
Agathis robusta Bark Extract Protects from Renal Ischemia-Reperfusion Injury: Phytochemical, In Silico and In Vivo Studies

 , , and
, , and
Abstract
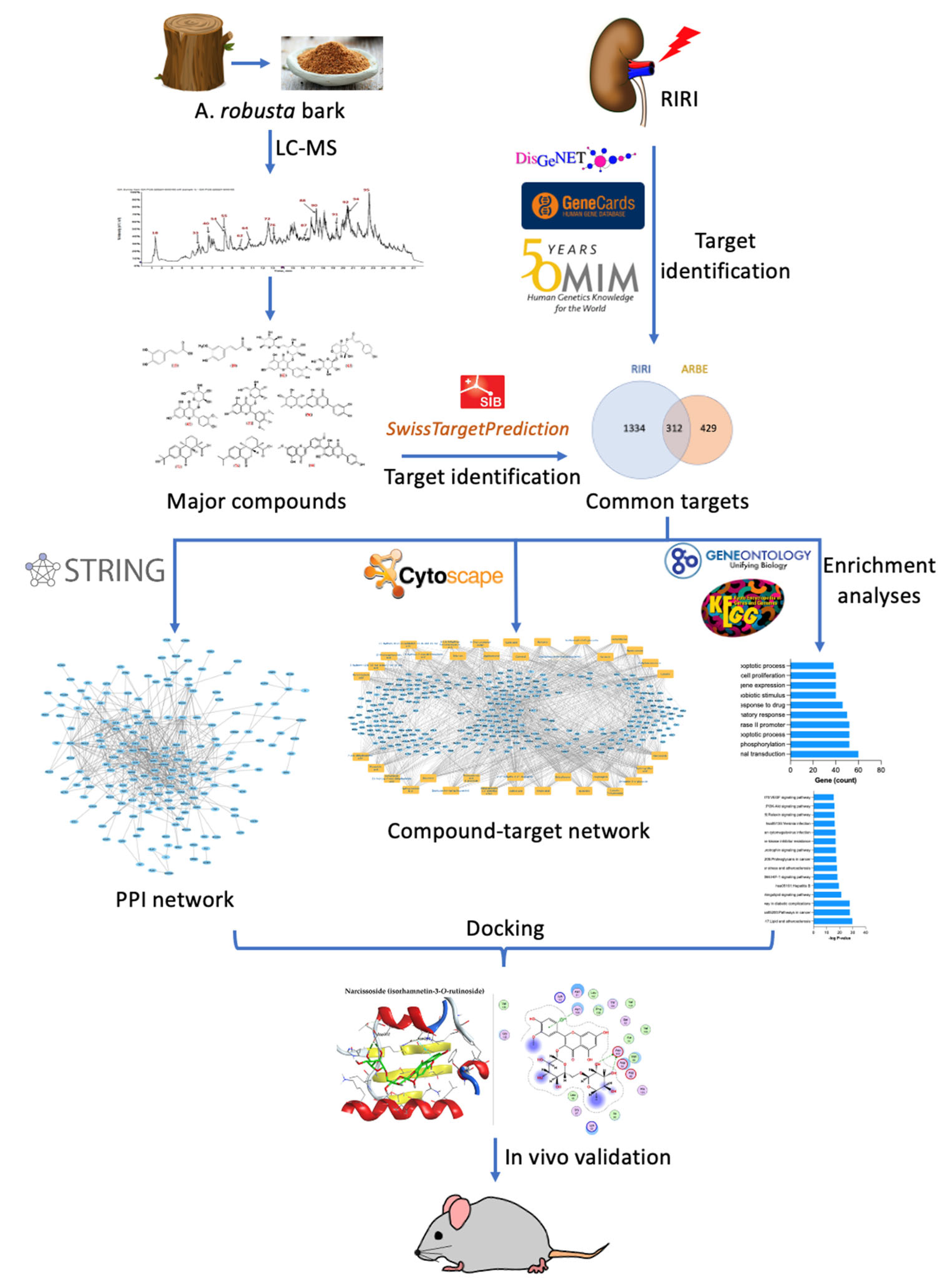
:1. Introduction
2. Results
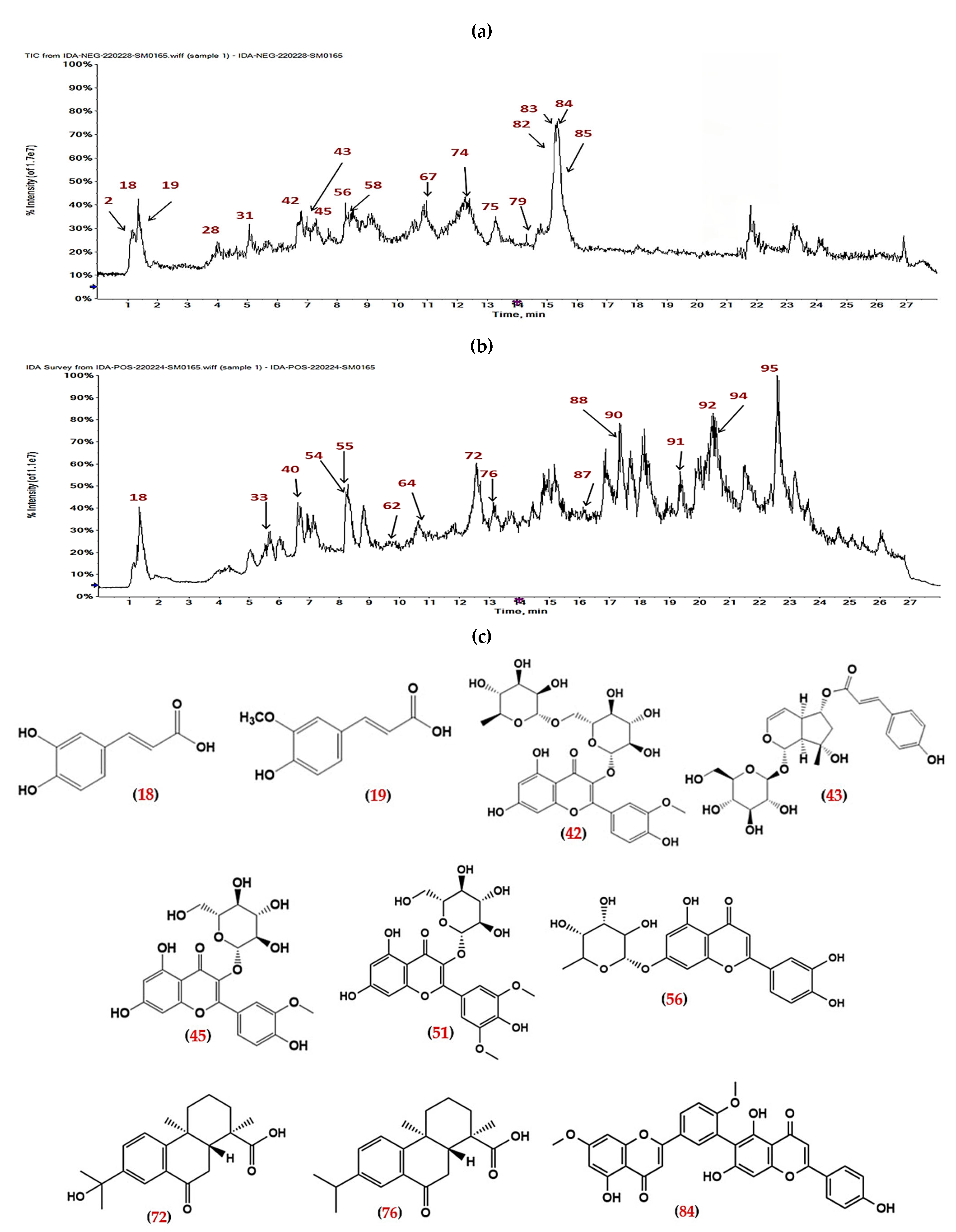
2.1. LC-ESI-MS/MS Profile
2.2. The Network Pahrmacology Analysis
2.2.1. Pharmacokinetics of ARBE Major Compounds
2.2.2. Molecular Targets of ARBE Major Compounds
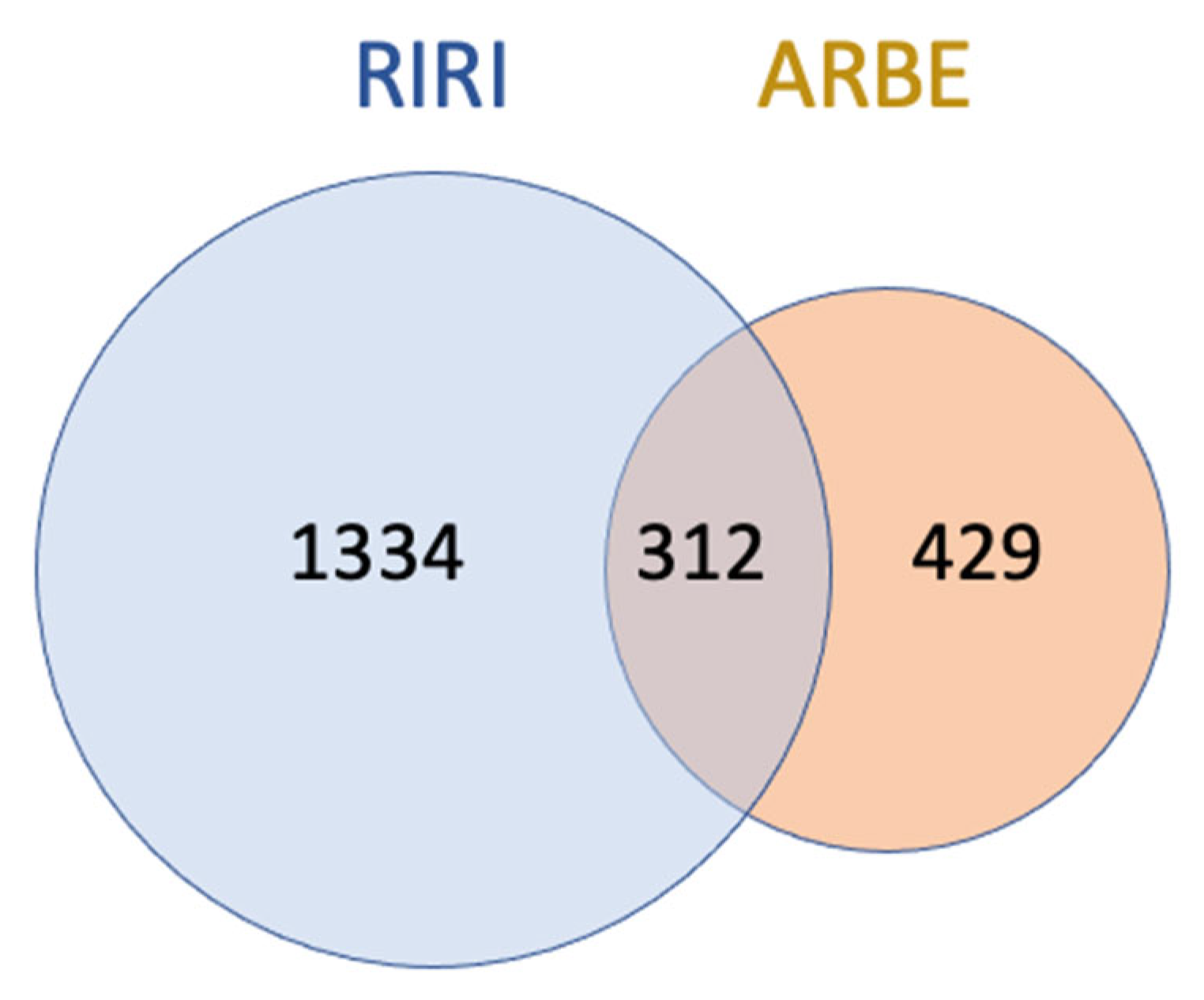
2.2.3. Molecular Targets of ARBE Associated with RIRI
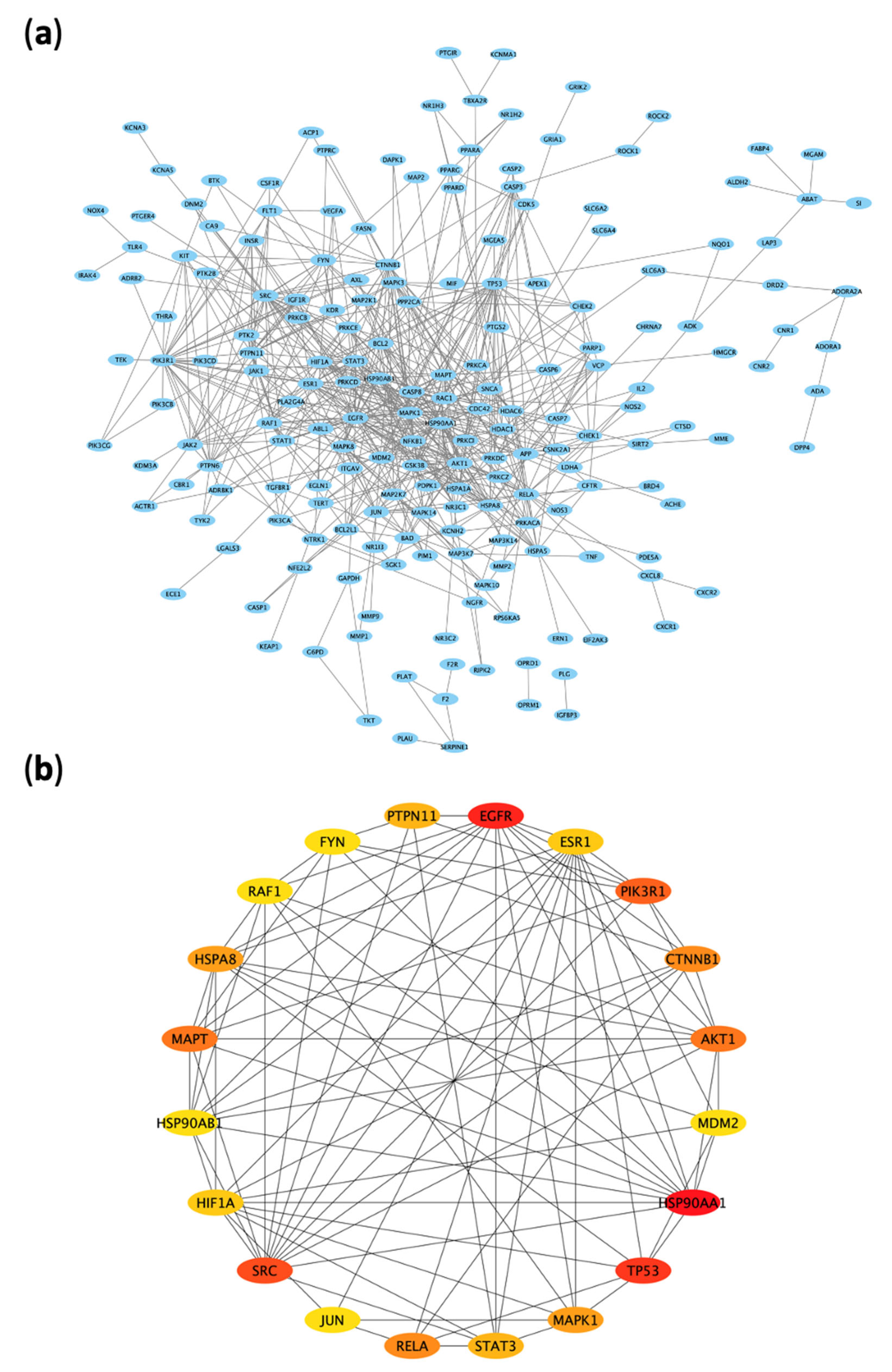
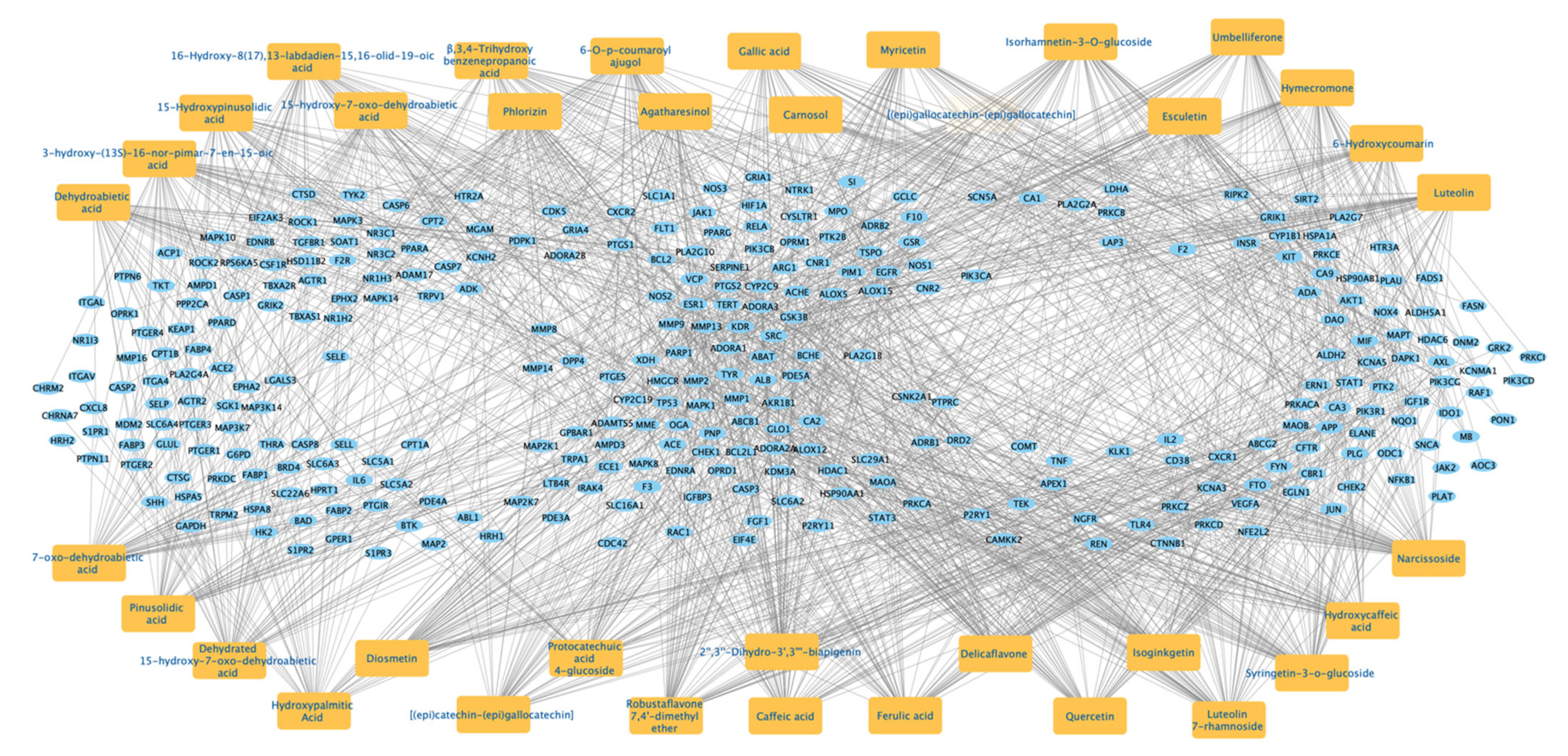
2.2.4. Protein–Protein Interaction (PPI) Network of the 312 Disease-Compounds Targets
2.2.5. Top ARBE Compounds Associated with RIRI Targets
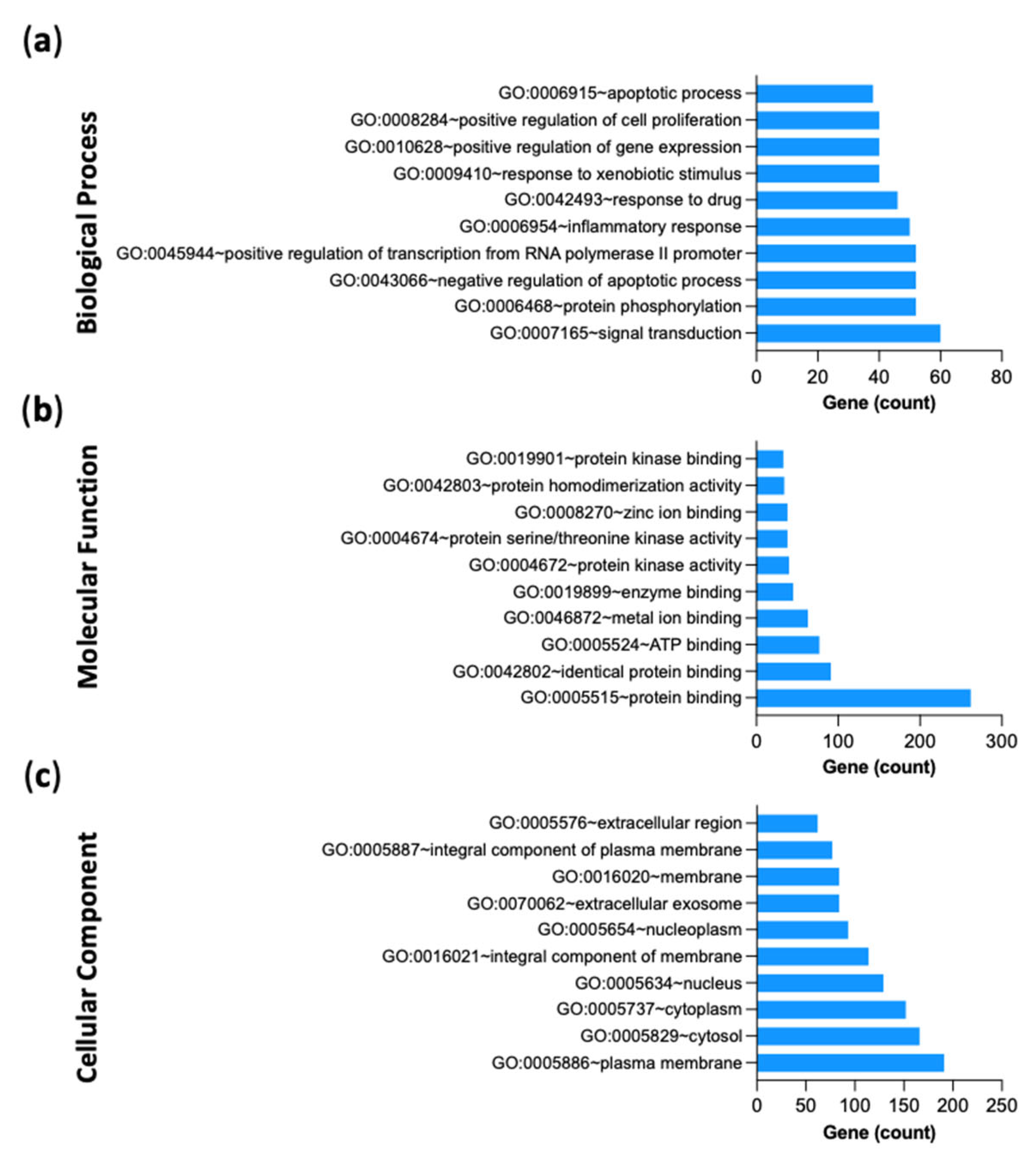
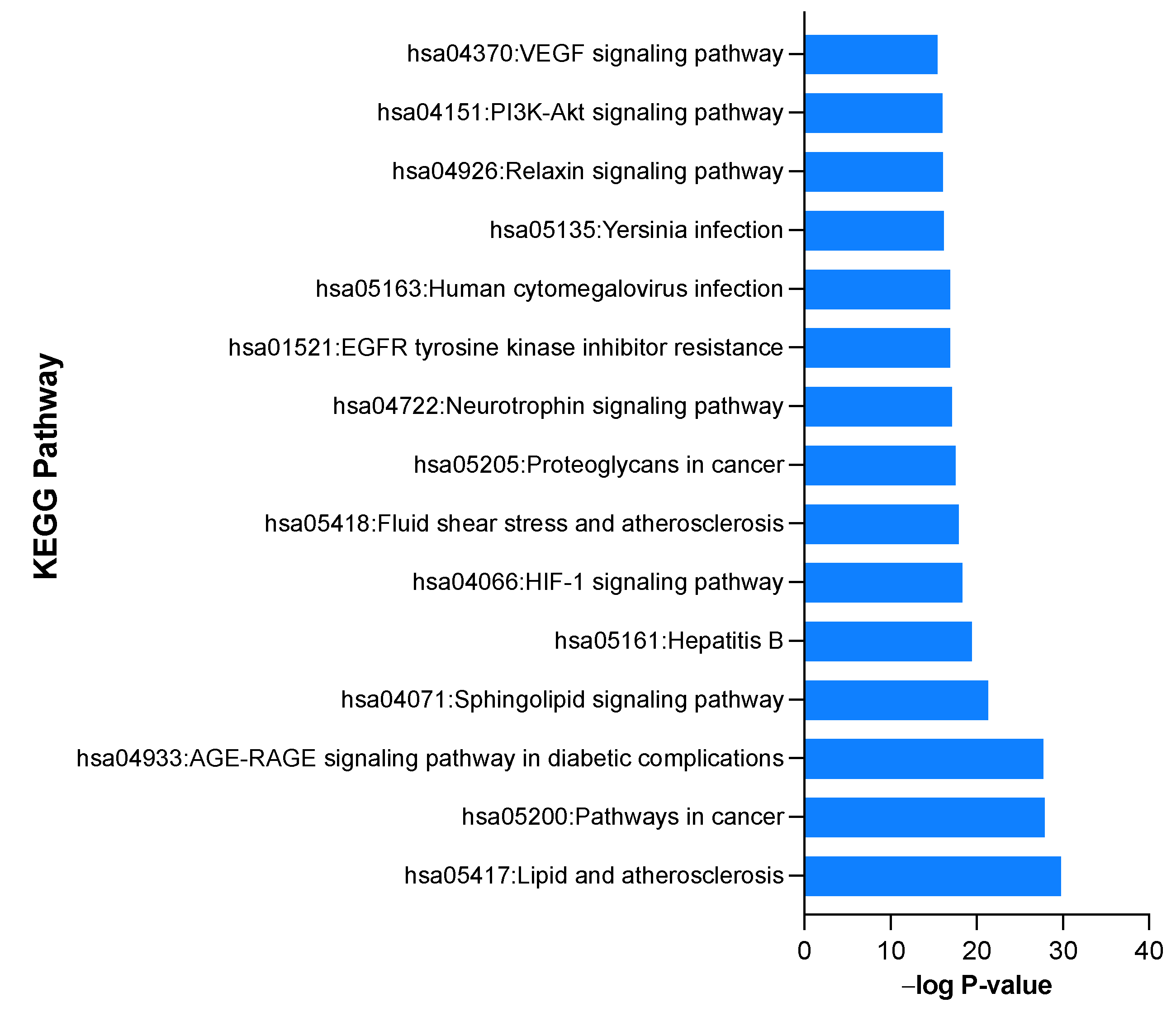
2.2.6. Gene-Ontology (GO) and KEGG Pathway Enrichment Analysis of 312 Common Targets
2.3. Molecular Docking Study
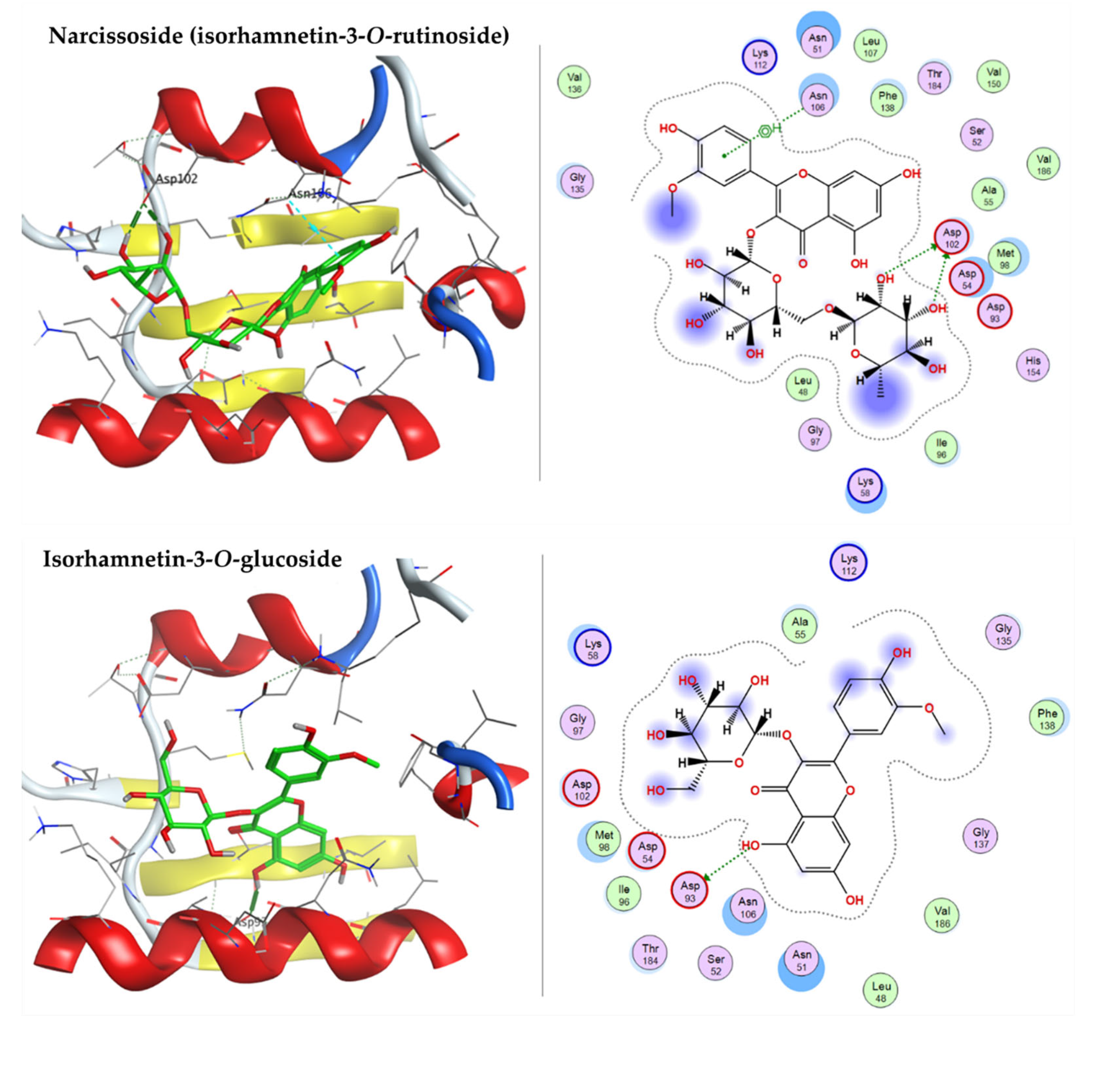
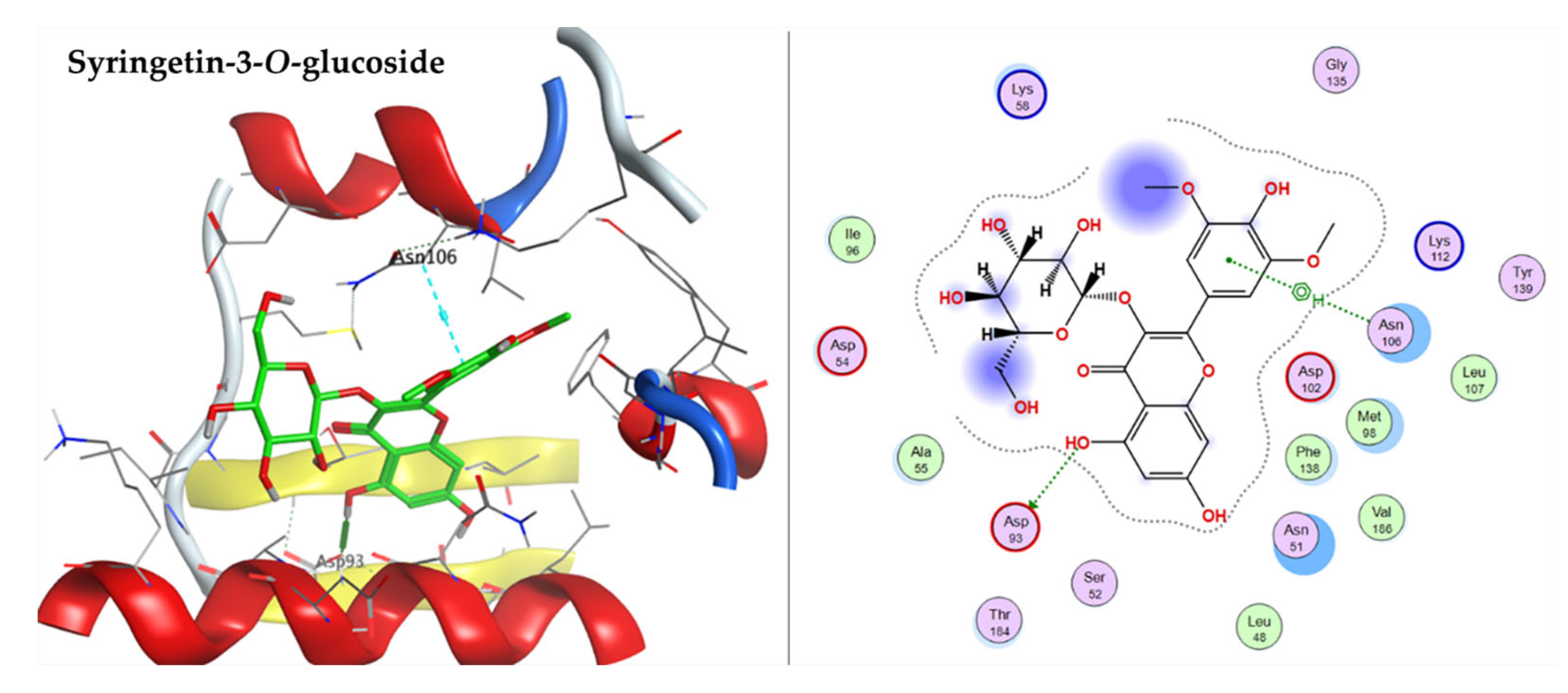
2.3.1. Docking with Heat shock Protein HSP 90-Alpha
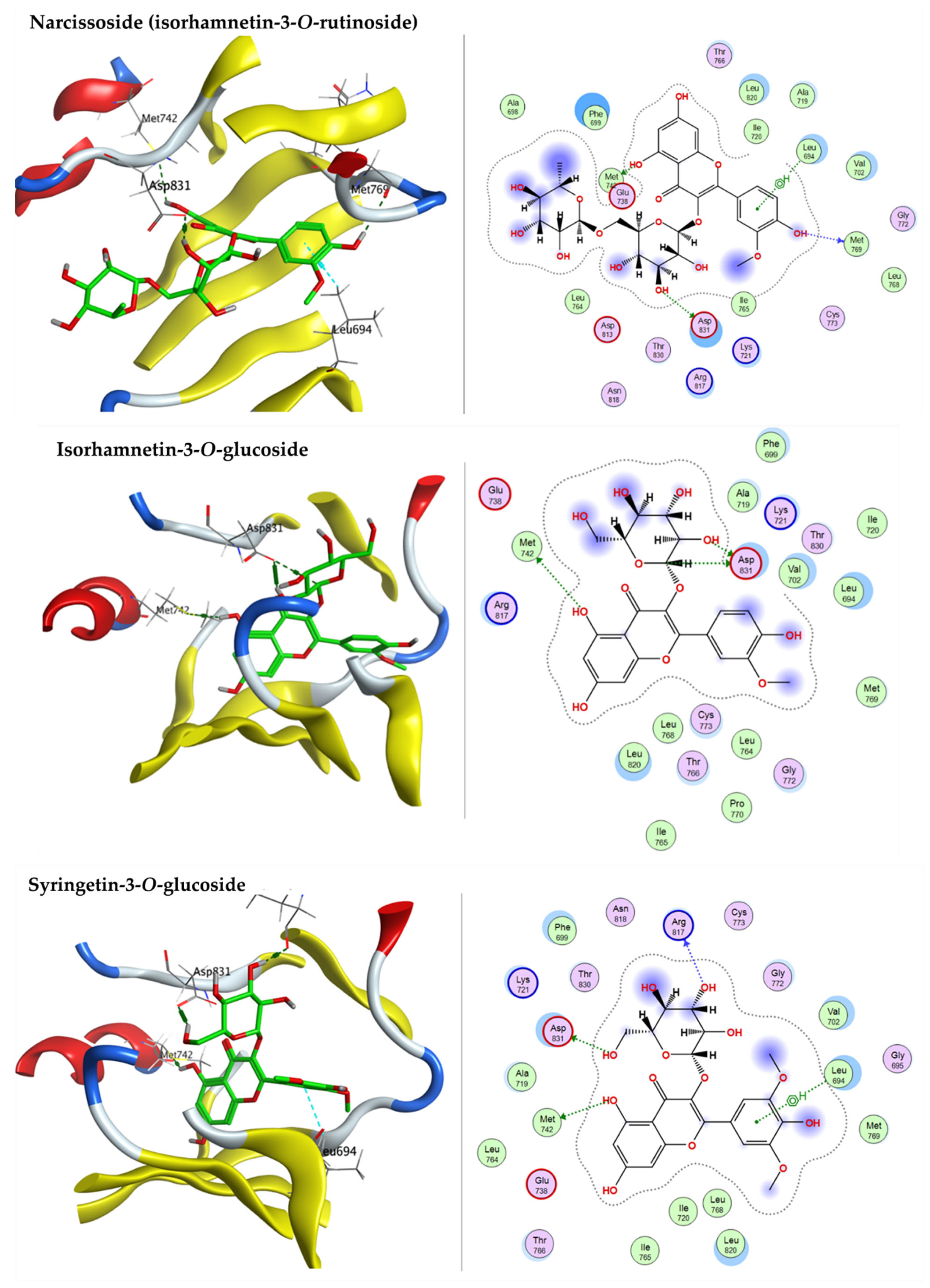
2.3.2. Docking with Epidermal Growth Factor Receptor (EGFR)
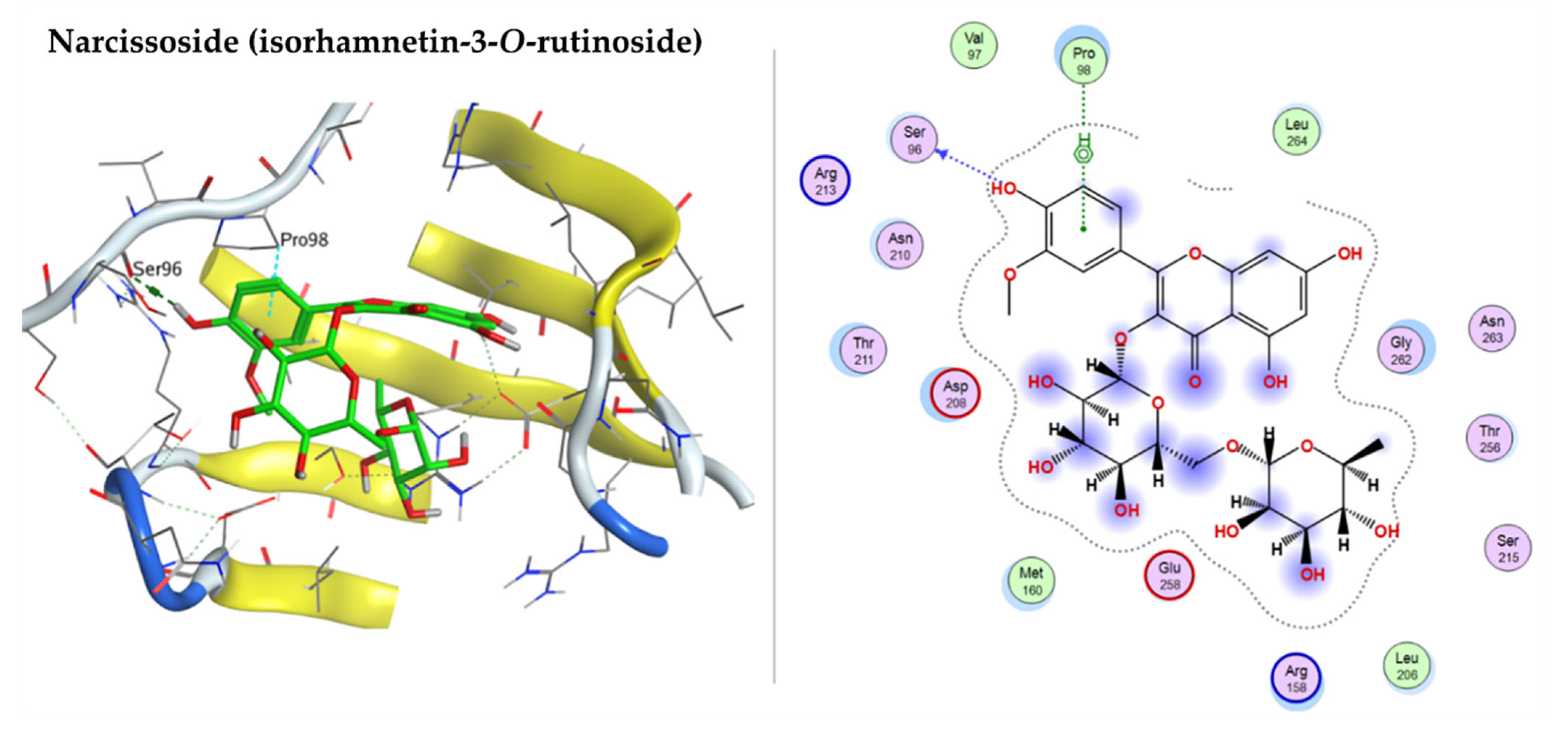
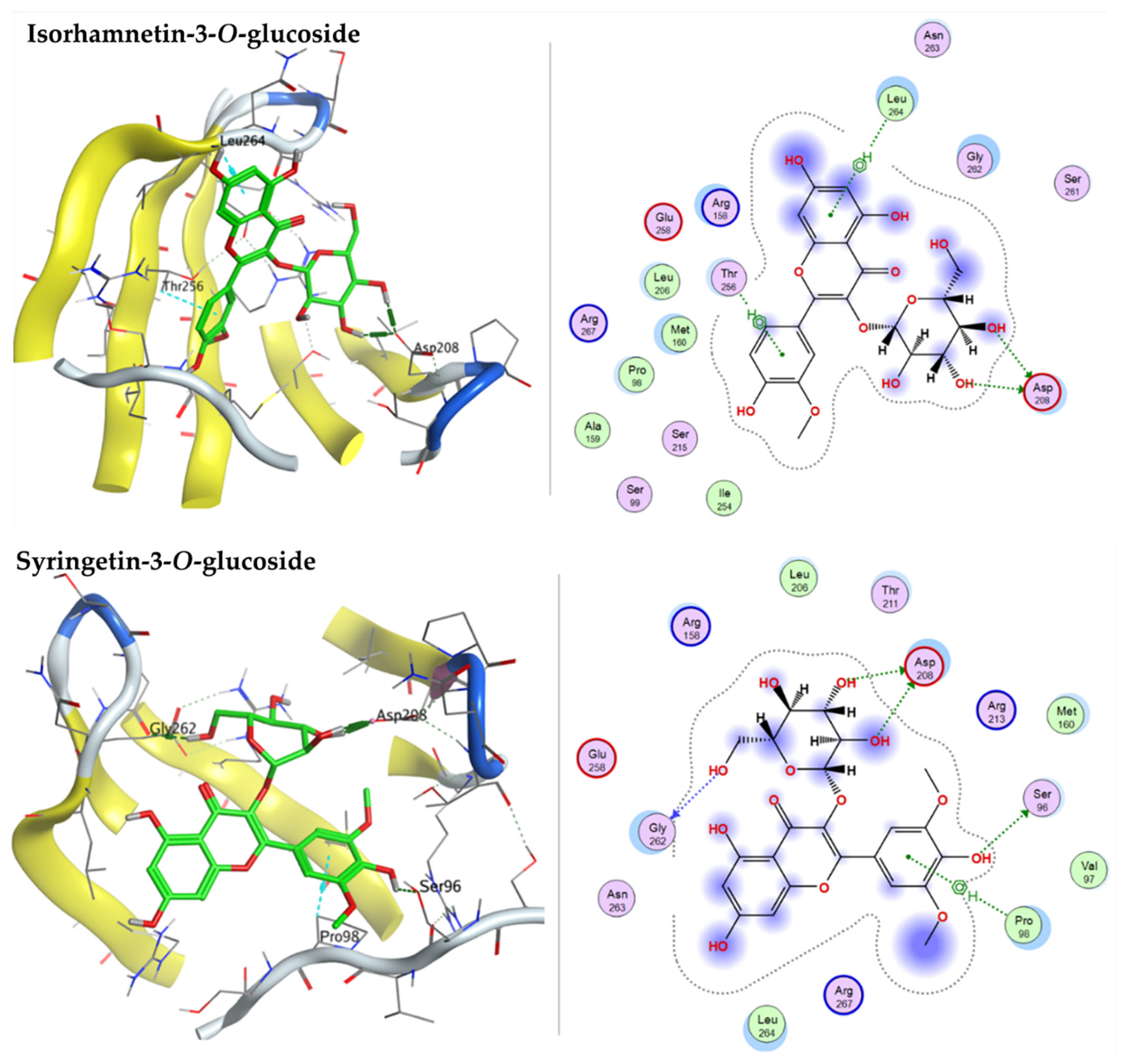
2.3.3. Docking with Cellular Tumor Antigen P53
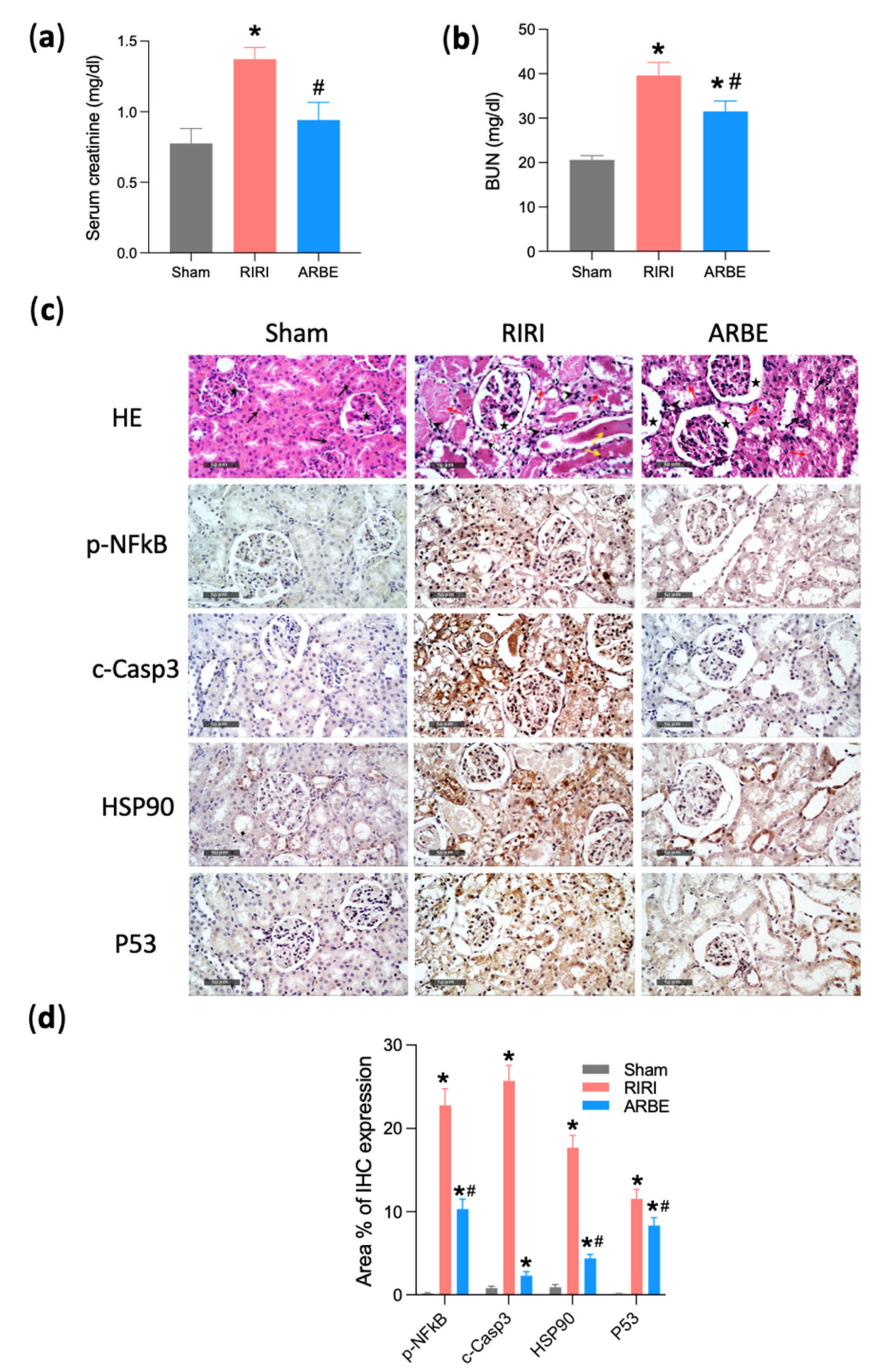
2.4. In Vivo Validation
3. Discussion
4. Materials and Methods
4.1. Plant Material and Extraction
4.2. LC-ESI-MS/MS Analysis
4.3. Network Pharmacology
4.3.1. Acquisition of Pharmacokinetics and Associated Targets of ARBE Major Compounds
4.3.2. Identification of RIRI-Associated Targets
4.3.3. Construction of PPI and Compound-Target Networks
4.3.4. The GO Analysis and KEGG Pathway Enrichment
4.4. Molecular Modelling
4.4.1. Molecular Docking Study
4.4.2. Protein Preparation
4.4.3. Ligand Preparation
4.5. In Vivo Experiments
4.5.1. Animals, Ethical Statement, and Experimental Design
4.5.2. Induction of Renal Ischemia-Reperfusion (RIRI)
4.5.3. Kidney Function Assessment
4.5.4. Histopathology
4.5.5. Immunohistochemistry
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- See, E.J.; Polkinghorne, K.R.; Toussaint, N.D.; Bailey, M.; Johnson, D.W.; Bellomo, R. Epidemiology and Outcomes of Acute Kidney Diseases: A Comparative Analysis. Am. J. Nephrol. 2021, 52, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Sharfuddin, A.A.; Molitoris, B.A. Pathophysiology of ischemic acute kidney injury. Nat. Rev. Nephrol. 2011, 7, 189–200. [Google Scholar] [CrossRef]
- Kellum, J.A.; Romagnani, P.; Ashuntantang, G.; Ronco, C.; Zarbock, A.; Anders, H.J. Acute kidney injury. Nat. Rev. Dis. Prim. 2021, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Bohlmann, J.; Gershenzon, J.; Aubourg, S. Biochemical, molecular genetic and evolutionary aspects of defense-related terpenoid metabolism in conifers. In Evolution of Metabolic Pathways; Elsevier: Amsterdam, The Netherlands, 2000; pp. 109–150. [Google Scholar]
- Frezza, C.; Venditti, A.; De Vita, D.; Toniolo, C.; Franceschin, M.; Ventrone, A.; Tomassini, L.; Foddai, S.; Guiso, M.; Nicoletti, M. Phytochemistry, chemotaxonomy, and biological activities of the Araucariaceae family—A review. Plants 2020, 9, 888. [Google Scholar] [CrossRef]
- Boland, D.J.; Brooker, M.I.H.; Chippendale, G.; Hall, N.; Hyland, B.; Johnston, R.; Kleinig, D.; McDonald, M.; Turner, J. Forest Trees of Australia; CSIRO publishing: Clayton, Australia, 2006. [Google Scholar]
- Verma, R.S.; Padalia, R.C.; Goswami, P.; Verma, S.K.; Chauhan, A.; Darokar, M.P. Chemical composition and antibacterial activity of the essential oil of Kauri Pine [Agathis robusta (C. Moore ex F. Muell.) FM Bailey] from India. J. Wood Chem. Technol. 2016, 36, 270–277. [Google Scholar] [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Wu, M.Z.; Fookes, C.J.; Forster, P.I. The steam volatile oil of Wollemia nobilis and its comparison with other members of the Araucariaceae (Agathis and Araucaria). Biochem. Syst. Ecol. 2000, 28, 563–578. [Google Scholar] [CrossRef]
- Dragota, S.; Riederer, M. Comparative study on epicuticular leaf waxes of Araucaria Araucana, Agathis robusta and Wollemia nobilis (Araucariaceae). Aust. J. Bot. 2008, 56, 644–650. [Google Scholar] [CrossRef]
- Carman, R.; Dennis, N. The diterpene acids of Agathis robusta oleoresin. Aust. J. Chem. 1964, 17, 390–392. [Google Scholar] [CrossRef] [Green Version]
- Carman, R.; Craig, W.; Shaw, I. Diterpenoids. XXXI. Three new resin acids. Aust. J. Chem. 1973, 26, 209–214. [Google Scholar] [CrossRef]
- Mohamed, M.E.; Tawfeek, N.; Elbaramawi, S.S.; Fikry, E. Agathis robusta Bark Essential Oil Effectiveness against COVID-19: Chemical Composition, In Silico and In Vitro Approaches. Plants 2022, 11, 663. [Google Scholar] [CrossRef]
- Venditti, A.; Frezza, C.; Campanelli, C.; Foddai, S.; Bianco, A.; Serafini, M. Phytochemical analysis of the ethanolic extract of Agathis robusta (C. Moore ex F. Muell.) FM Bailey. Nat. Prod. Res. 2017, 31, 1604–1611. [Google Scholar] [PubMed]
