
Caulis Polygoni Multiflori Accelerates Megakaryopoiesis and Thrombopoiesis via Activating PI3K/Akt and MEK/ERK Signaling Pathways
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Characterization of CPM
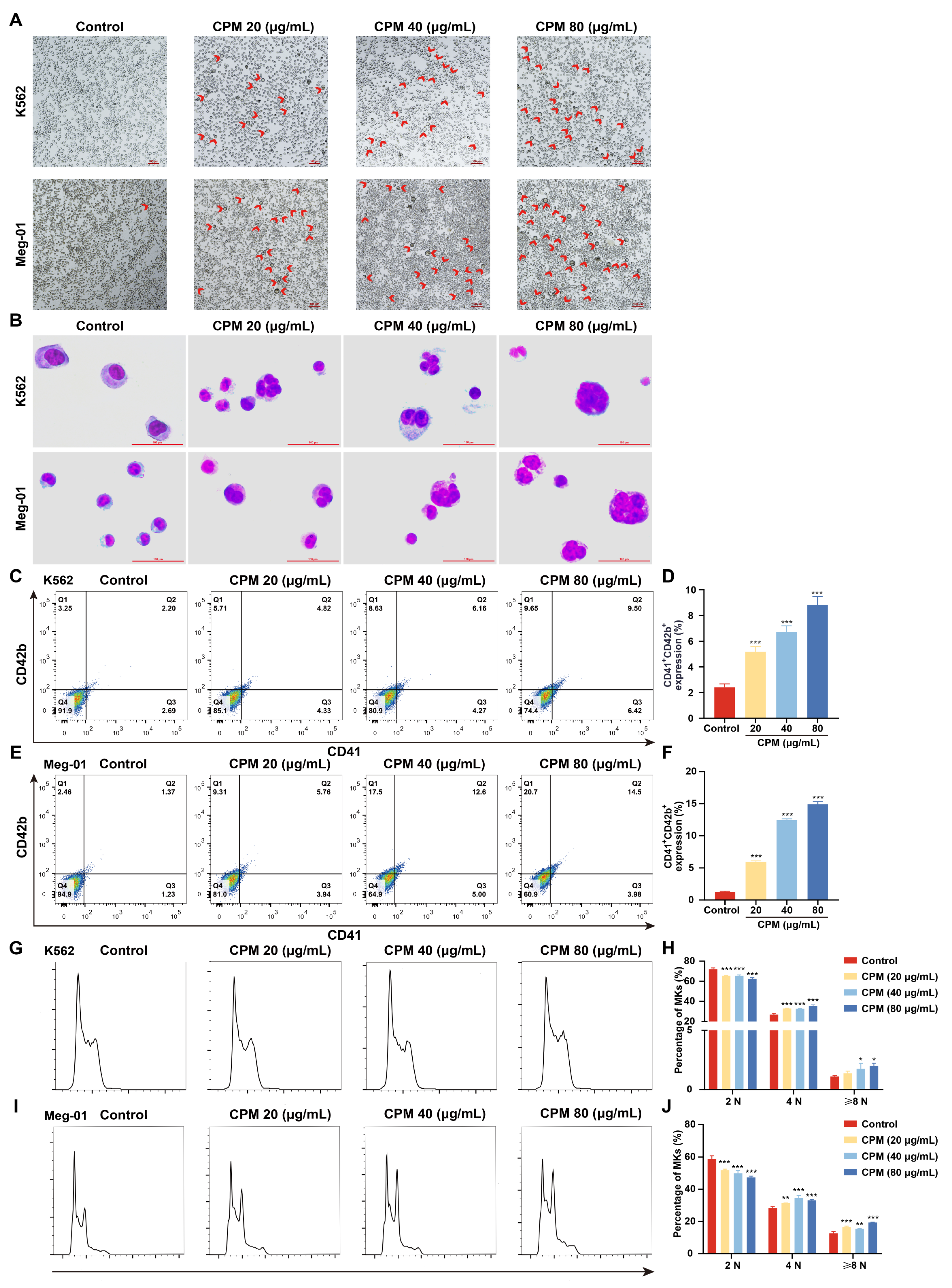
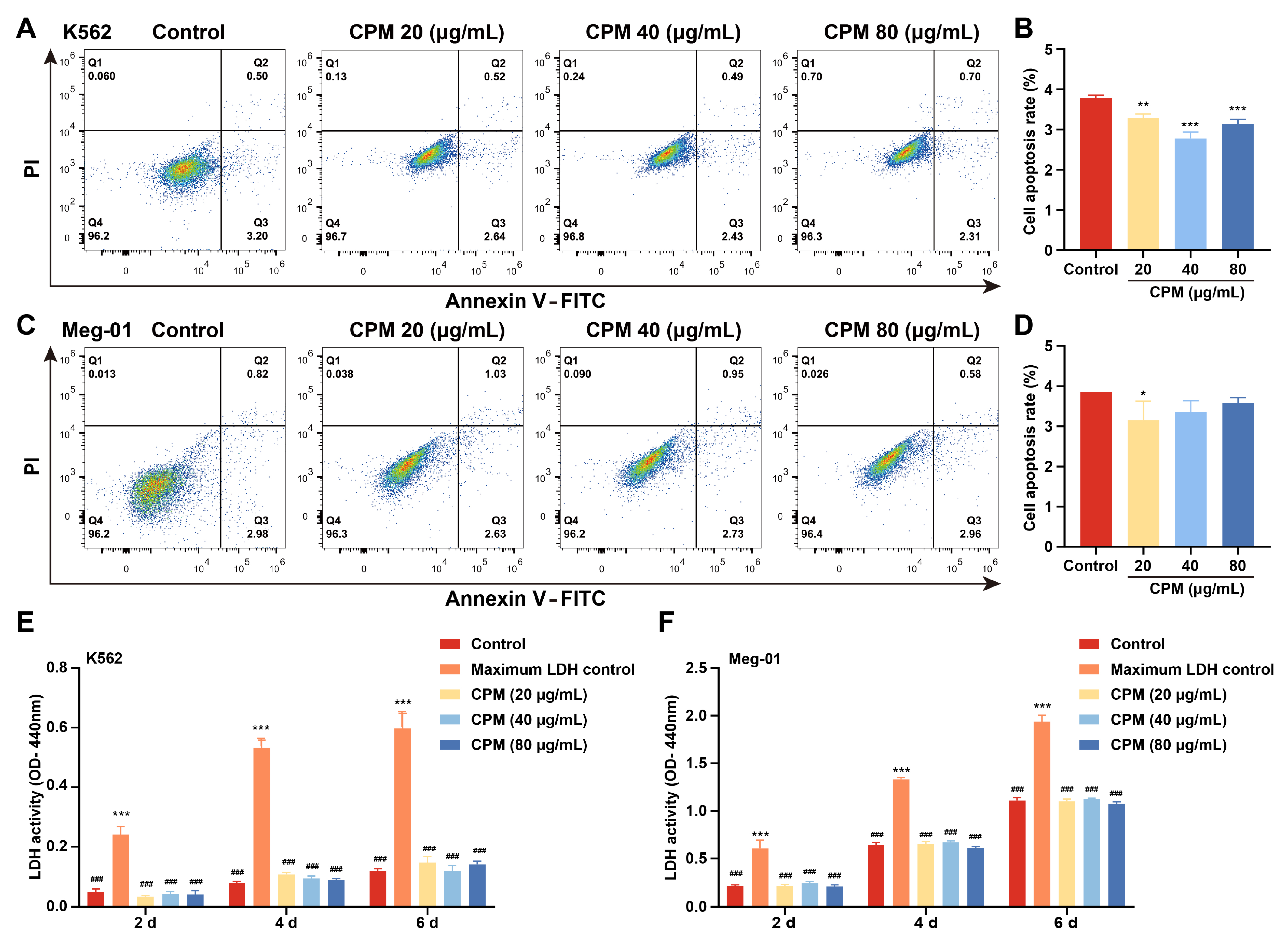
2.2. CPM induces MK Differentiation In Vitro
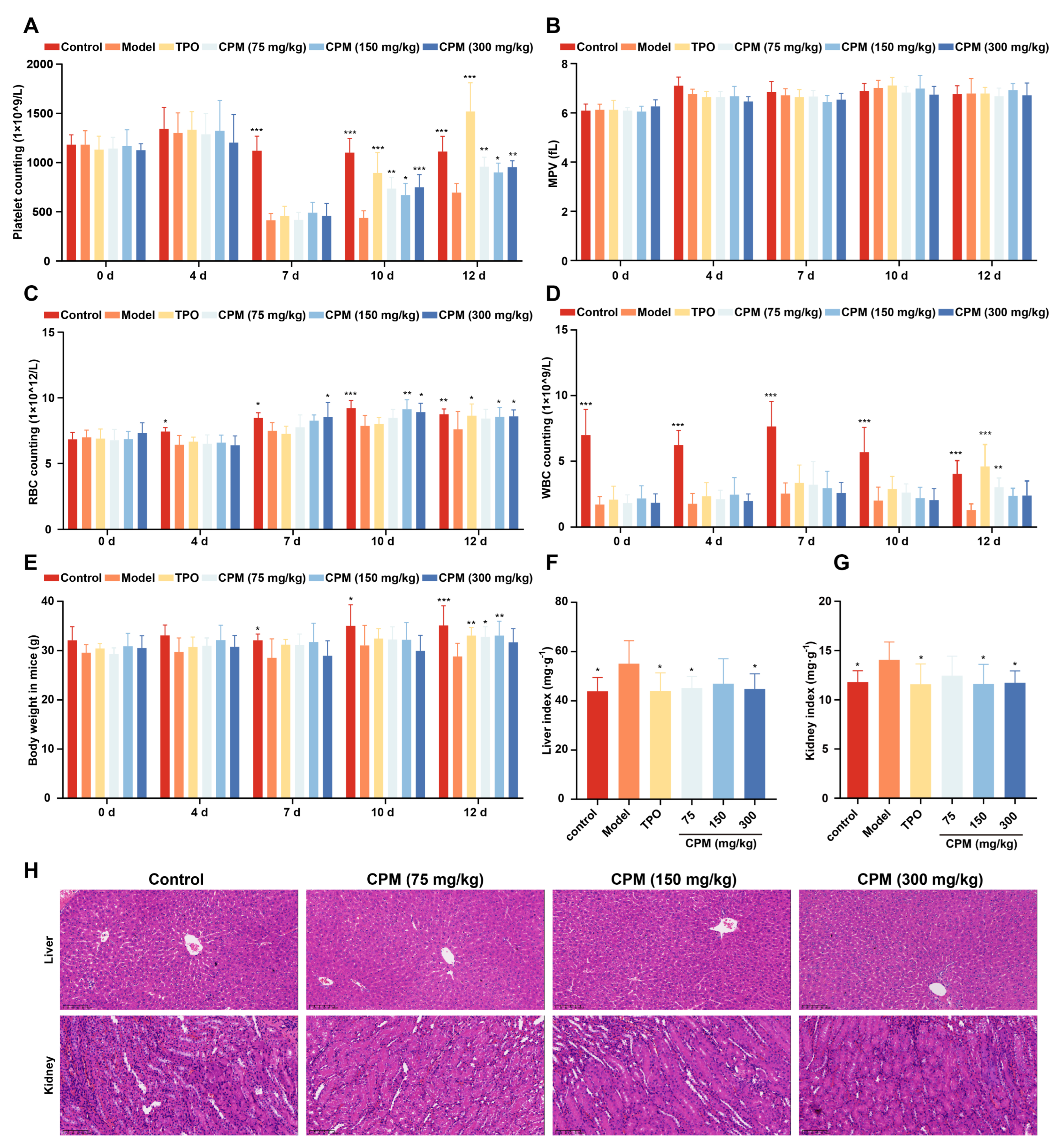
2.3. The Therapeutic Effects of CPM on Myelosuppressed Mice
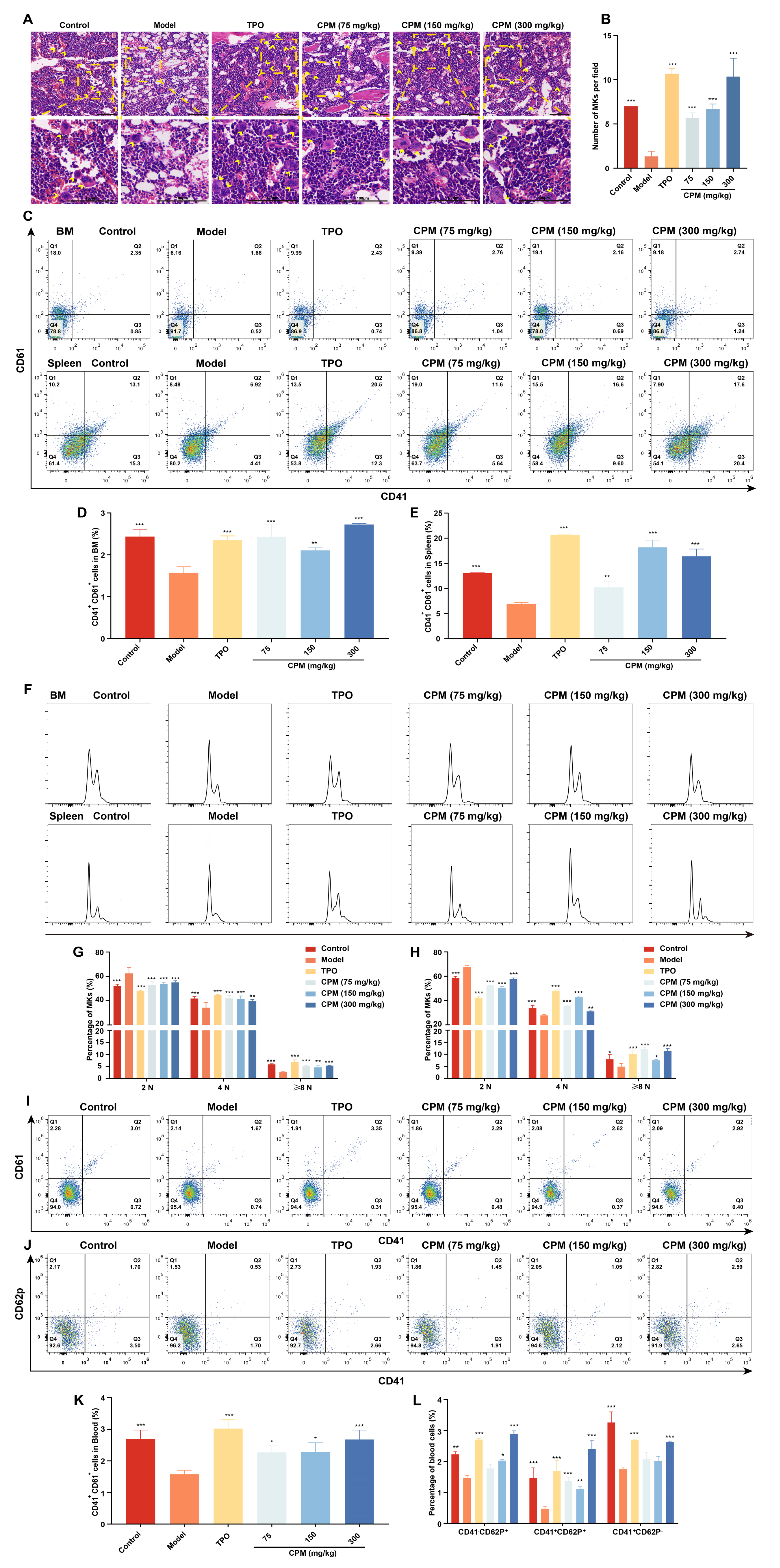
2.4. CPM Promotes Megakaryopoiesis and Platelet Activation In Vivo
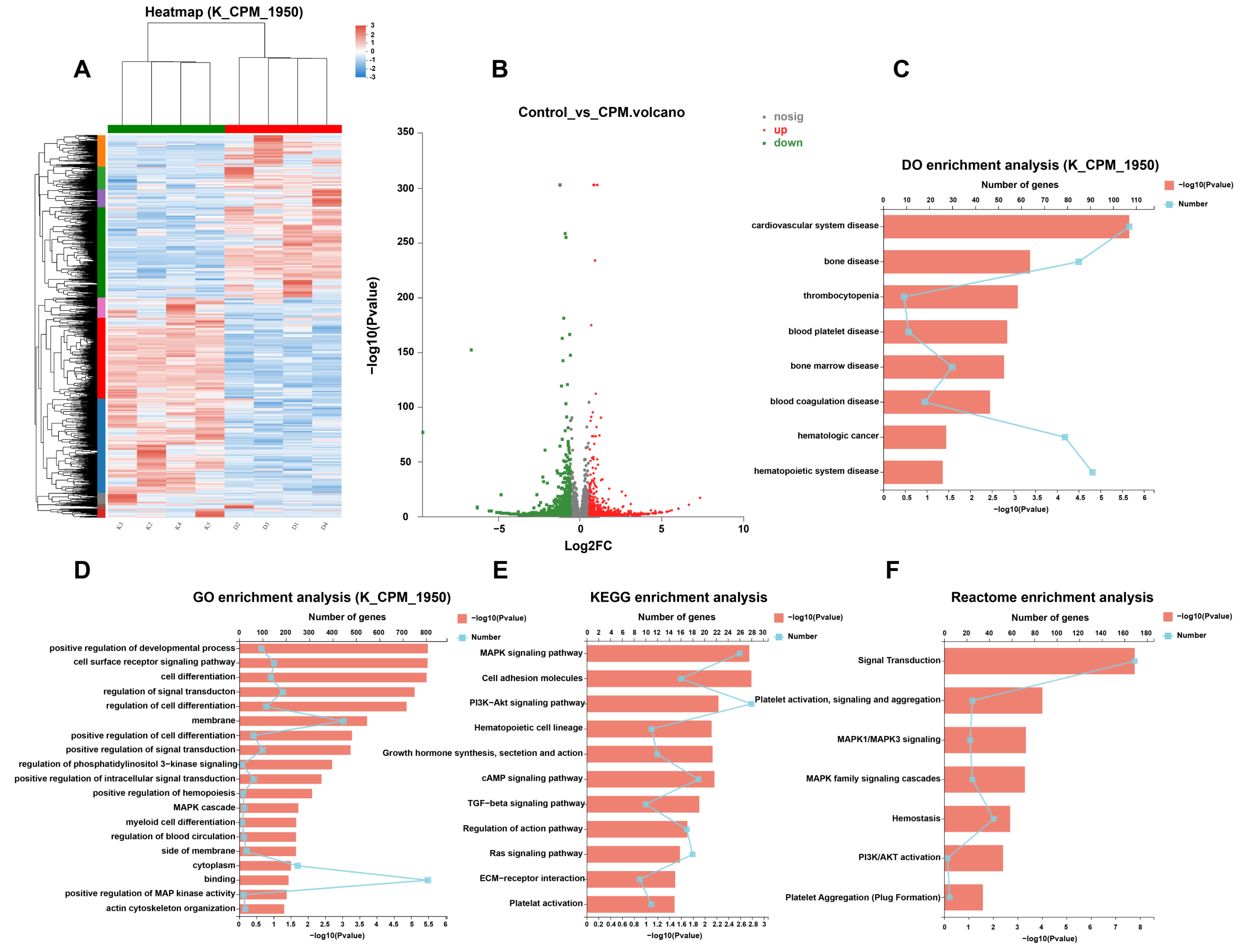
2.5. Gene Expression Profile in MK Differentiation Induced by CPM
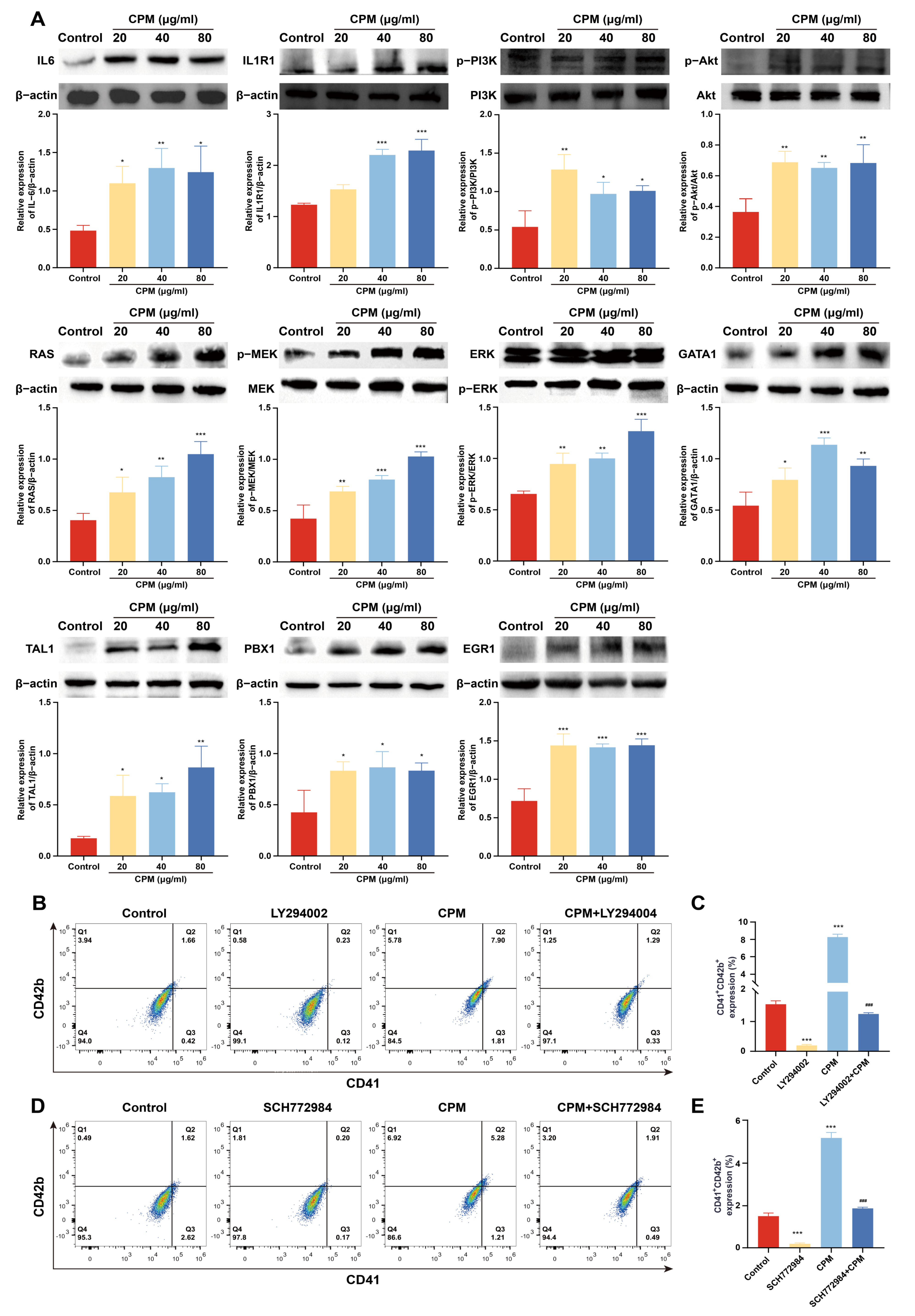
2.6. CPM Induces MK Differentiation through PI3K/Akt and MEK/ERK (MAPK) Signaling Pathways
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Sample Preparation
4.3. UPLC-MS Conditions
4.4. Cell Culture
4.5. LDH Assay
4.6. Cell Apoptosis Assay
4.7. Cell Morphological Observation
4.8. Giemsa Staining
4.9. Surface Marker Analysis
4.10. Polyploidy Analysis
4.11. Animals
