | - | 4(66.6%) | - | |
| II | - | - | 1(16.7%) | - | |
| III | - | 6 (100%) | - | - | |
| Deeply eosinophilic foci of hepatocytes | 0 | 6(100%) | - | 2(100%) | 5(83.8%) |
| I | - | 1(16.7%) | 3(50%) | 1(16.6%) | |
| II | - | 2(33.3%) | 1(16.7) | - | |
| III | - | 3(50%) | - | - | |
| Inflammatory cell infiltration | 0 | 6(100%) | - | 3(50%) | 4(66.6%) |
| I | - | - | 3(50%) | 2(33.3%) | |
| II | - | 1(16.7%) | - | - | |
| III | - | 5(83.3%) | - | - |
| Genes | GenBank Accession Number | Sequence (5′–3′) |
|---|---|---|
| IQGAP1 | NM_001108489.1 | F: GCGGCTTCCAACAAGATGTTT R: CAGCAGTTCATGGATGGGGT |
| IQGAP2 | XM_039103456.1 |
F: GGAATCTACGGACGCTGGAG R: CACAGTACTGGGTGTGTCCC |
| IQGAP3 | NM_001191709.1 |
F: AGCCTATGATCGTCTCACAGC R: CACAGGTACTGGTAGGCGAC |
| NF-κB | NM_001276711.1 |
F: TTCAACATGGCAGACGACGA R: TGCTCTAGTATTTGAAGGTATGGG |
| IL-8 | X77797.1 |
F: CAGAGACTTGGGAGCCACTC R: CAGAGTAAAGGGCGGGTCAG |
| Bcl-2 | NM_016993.1 | F: TAAGCTGTCACAGAGGGGCT R: TGAAGAGTTCCTCCACCACC |
| Bax | NM_007527.3 | F: CTGGATCCAAGACCAGGGTG R: CCTTTCCCCTTCCCCCATTC |
| P53 | NM_030989.3 |
F: GTTTTTGTTCCTGAGCCCCG R: GAGCAAGGGGTGACTTTGGG |
| PDCD5 | NM_001106247.1 |
F: TGAAGCGATTCCAACCGAGT R: GCTCCGTGGGTCTGTCTAAG |
| HRAS | NM_ 001130441.1 | F: CTGTCCTGACACCAGGCTC R: ATGGACCCTCCTGTAGCCAT |
| KRAS | NM_ 031515.3 | F: AGACACGAAACAGGCTCAGG R: GCATCGTCAACACCCTGTCT |
| β-actin | NM_031144.3 | F: TCACTATCGGCAATGTGCGG R: GCTCAGGAGGAGCAATGATG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamed, E.E.; Ahmed, O.M.; Abdel-Moneim, A.; Zoheir, K.M.A.; Elesawy, B.H.; Al Askary, A.; Hassaballa, A.; El-Shahawy, A.A.G. Protective Effects of Naringin–Dextrin Nanoformula against Chemically Induced Hepatocellular Carcinoma in Wistar Rats: Roles of Oxidative Stress, Inflammation, Cell Apoptosis, and Proliferation. Pharmaceuticals 2022, 15, 1558. https://doi.org/10.3390/ph15121558
Mohamed EE, Ahmed OM, Abdel-Moneim A, Zoheir KMA, Elesawy BH, Al Askary A, Hassaballa A, El-Shahawy AAG. Protective Effects of Naringin–Dextrin Nanoformula against Chemically Induced Hepatocellular Carcinoma in Wistar Rats: Roles of Oxidative Stress, Inflammation, Cell Apoptosis, and Proliferation. Pharmaceuticals. 2022; 15(12):1558. https://doi.org/10.3390/ph15121558
Chicago/Turabian StyleMohamed, Eman E., Osama M. Ahmed, Adel Abdel-Moneim, Khairy M. A. Zoheir, Basem H. Elesawy, Ahmad Al Askary, Ahmed Hassaballa, and Ahmed A. G. El-Shahawy. 2022. "Protective Effects of Naringin–Dextrin Nanoformula against Chemically Induced Hepatocellular Carcinoma in Wistar Rats: Roles of Oxidative Stress, Inflammation, Cell Apoptosis, and Proliferation" Pharmaceuticals 15, no. 12: 1558. https://doi.org/10.3390/ph15121558