- Bisht, B.; Nainwal, P.; Saini, P. Evaluation of in vitro anti-inflammatory activity of Agathis robusta. J. Pharma. Res. 2012, 2, 1304–1306. [Google Scholar]
- Karar, M.G.E.; Pletzer, D.; Jaiswal, R.; Weingart, H.; Kuhnert, N. Identification, characterization, isolation and activity against Escherichia coli of quince (Cydonia oblonga) fruit polyphenols. Food Res. Int. 2014, 65, 121–129. [Google Scholar] [CrossRef]
- López-Fernández, O.; Domínguez, R.; Pateiro, M.; Munekata, P.E.; Rocchetti, G.; Lorenzo, J.M. Determination of polyphenols using liquid chromatography–tandem mass spectrometry technique (LC–MS/MS): A review. Antioxidants 2020, 9, 479. [Google Scholar] [CrossRef]
- Al Kadhi, O.; Melchini, A.; Mithen, R.; Saha, S. Development of a LC-MS/MS method for the simultaneous detection of tricarboxylic acid cycle intermediates in a range of biological matrices. J. Anal. Methods Chem. 2017, 2017, 5391832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.; Price, W.E.; Ashton, J.; Tapsell, L.C.; Johnson, S. Identification and characterization of phenolic compounds in hydromethanolic extracts of sorghum wholegrains by LC-ESI-MSn. Food Chem. 2016, 211, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Pallerla, P.; Bhumireddy, S.R.; Lingampally, S.S.; Ragi, N.C.; Sripadi, P. ESI-MS/MS analysis of protonated N-methyl amino acids and their immonium ions. J. Mass Spectrom. 2019, 54, 761–771. [Google Scholar] [CrossRef]
- Kiprop, A.; Pourtier, E.; Kimutai, S. GC-MS and ESI-MS detection of catechol. Int. J. Educ. Res. 2013, 1, 1–12. [Google Scholar]
- Holm, P.I.; Ueland, P.M.; Kvalheim, G.; Lien, E.A. Determination of choline, betaine, and dimethylglycine in plasma by a high-throughput method based on normal-phase chromatography–tandem mass spectrometry. Clin. Chem. 2003, 49, 286–294. [Google Scholar] [CrossRef]
- Cocuron, J.-C.; Alonso, A.P. Liquid chromatography tandem mass spectrometry for measuring 13 C-labeling in intermediates of the glycolysis and pentose phosphate pathway. In Plant Metabolic Flux Analysis; Springer: Berlin/Heidelberg, Germany, 2014; pp. 131–142. [Google Scholar]
- Modick, H.; Schütze, A.; Pälmke, C.; Weiss, T.; Brüning, T.; Koch, H.M. Rapid determination of N-acetyl-4-aminophenol (paracetamol) in urine by tandem mass spectrometry coupled with on-line clean-up by two dimensional turbulent flow/reversed phase liquid chromatography. J. Chromatogr. B. 2013, 925, 33–39. [Google Scholar] [CrossRef]
- Chen, F.; Zhang, F.; Yang, N.; Liu, X. Simultaneous determination of 10 nucleosides and nucleobases in Antrodia camphorata using QTRAP LC–MS/MS. J. Chromatogr. Sci. 2014, 52, 852–861. [Google Scholar] [CrossRef] [PubMed]
- Avula, B.; Wang, Y.-H.; Smillie, T.J.; Khan, I.A. Determination of shikimic acid in fruits of Illicium species and various other plant samples by LC–UV and LC–ESI–MS. Chromatographia 2009, 69, 307–314. [Google Scholar] [CrossRef]
- Zhang, Q.; Ford, L.A.; Evans, A.M.; Toal, D.R. Structure elucidation of metabolite x17299 by interpretation of mass spectrometric data. Metabolomics 2017, 13, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.-Z.; Harnly, J.M. Identification of the phenolic components of chrysanthemum flower (Chrysanthemum morifolium Ramat). Food Chem. 2010, 120, 319–326. [Google Scholar] [CrossRef]
- Chen, H.; Zhou, Y.; Shao, Y.; Chen, F. Free phenolic acids in Shanxi aged vinegar: Changes during aging and synergistic antioxidant activities. Int. J. Food Prop. 2016, 19, 1183–1193. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Lin, Z.; Jiang, H.; Tong, L.; Wang, H.; Chen, S. Rapid identification and assignation of the active ingredients in fufang banbianlian injection using HPLC-DAD-ESI-IT-TOF-MS. J. Chromatogr. Sci. 2016, 54, 1225–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, K.V.; Bonham, C.C.; Miles, D.; Rothwell, A.P.; Peel, G.; Wood, B.C.; Rhodes, D. Characterization of betaines using electrospray MS/MS. Phytochemistry 2002, 59, 759–765. [Google Scholar] [CrossRef]
- Jaiswal, R.; Jayasinghe, L.; Kuhnert, N. Identification and characterization of proanthocyanidins of 16 members of the Rhododendron genus (Ericaceae) by tandem LC–MS. J. Mass Spectrom. 2012, 47, 502–515. [Google Scholar] [CrossRef]
- Tawfeek, N.; Sobeh, M.; Hamdan, D.I.; Farrag, N.; Roxo, M.; El-Shazly, A.M.; Wink, M. Phenolic compounds from Populus alba L. and Salix subserrata Willd. (Salicaceae) counteract oxidative stress in Caenorhabditis elegans. Molecules 2019, 24, 1999. [Google Scholar] [CrossRef] [Green Version]
- Pozo, O.J.; Gómez, C.; Marcos, J.; Segura, J.; Ventura, R. Detection and characterization of urinary metabolites of boldione by LC-MS/MS. Part II: Conjugates with cysteine and N-acetylcysteine. Drug Test. Anal. 2012, 4, 786–797. [Google Scholar] [CrossRef]