4.12. Routine Blood Test
4.13. Visceral Index
4.14. Histology Analysis
4.15. Flow Cytometry Analysis of PB Cells
4.16. Flow Cytometry Analysis of BM and Spleen Cells
4.17. RNA-Seq and Data Analysis
4.18. Differential Expression Analysis and Functional Enrichment
4.19. Western Blotting
4.20. Statistical Analysis
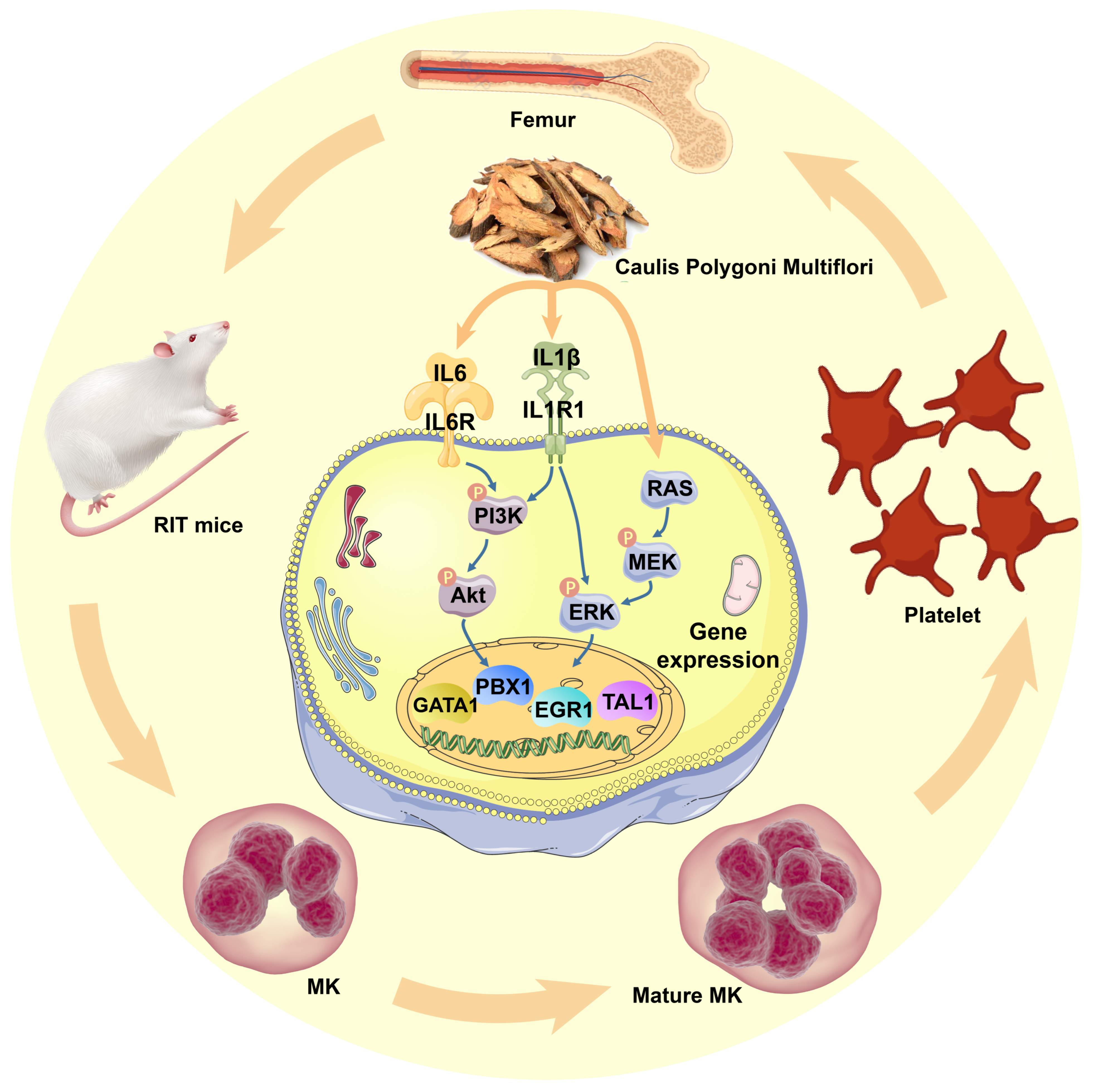
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Soerjomataram, I.; Bray, F. Planning for tomorrow: Global cancer incidence and the role of prevention 2020–2070. Nat. Rev. Clin. Oncol. 2021, 18, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Baskar, R.; Lee, K.A.; Yeo, R.; Yeoh, K.W. Cancer and radiation therapy: Current advances and future directions. Int. J. Med. Sci. 2012, 9, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Schaue, D.; McBride, W.H. Opportunities and challenges of radiotherapy for treating cancer. Nat. Rev. Clin. Oncol. 2015, 12, 527–540. [Google Scholar] [CrossRef]
- Razzaghdoust, A.; Mofid, B.; Zangeneh, M. Predicting chemotherapy-induced thrombocytopenia in cancer patients with solid tumors or lymphoma. J. Oncol. Pharm. Pract. Off. Publ. Int. Soc. Oncol. Pharm. Pract. 2020, 26, 587–594. [Google Scholar] [CrossRef]
- Tkaczynski, E.; Arulselvan, A.; Tkaczynski, J.; Avery, S.; Xiao, L.; Torok-Storb, B.; Abrams, K.; Rao, N.V.; Johnson, G.; Kennedy, T.P.; et al. 2-O, 3-O desulfated heparin mitigates murine chemotherapy- and radiation-induced thrombocytopenia. Blood Adv. 2018, 2, 754–761. [Google Scholar] [CrossRef]
- Machlus, K.R.; Italiano, J.E., Jr. The incredible journey: From megakaryocyte development to platelet formation. J. Cell Biol. 2013, 201, 785–796. [Google Scholar] [CrossRef]
- Erickson-Miller, C.L.; Ji, H.; Murphy, M.J., Jr. Megakaryocytopoiesis and platelet production: Does stem cell factor play a role? Stem Cells 1993, 11 (Suppl. 2), 163–169. [Google Scholar] [CrossRef]
- Metcalf, D.; Hilton, D.; Nicola, N.A. Leukemia inhibitory factor can potentiate murine megakaryocyte production in vitro. Blood 1991, 77, 2150–2153. [Google Scholar] [CrossRef]