- Hong, Y.; Wang, Z.; Barrow, C.J.; Dunshea, F.R.; Suleria, H.A. High-throughput screening and characterization of phenolic compounds in stone fruits waste by lc-esi-qtof-ms/ms and their potential antioxidant activities. Antioxidants 2021, 10, 234. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.A.; Khalil, W.A.; Hassan, M.A.; Yousif, A.I.; Sabry, O.M.; Wink, M.; Sobeh, M. Antioxidant and Antiapoptotic Effects of a Turraea fischeri Leaf Extract on Cryopreserved Goat Sperm. Animals 2021, 11, 2840. [Google Scholar] [CrossRef]
- Zhao, H.-Q.; Wang, X.; Li, H.-M.; Yang, B.; Yang, H.-J.; Huang, L. Characterization of nucleosides and nucleobases in natural cordyceps by HILIC–ESI/TOF/MS and HILIC–ESI/MS. Molecules 2013, 18, 9755–9769. [Google Scholar] [CrossRef] [Green Version]
- Lv, L.; Shao, X.; Wang, L.; Huang, D.; Ho, C.-T.; Sang, S. Stilbene glucoside from Polygonum multiflorum Thunb.: A novel natural inhibitor of advanced glycation end product formation by trapping of methylglyoxal. J. Agric. Food Chem. 2010, 58, 2239–2245. [Google Scholar] [CrossRef] [PubMed]
- Yun, E.S.; Park, S.K.; Kim, B.S.; Chae, Y.Z.; Cho, S.M.; Yi, H.; Cho, H.J.; Shin, H.C. Determination of the esculetin contents of medicinal plants by liquid chromatography–tandem mass spectrometry. Biomed. Chromatogr. 2012, 26, 1247–1251. [Google Scholar] [CrossRef] [PubMed]
- Routaboul, J.-M.; Kerhoas, L.; Debeaujon, I.; Pourcel, L.; Caboche, M.; Einhorn, J.; Lepiniec, L. Flavonoid diversity and biosynthesis in seed of Arabidopsis thaliana. Planta 2006, 224, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Yang, W.-Z.; Liu, K.-D.; Qiao, X.; Li, B.-J.; Cheng, J.; Feng, J.; Guo, D.-A.; Zhao, Y.-Y. Characterization of flavonoids in Millettia nitida var. hirsutissima by HPLC/DAD/ESI-MSn. J. Pharm. Anal. 2012, 2, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Yu, H.; Wu, H.; Pan, Y.; Wang, K.; Jin, Y.; Zhang, C. Characterization and quantification by LC-MS/MS of the chemical components of the heating products of the flavonoids extract in pollen typhae for transformation rule exploration. Molecules 2015, 20, 18352–18366. [Google Scholar] [CrossRef] [Green Version]
- Park, D.; Shin, H.; Byun, Y.; Lee, K.Y. Bioassay-coupled LC-QTOF MS/MS to Characterize Constituents Inhibiting Nitric Oxide Production of Thuja orientalis. Nat. Prod. Sci. 2021, 27, 293–299. [Google Scholar]
- Li, X.-N.; Sun, J.; Shi, H.; Yu, L.L.; Ridge, C.D.; Mazzola, E.P.; Okunji, C.; Iwu, M.M.; Michel, T.K.; Chen, P. Profiling hydroxycinnamic acid glycosides, iridoid glycosides, and phenylethanoid glycosides in baobab fruit pulp (Adansonia digitata). Food Res. Int. 2017, 99, 755–761. [Google Scholar] [CrossRef]
- Cambie, R.C.; Coddington, J.M.; Stone, M.J.; Tanaka, N.; Li, Y.H.; Arigayo, S. Diterpenoids of the wood of Agathis vitiensis. Phytochemistry 1989, 28, 1675–1679. [Google Scholar] [CrossRef]
- Bell, L.; Oruna-Concha, M.J.; Wagstaff, C. Identification and quantification of glucosinolate and flavonol compounds in rocket salad (Eruca sativa, Eruca vesicaria and Diplotaxis tenuifolia) by LC–MS: Highlighting the potential for improving nutritional value of rocket crops. Food Chem. 2015, 172, 852–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Yousef, H.M.; Abdelaziz, S.; Hassan, W.H.; El-Sayed, M.A. Phytochemical and biological characterization of Tephrosia nubica boiss. Growing in Saudi Arabia. Arab. J. Chem. 2020, 13, 9216–9230. [Google Scholar] [CrossRef]
- Lijia, X.; Guo, J.; Chen, Q.; Baoping, J.; Zhang, W. Quantitation of phlorizin and phloretin using an ultra high performance liquid chromatography–electrospray ionization tandem mass spectrometric method. J. Chromatogr. B. 2014, 960, 67–72. [Google Scholar] [CrossRef]
- Yang, Z.; Nakabayashi, R.; Okazaki, Y.; Mori, T.; Takamatsu, S.; Kitanaka, S.; Kikuchi, J.; Saito, K. Toward better annotation in plant metabolomics: Isolation and structure elucidation of 36 specialized metabolites from Oryza sativa (rice) by using MS/MS and NMR analyses. Metabolomics 2014, 10, 543–555. [Google Scholar] [CrossRef] [Green Version]
- Xian, X.; Sun, B.; Ye, X.; Zhang, G.; Hou, P.; Gao, H. Identification and analysis of alkaloids in cortex Phellodendron amurense by high-performance liquid chromatography with electrospray ionization mass spectrometry coupled with photodiode array detection. J. Sep. Sci. 2014, 37, 1533–1545. [Google Scholar] [CrossRef]
- Latiff, N.A.; Suan, C.L.; Sarmidi, M.R.; Ware, I.; Rashid, S.N.A.A.; Yahayu, M. Liquid chromatography tandem mass spectrometry for the detection and validation of quercetin-3-O-rutinoside and myricetin from fractionated Labisia pumila var. Alata. Malays. J. Anal. Sci. 2018, 22, 817–827. [Google Scholar]
- Wang, B.; Liu, X.; Zhou, A.; Meng, M.; Li, Q. Simultaneous analysis of coumarin derivatives in extracts of Radix Angelicae pubescentis (Duhuo) by HPLC-DAD-ESI-MS n technique. Anal. Methods 2014, 6, 7996–8002. [Google Scholar] [CrossRef]