- Behrens, K.; Alexander, W.S. Cytokine control of megakaryopoiesis. Growth Factors 2018, 36, 89–103. [Google Scholar] [CrossRef]
- Yang, M.; Chesterman, C.N.; Chong, B.H. Recombinant PDGF enhances megakaryocytopoiesis in vitro. Br. J. Haematol. 1995, 91, 285–289. [Google Scholar] [CrossRef]
- Greenberg, E.M.; Kaled, E.S. Thrombocytopenia. Crit. Care Nurs. Clin. N. Am. 2013, 25, 427–434. [Google Scholar] [CrossRef]
- Eto, K.; Kunishima, S. Linkage between the mechanisms of thrombocytopenia and thrombopoiesis. Blood 2016, 127, 1234–1241. [Google Scholar] [CrossRef]
- Kaye, J.A. The clinical development of recombinant human interleukin 11 (NEUMEGA rhIL-11 growth factor). Stem Cells 1996, 14 (Suppl. 1), 256–260. [Google Scholar] [CrossRef]
- Vadhan-Raj, S. Clinical experience with recombinant human thrombopoietin in chemotherapy-induced thrombocytopenia. Semin. Hematol. 2000, 37 (Suppl. 4), 28–34. [Google Scholar] [CrossRef]
- Al-Samkari, H.; Soff, G.A. Clinical challenges and promising therapies for chemotherapy-induced thrombocytopenia. Expert Rev. Hematol. 2021, 14, 437–448. [Google Scholar] [CrossRef]