- Sulaiman, C.T.; Ramesh, P.R.; Madhu, K.M.; Anandan, E.M.; Balachandran, I. Chemical profiling of a polyherbal formulation by tandem mass spectroscopic analysis with multiple ionization techniques. Future J. Pharm. Sci. 2020, 6, 1–8. [Google Scholar]
- Colombo, R.; Yariwake, J.H.; Queiroz, E.F.; Ndjoko, K.; Hostettmann, K. LC-MS/MS analysis of sugarcane extracts and differentiation of monosaccharides moieties of flavone C-glycosides. J. Liq. Chromatogr. Relat. Technol. 2013, 36, 239–248. [Google Scholar] [CrossRef]
- Formigonia, M.; Milanib, P.G.; Zorzenona, M.R. Analysis of commercial stevia extracts composition by HPLC and UHPLC-MS-MS-QToF. Chem. Eng. 2019, 75, 355–360. [Google Scholar]
- Saber, F.R.; Mohsen, E.; El-Hawary, S.; Eltanany, B.M.; Elimam, H.; Sobeh, M.; Elmotayam, A.K. Chemometric-enhanced metabolic profiling of five Pinus species using HPLC-MS/MS spectrometry: Correlation to in vitro anti-aging, anti-Alzheimer and antidiabetic activities. J. Chromatogr. B. 2021, 1177, 122759. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, K.J. Analysis of Diterpenoid Resins And Polymers in Paint Media and Varnishes with an Atlas of Mass Spectra; FOM Institute AMOLF: Amsterdam, The Netherlands, (MOLART Report 10); 2003. [Google Scholar]
- Lobo, G.P.; Amengual, J.; Li, H.N.M.; Golczak, M.; Bonet, M.L.; Palczewski, K.; Von Lintig, J. β, β-carotene decreases peroxisome proliferator receptor γ activity and reduces lipid storage capacity of adipocytes in a β, β-carotene oxygenase 1-dependent manner. J. Biol. Chem. 2010, 285, 27891–27899. [Google Scholar] [CrossRef] [Green Version]
- Tsimogiannis, D.; Samiotaki, M.; Panayotou, G.; Oreopoulou, V. Characterization of flavonoid subgroups and hydroxy substitution by HPLC-MS/MS. Molecules 2007, 12, 593–606. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.; Chen, B.; Zhang, Y.; Ou, H.; Li, Y.; Li, S.; Shi, P.; Lin, X. Analysis of the total biflavonoids extract from Selaginella doederleinii by HPLC-QTOF-MS and its in vitro and in vivo anticancer effects. Molecules 2017, 22, 325. [Google Scholar] [CrossRef] [Green Version]
- Falcão, S.I.; Vilas-Boas, M.; Estevinho, L.M.; Barros, C.; Domingues, M.R.; Cardoso, S.M. Phenolic characterization of Northeast Portuguese propolis: Usual and unusual compounds. Anal. Bioanal. Chem. 2010, 396, 887–897. [Google Scholar] [CrossRef]
- Chen, G.; Li, X.; Saleri, F.; Guo, M. Analysis of flavonoids in Rhamnus davurica and its antiproliferative activities. Molecules 2016, 21, 1275. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Liu, L.; Wang, L.; Hu, Y.; Zhang, W.; Liu, R. Structural characterization and identification of major constituents in Jitai tablets by high-performance liquid chromatography/diode-array detection coupled with electrospray ionization tandem mass spectrometry. Molecules 2012, 17, 10470–10493. [Google Scholar] [CrossRef] [PubMed]
- Rauf, A.; Imran, M.; Abu-Izneid, T.; Patel, S.; Pan, X.; Naz, S.; Silva, A.S.; Saeed, F.; Suleria, H.A.R. Proanthocyanidins: A comprehensive review. Biomed. Pharmacother. 2019, 116, 108999. [Google Scholar] [CrossRef]
- Murray, C.W.; Carr, M.G.; Callaghan, O.; Chessari, G.; Congreve, M.; Cowan, S.; Coyle, J.E.; Downham, R.; Figueroa, E.; Frederickson, M.; et al. Fragment-Based Drug Discovery Applied to Hsp90. Discovery of Two Lead Series with High Ligand Efficiency. J. Med. Chem. 2010, 53, 5942–5955. [Google Scholar] [CrossRef]
- Stamos, J.; Sliwkowski, M.X.; Eigenbrot, C. Structure of the epidermal growth factor receptor kinase domain alone and in complex with a 4-anilinoquinazoline inhibitor. J. Biol. Chem. 2002, 277, 46265–46272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petty, T.J.; Emamzadah, S.; Costantino, L.; Petkova, I.; Stavridi, E.S.; Saven, J.G.; Vauthey, E.; Halazonetis, T.D. An induced fit mechanism regulates p53 DNA binding kinetics to confer sequence specificity. EMBO J. 2011, 30, 2167–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O'Neill, S.; Humphries, D.; Tse, G.; Marson, L.P.; Dhaliwal, K.; Hughes, J.; Ross, J.A.; Wigmore, S.J.; Harrison, E.M. Heat shock protein 90 inhibition abrogates TLR4-mediated NF-κB activity and reduces renal ischemia-reperfusion injury. Sci. Rep. 2015, 5, 12958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, Y.; Kim, J.; Westphal, S.N.; Long, K.E.; Padanilam, B.J. Targeted deletion of p53 in the proximal tubule prevents ischemic renal injury. J. Am. Soc. Nephrol. 2014, 25, 2707–2716. [Google Scholar] [CrossRef] [Green Version]
- Adnan, M.; Jeon, B.B.; Chowdhury, M.H.U.; Oh, K.K.; Das, T.; Chy, M.N.U.; Cho, D.H. Network Pharmacology Study to Reveal the Potentiality of a Methanol Extract of Caesalpinia sappan L. Wood against Type-2 Diabetes Mellitus. Life 2022, 12, 277. [Google Scholar] [CrossRef]