- Yang, B.B.; Doshi, S.; Arkam, K.; Franklin, J.; Chow, A.T. Development of Romiplostim for Treatment of Primary Immune Thrombocytopenia From a Pharmacokinetic and Pharmacodynamic Perspective. Clin. Pharmacokinet. 2016, 55, 1045–1058. [Google Scholar] [CrossRef]
- Clemons Bankston, P.; Al-Horani, R.A. New Small Molecule Drugs for Thrombocytopenia: Chemical, Pharmacological, and Therapeutic Use Considerations. Int. J. Mol. Sci. 2019, 20, 3013. [Google Scholar] [CrossRef]
- Li, J.; Yang, C.; Xia, Y.; Bertino, A.; Glaspy, J.; Roberts, M.; Kuter, D.J. Thrombocytopenia caused by the development of antibodies to thrombopoietin. Blood 2001, 98, 3241–3248. [Google Scholar] [CrossRef]
- Basser, R.L.; O’Flaherty, E.; Green, M.; Edmonds, M.; Nichol, J.; Menchaca, D.M.; Cohen, B.; Begley, C.G. Development of pancytopenia with neutralizing antibodies to thrombopoietin after multicycle chemotherapy supported by megakaryocyte growth and development factor. Blood 2002, 99, 2599–2602. [Google Scholar] [CrossRef] [Green Version]
- Virk, Z.M.; Kuter, D.J.; Al-Samkari, H. An evaluation of avatrombopag for the treatment of thrombocytopenia. Expert Opin. Pharmacother. 2021, 22, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Tjepkema, M.; Amini, S.; Schipperus, M. Risk of thrombosis with thrombopoietin receptor agonists for ITP patients: A systematic review and meta-analysis. Crit. Rev. Oncol./Hematol. 2022, 171, 103581. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, W.B.; Bussel, J.B. Thrombopoietin receptor agonists: A critical review. Semin. Hematol. 2015, 52, 46–52. [Google Scholar] [CrossRef] [PubMed]
- McElroy, P.L.; Wei, P.; Buck, K.; Sinclair, A.M.; Eschenberg, M.; Sasu, B.; Molineux, G. Romiplostim promotes platelet recovery in a mouse model of multicycle chemotherapy-induced thrombocytopenia. Exp. Hematol. 2015, 43, 479–487. [Google Scholar] [CrossRef]
- Winer, E.S.; Safran, H.; Karaszewska, B.; Bauer, S.; Khan, D.; Doerfel, S.; Burgess, P.; Kalambakas, S.; Mostafa Kamel, Y.; Forget, F. Eltrombopag for thrombocytopenia in patients with advanced solid tumors receiving gemcitabine-based chemotherapy: A randomized, placebo-controlled phase 2 study. Int. J. Hematol. 2017, 106, 765–776. [Google Scholar] [CrossRef]