- Zhang, R.; Zhu, X.; Bai, H.; Ning, K. Network Pharmacology Databases for Traditional Chinese Medicine: Review and Assessment. Front. Pharmacol. 2019, 10, 123. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, S.H.; Zhong, K.; Jiang, T.; Zhang, M.; Kwan, H.Y.; Su, T. Network Pharmacology-Based Strategy for the Investigation of the Anti-Obesity Effects of an Ethanolic Extract of Zanthoxylum bungeanum Maxim. Front. Pharmacol. 2020, 11, 572387. [Google Scholar] [CrossRef]
- Elbatreek, M.H.; Sadegh, S.; Anastasi, E.; Guney, E.; Nogales, C.; Kacprowski, T.; Hassan, A.A.; Teubner, A.; Huang, P.H.; Hsu, C.Y.; et al. NOX5-induced uncoupling of endothelial NO synthase is a causal mechanism and theragnostic target of an age-related hypertension endotype. PLoS Biol. 2020, 18, e3000885. [Google Scholar] [CrossRef]
- Dubey, A.; Prajapati, K.S.; Swamy, M.; Pachauri, V. Heat shock proteins: A therapeutic target worth to consider. Vet. World. 2015, 8, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Sabbah, D.A.; Hajjo, R.; Sweidan, K. Review on Epidermal Growth Factor Receptor (EGFR) Structure, Signaling Pathways, Interactions, and Recent Updates of EGFR Inhibitors. Curr. Top. Med. Chem. 2020, 20, 815–834. [Google Scholar] [CrossRef]
- Hafner, A.; Bulyk, M.L.; Jambhekar, A.; Lahav, G. The multiple mechanisms that regulate p53 activity and cell fate. Nat. Rev. Mol. Cell. Biol. 2019, 20, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.Y.; Zhang, H.X.; Mezei, M.; Cui, M. Molecular docking: A powerful approach for structure-based drug discovery. Curr. Comput. Aided Drug Des. 2011, 7, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Pratt, W.B.; Toft, D.O. Regulation of signaling protein function and trafficking by the hsp90/hsp70-based chaperone machinery. Exp. Biol. Med. 2003, 228, 111–133. [Google Scholar] [CrossRef] [PubMed]
- Pearl, L.H.; Prodromou, C. Structure and mechanism of the Hsp90 molecular chaperone machinery. Annu. Rev. Biochem. 2006, 75, 271–294. [Google Scholar] [CrossRef] [PubMed]
- Woodhead, A.J.; Angove, H.; Carr, M.G.; Chessari, G.; Congreve, M.; Coyle, J.E.; Cosme, J.; Graham, B.; Day, P.J.; Downham, R. Discovery of (2, 4-Dihydroxy-5-isopropylphenyl)-[5-(4-methylpiperazin-1-ylmethyl)-1, 3-dihydroisoindol-2-yl] methanone (AT13387), a Novel Inhibitor of the Molecular Chaperone Hsp90 by Fragment Based Drug Design. J. Med. Chem. 2010, 53, 5956–5969. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Chen, C.; Lei, X.; Zhao, J.; Liang, J. CASTp 3.0: Computed atlas of surface topography of proteins. Nucleic Acids Res. 2018, 46, W363–W367. [Google Scholar] [CrossRef]
- Liu, X.Q.; Liu, M.M.; Jiang, L.; Gao, L.; Zhang, Y.; Huang, Y.B.; Wang, X.; Zhu, W.; Zeng, H.X.; Meng, X.M.; et al. A novel small molecule Hsp90 inhibitor, C-316–1, attenuates acute kidney injury by suppressing RIPK1-mediated inflammation and necroptosis. Int. Immunopharmacol. 2022, 108, 108849. [Google Scholar] [CrossRef]
- Baisantry, A.; Berkenkamp, B.; Rong, S.; Bhayadia, R.; Sörensen-Zender, I.; Schmitt, R.; Melk, A. Time-dependent p53 inhibition determines senescence attenuation and long-term outcome after renal ischemia-reperfusion. Am. J. Physiol. Renal. Physiol. 2019, 316, F1124–F1132. [Google Scholar] [CrossRef]
- Molitoris, B.A.; Dagher, P.C.; Sandoval, R.M.; Campos, S.B.; Ashush, H.; Fridman, E.; Brafman, A.; Faerman, A.; Atkinson, S.J.; Thompson, J.D.; et al. siRNA targeted to p53 attenuates ischemic and cisplatin-induced acute kidney injury. J. Am. Soc. Nephrol. 2009, 20, 1754–1764. [Google Scholar] [CrossRef] [Green Version]
- Peddi, V.; Ratner, L.; Cooper, M.; Gaber, O.; Feng, S.; Tso, P.; Bowers, V.; Naraghi, R.; Budde, K.; Polinsky, M.; et al. Treatment with QPI-1002, a Short Interfering (SI) RNA for the Prophylaxis of Delayed Graft Function.: Abstract# 2967. Transplantation 2014, 98, 153. [Google Scholar]
- Mohammed, H.A.; Khan, R.A.; Abdel-Hafez, A.A.; Abdel-Aziz, M.; Ahmed, E.; Enany, S.; Mahgoub, S.; Al-Rugaie, O.; Alsharidah, M.; Aly, M.S. Phytochemical profiling, in vitro and in silico anti-microbial and anti-cancer activity evaluations and Staph GyraseB and h-TOP-IIβ receptor-docking studies of major constituents of Zygophyllum coccineum L. Aqueous-ethanolic extract and its subsequent fractions: An approach to validate traditional phytomedicinal knowledge. Molecules 2021, 26, 577. [Google Scholar] [PubMed]
- Daina, S. A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piñero, J.; Ramírez-Anguita, J.M.; Saüch-Pitarch, J.; Ronzano, F.; Centeno, E.; Sanz, F.; Furlong, L.I. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2020, 48, D845–D855. [Google Scholar] [CrossRef] [Green Version]
- Rebhan, M.; Chalifa-Caspi, V.; Prilusky, J.; Lancet, D. GeneCards: Integrating information about genes, proteins and diseases. Trends Genet. TIG 1997, 13, 163. [Google Scholar] [CrossRef]
- Safran, M.; Dalah, I.; Alexander, J.; Rosen, N.; Iny Stein, T.; Shmoish, M.; Nativ, N.; Bahir, I.; Doniger, T.; Krug, H. GeneCards Version 3: The human gene integrator. Database 2010, 2010, baq020. [Google Scholar] [CrossRef] [Green Version]
- Hamosh, A.; Scott, A.F.; Amberger, J.S.; Bocchini, C.A.; McKusick, V.A. Online Mendelian Inheritance in Man (OMIM), a knowledgebase of human genes and genetic disorders. Nucleic Acids Res. 2005, 33, D514–D517. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P. The STRING database in 2021: Customizable protein–protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef] [Green Version]
- Dennis, G.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for annotation, visualization, and integrated discovery. Genome Biol. 2003, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Molecular Operating Environment. Chemical Computing Group Inc.: Montreal, QC, Canada, 2019; (MOE 2019.0102).
- RCSB Protein Data Bank (PDB). Available online: https://www.rcsb.org/ (accessed on 17 July 2022).
- Xie, G.-L.; Zhu, L.; Zhang, Y.-m.; Zhang, Q.-N.; Yu, Q. Change in iron metabolism in rats after renal ischemia/reperfusion injury. PLoS ONE 2017, 12, e0175945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsayed, H.E.; Ebrahim, H.Y.; Mady, M.S.; Khattab, M.A.; El-Sayed, E.K.; Moharram, F.A. Ethnopharmacological impact of Melaleuca rugulosa (Link) Craven leaves extract on liver inflammation. J. Ethnopharmacol. 2022, 292, 115215. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Rt. | [M+H]+ | [M−H]− | MS2 Fragments (m/z) | Tentative Identification | Class | Ref. |
|---|---|---|---|---|---|---|---|
| 1. | 1.065 | 191.056 | 173, 85 | Quinic acid | Cyclohexanecarboxylic acid | [15] | |
| 2. | 1.117 | 169.014 | 125 | Trihydroxy benzoic acid (gallic acid) * | Phenolic acid | [16] | |
| 3. | 1.143 | 117.019 | 73, 99 | Succinic acid | Omega-dicarboxylic acid | [17] | |
| 4. | 1.144 | 153.019 | 109 | Dihydroxybenzoic acid (protocatechuic acid) | Phenolic acid | [16] | |
| 5. | 1.145 | 181.013 | 137, 113, 109 | Dihydrocaffeic acid [3-(3,4-Dihydroxyphenyl)propionic acid] | Phenolic acid | [16] | |
| 6. | 1.145 | 197.0106 | 179, 153,109 | β,3,4-Trihydroxy benzenepropanoic acid * | Phenolic acid | [18] | |
| 7. | 1.147 | 104.107 | 60, 58 | N-methyl alanine | Amino acid | [19] | |
| 8. | 1.169 | 109.028 | 81 | 1,2-Benzenediol | Phenol dvs. | [20] | |
| 9. | 1.173 | 104.105 | 71, 60, 58 | Choline | Cholines | [21] | |
| 10. | 1.182 | 137.025 | 93 | Salicylic acid* | Phenolic acid | [16] | |
| 11. | 1.185 | 261.045 | 97 | Sorbitol 6-phosphate | Sugar | [22] | |
| 12. | 1.186 | 110.060 | 93, 80, 69 | 4- Aminophenol | Phenol derivative | [23] | |
| 13. | 1.211 | 136.061 | 119, 109, 92, 65 | Adenine | Nucleobase (a purine derivative) | [24] | |
| 14. | 1.237 | 315.071 | 153 | Protocatechuic acid 4-glucoside | Phenolic acid hexoside | [16] | |
| 15. | 1.246 | 173.081 | 129, 36 | Shikimic acid | Cyclohexanecarboxylic acid | [25] | |
| 16. | 1.349 | 167.035 | 152 | Methoxy hydroxy benzoic acid (Vanillic acid) | Phenolic acid | [16] | |
| 17. | 1.351 | 229.151 | 142, 114, 96, 70 | N,N,N-trimethyl-l-alanyl-l-proline betaine | Stilbene | [26] | |
| 18. | 1.380 | 179 | 161, 135 | Caffeic acid * | Phenolic acid | [27] | |
| 19. | 1.401 | 193.073 | 147 | Ferulic acid | Phenolic acid | [16] | |
| 20 | 1.501 | 225.044 | 181, 151 | Dihydrosinapic acid * | Phenolic acid | [28] | |
| 21. | 1.658 | 161.047 | 117, 73 | 6-Hydroxycoumarin | Coumarin | [29] | |
| 22. | 2.212 | 118.086 | 59, 58 | Glycine- betaine | Alpha amino acid | [30] | |
| 23. | 2.232 | 609.127 | 483, 441, 423, 305, 303, 177 | [(epi)gallocatechin-(epi)gallocatechin] * | Procyanidin | [31] | |
| 24. | 3.618 | 593.208 | 425, 407,289 | [(epi)catechin-(epi)gallocatechin] | Procyanidin | [31] | |
| 25. | 3.643 | 593.205 | 425, 407,289, 177 | [(epi)catechin-(epi)gallocatechin] | Procyanidin | [31] | |
| 26. | 3.684 | 593.208 | 425, 407 | [(epi)catechin-(epi)gallocatechin] | Procyanidin | [31] | |
| 27. | 3.815 | 609.240 | 483, 441, 423, 305, 303, 177 | [(epi)gallocatechin-(epi)gallocatechin] * | procyanidin | [31] | |
| 28. | 4.036 | 305.114 | 261, 221, 219, 179, 165 | (epi)gallocatechin | Procyanidin | [32] | |
| 29. | 4.495 | 609.131 | 483, 441, 423, 305, 303, 177 | [(epi)gallocatechin-(epi)gallocatechin]* | Procyanidin | [31] | |
| 30. | 4.515 | 122.079 | Cysteine | Amino acid | [33] | ||
| 31. | 5.150 | 195.065 | 151, 150 | Hydroxycaffeic acid* | Phenolic acid | [34] | |
| 32. | 5.608 | 477.177 | 315, 300, 269 | Isorhamnetin-O-glucoside | Flavone glucoside | [35] | |
| 33. | 5.629 | 137.131 | 119, 110, 64 | Hypoxanthine | Nucleobase (a purine derivative) | [36] | |
| 34. | 5.854 | 405.191 | 243 | 3,4,3′,5′-Tetrahydroxystilbene 3′-glucoside | Stilbene glucoside | [37] | |
| 35. | 5.903 | 177.019 | 149, 133, 109 | 6,7-dihydroxycoumarin (esculetin) | Coumarin | [38] | |
| 36. | 6.424 | 611.166 | 449,465, 303 | Quercetin-rhamnose-hexose | Flavonol glycoside | [39] | |
| 37. | 6.625 | 267.088 | 252, 131 | Formononetin | Isoflavone | [40] | |
| 38. | 6.697 | 197.0091 | 179, 153, 135 | Danshensu (α,3,4-trihydroxy benzenepropanoic acid) | Phenolic acid | [18] | |
| 39. | 6.816 | 593.156 | 285, 284 | Kaempferol-7-neohesperidoside | Flavonol glycoside | [41] | |
| 40. | 6.847 | 465.235 | 303 | Quercetin hexoside | Flavonol glycoside | [42] | |
| 41. | 6.852 | 449.113 | 287 | Okanin-4′-O-glucoside [2′,3′,4′,3,4-pentahydroxychalcone glucoside]* | Chalcone glucoside | [42] | |
| 42. | 6.960 | 623.166 | 315, 314 | Isorhamnetin-3-O-rutinoside (narcissoside)* | Flavonol glycoside | [41] | |
| 43. | 7.011 | 493.200 [M-H], 539.267[M+HCOO] | 493,331,313, 145 | 6-O-p-coumaroyl ajugol | Coumaroyl iridoid glycoside | [43] | |
| 44. | 7.098 | 285.115 | 225 | Agatharesinol | Norlignan | [44] | |
| 45. | 7.293 | 477.106 | 315, 314 | Isorhamnetin-3-O-glucoside* | Flavonol glycoside | [41] | |
| 46. | 7.353 | 289.145 | 245 | (epi)catechin* | Procyanidin | [32] | |
| 47. | 7.412 | 449.108 | 287 | Kaempferol hexoside | Flavonol glycoside | [45] | |
| 48. | 7.451 | 301.107 | 153, 229, 257 | 3, 5, 7-trihydroxy-4′-methoxyflavone (Diosmetin) | Flavonols | [27] | |
| 49. | 7.584 | 433.127 | 271, 85 | Apigenin-O-hexoside | Flavonone -O-glycosides | [46] | |
| 50. | 7.913 | 435.130 | 273, 167, 123 | Phlorizin* | Dihydrochalcone glucoside | [47] | |
| 51. | 7.942 | 509.099 | 347, 332 | Syringetin-3-O-glucoside | Flavonol glycoside | [48] | |
| 52. | 8.024 | (331.191) M+ | 313, 287, 255 | 7-Hydroxy-8-methoxydedihydrorutaecarpine (7-Hydroxy-8-methoxy,7,8-Dehydrorutaecarpine) | Alkaloid | [49] | |
| 53. | 8.092 | 317.029 | 179, 151, 137 | Myricetin* | Flavone | [50] | |
| 54. | 8.192 | 163.076 | 131, 103 | Umbelliferone | Coumarin | [51] | |
| 55. | 8.274 | 177.091 | 162, 149 | Hymecromone (4-methylumbelliferone) | Coumarin | [52] | |
| 56. | 8.296 | 431.099 | 285,284, 255 | luteolin 7-O-rhamnoside | Flavone glycoside | [53] | |
| 57. | 8.375 | 319.045 | 301, 273, 217 | Steviol | Ent-kaurane diterpenoid | [54] | |
| 58. | 8.397 | 347.187 | 303, 301, 285 | 16-Hydroxy-8(17),13-labdadien-15,16-olid-19-oic acid | Diterpenoid acid | [42] | |
| 59. | 9.094 | 377.165 | 359 | Angelol A | Coumarins | [51] | |
| 60. | 9.668 | 541.239[M+HCOO], 495[M-H] | 349,333, 163 | 6-O-p-coumaroyl dihydroajugol | Coumaroyl iridoid glycoside | [43] | |
| 61. | 9.704 | 303.123 | 259, 285 | Copalic acid* | Diterpenoid acid | [55] | |
| 62. | 9.771 | 303.050 | 257, 201 | Abietic acid | Diterpenoid acid | [56] | |
| 63. | 10.399 | 301.216 | 283, 255 | Retinoic acid | Retinoids | [57] | |
| 64. | 10.934 | 301.217 | 255, 173, 147,133, 109 | Dehydroabietic acid | Diterpenoid acid | [56] | |
| 65. | 11.015 | 305.174 | 287, 259, 159 | Taxifolin (dihydroquercetin) | Flavanonols | [58] | |
| 66. | 11.106 | 287.055 | 269, 153 | Luteolin | Flavones | [27] | |
| 67. | 11.401 | 347.223 | 303, 301 | 15-Hydroxypinusolidic acid | Diterpenoid acid | [42] | |
| 68. | 11.814 | 377.143 | 359 | Isoangelol | Coumarins | [51] | |
| 69. | 12.278 | 897.071 | 693, 289 | (epi)gallocatechin-(epi)gallocatechin-(epi)catechin | Procyanidin | [31] | |
| 70. | 12.419 | 577.117 | 289 | (epi)catechin-(epi)catechin | Procyanidin | [32] | |
| 71. | 12.57 | 305.212 | 261, 287 | 3-hydroxy-(13S)-l6-nor-pimar-7-en-l5-oic acid * | Diterpenoid acid | [44] | |
| 72. | 12.658 | 331.189 | 203 | 15-hydroxy-7-oxo-dehydroabietic acid | Diterpenoid acid | [56] | |
| 73. | 12.711 | 329.177 | 285,311 | Carnosol | Phenolic diterpene | [55] | |
| 74. | 12.788 | 301.0349 | 255, 179, 151 | Quercetin* | Flavonol | [32] | |
| 75. | 13.014 | 565.113 | 389 | Isoginkgetin | Biflavonoids | [59] | |
| 76. | 13.058 | 315.161 | 199, 187 | 7-oxo-dehydroabietic acid | Diterpenoid acid | [56] | |
| 77. | 13.36 | 283.061 | 268 | Chrysin-6-methyl-ether | Flavonone | [60] | |
| 78. | 13.449 | 283.061 | 268,239 | Physcion | dihydroxyanthraquinone | [61] | |
| 79. | 14.413 | 331.229 | 287 | Pinusolidic acid * | Diterpenoid acid | [42] | |
| 80. | 14.567 | 271.228 | 253,227, 225 | Hydroxypalmitic Acid | Fatty acid | [55] | |
| 81. | 14.76 | 299.166 | 284, 255 | 4′-Hydroxywogonin | Trihydroxy-methoxyflavone | [29] | |
| 82. | 15.382 | 539.168 | 539, 387 | 2″,3″-Dihydro-3′,3‴-biapigenin | Biflavonoids | [59] | |
| 83. | 15.602 | 536.883 | 537, 385 | 2′,8″-Biapigenin | Biflavonoids | [59] | |
| 84. | 15.618 | 565.114 | 445, 388, 403, 456, 471 | Robustaflavone 7,4′-dimethyl ether | Biflavonoids | [59] | |
| 85. | 15.911 | 537.305 | 537, 193, 192 | Delicaflavone | Biflavonoids | [59] | |
| 86. | 15.929 | 303.229 | 285, 257, 239 | Abietic acid isomer | Diterpenoid acid | [56] | |
| 87. | 16.014 | 301.216 | 283, 255, 133, 109 | Dehydroabietic acid isomer | Diterpenoid acid | [56] | |
| 88. | 17.120 | 313.179 | 211, 197, 185 | Dehydrated 15-hydroxy-7-oxo-dehydroabietic acid | Diterpenoid acid | [56] | |
| 89. | 17.17 | 363.212 [M+HCOO] | 317, 159 | Agarotetrol formate adduct | 2-(2-phenylethyl) chromones | [62] | |
| 90 | 17.679 | 315.195 | 199, 187 | 7-oxo-dehydroabietic acid isomer | Diterpenoid acid | [56] | |
| 91. | 19.553 | 303.199 | 285, 257, 109 | Abietic acid isomer | Diterpenoid acid | [56] | |
| 92. | 20.795 | 301.217 | 255, 173, 147,133, 109 | Dehydroabietic acid isomer | Diterpenoid acid | [56] | |
| 93. | 20.857 | 301.216 | 255, 173,133, 109 | Dehydroabietic acid isomer | Diterpenoid acid | [56] | |
| 94. | 20.883 | 301.217 | 255, 173, 147,133, 109 | Dehydroabietic acid isomer | Diterpenoid acid | [56] | |
| 95. | 22.529 | 315.196 | 297,199, 187 | 7-oxo-dehydroabietic acid isomer | Diterpenoid acid | [56] |
| Rank | Target Name | Score |
|---|---|---|
| 1 | Narcissoside | 60 |
| 2 | Isorhamnetin-3-O-glucoside | 58 |
| 2 | Syringetin-3-O-glucoside | 58 |
| 4 | 6-O-p-coumaroyl ajugol | 56 |
| 5 | Luteolin 7-rhamnoside | 55 |
| 6 | Robustaflavone 7,4′-dimethyl ether | 53 |
| 6 | Ferulic acid | 53 |
| 6 | 2″,3″-Dihydro-3′,3‴-biapigenin | 53 |
| 9 | 3-hydroxy-(13S)-16-nor-pimar-7-en-15-oic acid | 52 |
| 9 | Caffeic acid | 52 |
| 11 | Agatharesinol | 51 |
| 12 | Delicaflavone | 50 |
| 12 | Dehydrated 15-hydroxy-7-oxo-dehydroabietic acid | 50 |
| 12 | Diosmetin | 50 |
| 15 | Esculetin | 49 |
| 15 | Hydroxycaffeic acid | 49 |
| 17 | Hymecromone | 48 |
| 17 | Myricetin | 48 |
| 17 | Isoginkgetin | 48 |
| 17 | 16-Hydroxy-8(17)13-labdadien-1516-olid-19-oic acid | 48 |
| 21 | Quercetin | 47 |
| 21 | [(epi)gallocatechin-(epi)gallocatechin] | 47 |
| 21 | Luteolin | 47 |
| 21 | Umbelliferone | 47 |
| 25 | 15-hydroxy-7-oxo-dehydroabietic acid | 46 |
| 25 | 7-oxo-dehydroabietic acid | 46 |
| 27 | Phlorizin | 45 |
| 27 | Protocatechuic acid 4-glucoside | 45 |
| 29 | 6-Hydroxycoumarin  | 44 |
| 29 | 3,4-Trihydroxy benzenepropanoic acid | 44 |
| 29 | Dehydroabietic acid | 44 |
| 32 | [(epi)catechin-(epi)gallocatechin] | 43 |
| 32 | Gallic acid | 43 |
| 34 | Pinusolidic acid | 42 |
| 35 | Hydroxypalmitic Acid | 41 |
| 35 | 15-Hydroxypinusolidic acid | 41 |
| 37 | Carnosol | 40 |
| Component | S Score Kcal/mol | H-Bond Interactions | Pi-H Interactions |
|---|---|---|---|
| 7-oxo-dehydroabietic acid | −6.5439 | THR184 | ASN51 |
| Caffeic acid | −5.1359 | SER52 | - |
| Narcissoside (Isorhamnetin-3-O-rutinoside) | −8.4051 | ASP102 | ASN106 |
| Isorhamnetin-3-O-glucoside | −7.6758 | ASP93 | - |
| Syringetin-3-O-glucoside | −8.0354 | ASP93 | ASN106 |
| 15-hydroxy-7-oxo-dehydroabietic acid | −5.5907 | MET98, PHE 138 | - |
| 6-O-p-coumaroyl ajugol | −7.5511 | GLU47, ASN 51 | - |
| Luteolin 7-rhamnoside | −7.1269 | GLY97 | ASN106 |
| Robustaflavone 7,4’-dimethyl ether | −7.0831 | MET98 | - |
| Ferulic acid | −5.0937 | ASN51 | - |
| T5M | −6.4559 | THR184 | ASN51 |
| Component | S Score Kcal/mol | H-Bond Interactions | Pi-H Interactions |
|---|---|---|---|
| 7-oxo-dehydroabietic acid | −5.7797 | MET769, THR830 | GLY772 |
| Caffeic acid | −5.2224 | MET742, GLN767, ASP831 | - |
| Narcissoside (Isorhamnetin-3-O-rutinoside) | −9.0112 | MET742, MET769, ASP 831 | LEU694 |
| Isorhamnetin-3-O-glucoside | −8.42596 | MET742, ASP 831 | - |
| Syringetin-3-O-glucoside | −8.5239 | MET742, ARG817, ASP 831 | LEU694 |
| 15-hydroxy-7-oxo-dehydroabietic acid | −5.9105 | LYS721 | VAL702 |
| 6-O-p-coumaroyl ajugol | −7.4587 | MET742, GLU780 | - |
| Luteolin 7-rhamnoside | −7.2894 | THR766, ASP 831 | LEU694, GLY772 |
| Robustaflavone 7,4′-dimethyl ether | −8.4544 | LYS721 | LEU694, VAL 702 |
| Ferulic acid | −5.1249 | MET769 | - |
| erlotinib | −7.76417 | VAL702, MET769, | - |
| Component | S Score Kcal/mol | H-Bond Interactions | Pi–H Interactions |
|---|---|---|---|
| 7-oxo-dehydroabietic acid | −4.9993 | GLY262 | - |
| Caffeic acid | −4.5554 | SER99 | THR256 |
| Narcissoside (Isorhamnetin-3-O-rutinoside) | −7.1544 | SER96 | PRO 98 |
| Isorhamnetin-3-O-glucoside | −7.1453 | ASP208 | THR256, LEU 264 |
| Syringetin-3-O-glucoside | −6.7796 | SER96, ASP 208, GLY262 | PRO98 |
| 15-hydroxy-7-oxo-dehydroabietic acid | −5.0645 | ASP208 | - |
| 6-O-p-coumaroyl ajugol | −6.7463 | GLU 204, ILE 254 | - |
| Luteolin 7-rhamnoside | −5.9100 | SER260, ARG 267 | - |
| Robustaflavone 7,4′-dimethyl ether | −6.6223 | SER99, ARG 158, ASP 208 | THR256 |
| Ferulic acid | −4.5974 | SER99, GLY 262 | THR256 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamed, M.E.; Tawfeek, N.; Elbaramawi, S.S.; Elbatreek, M.H.; Fikry, E. Agathis robusta Bark Extract Protects from Renal Ischemia-Reperfusion Injury: Phytochemical, In Silico and In Vivo Studies. Pharmaceuticals 2022, 15, 1270. https://doi.org/10.3390/ph15101270
Mohamed ME, Tawfeek N, Elbaramawi SS, Elbatreek MH, Fikry E. Agathis robusta Bark Extract Protects from Renal Ischemia-Reperfusion Injury: Phytochemical, In Silico and In Vivo Studies. Pharmaceuticals. 2022; 15(10):1270. https://doi.org/10.3390/ph15101270
Chicago/Turabian StyleMohamed, Maged E., Nora Tawfeek, Samar S. Elbaramawi, Mahmoud H. Elbatreek, and Eman Fikry. 2022. "Agathis robusta Bark Extract Protects from Renal Ischemia-Reperfusion Injury: Phytochemical, In Silico and In Vivo Studies" Pharmaceuticals 15, no. 10: 1270. https://doi.org/10.3390/ph15101270