- Busch, M.P.; Bloch, E.M.; Kleinman, S. Prevention of transfusion-transmitted infections. Blood 2019, 133, 1854–1864. [Google Scholar] [CrossRef]
- Burnouf, T.; Elemary, M.; Radosevic, J.; Seghatchian, J.; Goubran, H. Platelet transfusion in thrombocytopenic cancer patients: Sometimes justified but likely insidious. Transfus. Apher. Sci. Off. J. World Apher. Assoc. Off. J. Eur. Soc. Haemapheresis 2017, 56, 305–309. [Google Scholar] [CrossRef]
- Blumberg, N.; Heal, J.M.; Phillips, G.L. Platelet transfusions: Trigger, dose, benefits, and risks. F1000 Med. Rep. 2010, 2, 5. [Google Scholar] [CrossRef]
- Yang, M.; Chan, G.C.; Deng, R.; Ng, M.H.; Cheng, S.W.; Lau, C.P.; Ye, J.Y.; Wang, L.; Liu, C. An herbal decoction of Radix astragali and Radix angelicae sinensis promotes hematopoiesis and thrombopoiesis. J. Ethnopharmacol. 2009, 124, 87–97. [Google Scholar] [CrossRef]
- Liu, C.; Li, J.; Meng, F.Y.; Liang, S.X.; Deng, R.; Li, C.K.; Pong, N.H.; Lau, C.P.; Cheng, S.W.; Ye, J.Y.; et al. Polysaccharides from the root of Angelica sinensis promotes hematopoiesis and thrombopoiesis through the PI3K/AKT pathway. BMC Complement. Altern. Med. 2010, 10, 79. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Xu, W.; Yi, C.; Cheng, Y.; Xin, H.; Xue, H.; Li, C.K.; Fang, X.; Yang, L.; Chen, C.; et al. Astragalus polysaccharide has a protective effect on hematopoiesis in an irradiated mouse model and decreases apoptosis in megakaryocytes. Mol. Med. Rep. 2021, 23, 15. [Google Scholar] [CrossRef]
- Chen, C.J.; Liu, X.; Chiou, J.S.; Hang, L.W.; Li, T.M.; Tsai, F.J.; Ko, C.H.; Lin, T.H.; Liao, C.C.; Huang, S.M.; et al. Effects of Chinese herbal medicines on dementia risk in patients with sleep disorders in Taiwan. J. Ethnopharmacol. 2021, 264, 113267. [Google Scholar] [CrossRef]
- National Health Commission of the People’s Republic of China. Available online: http://www.nhc.gov.cn/wjw/gfxwj/201304/e33435ce0d894051b15490aa3219cdc4.shtml (accessed on 10 July 2022).
- Zhao, Y.; Kao, C.P.; Chang, Y.S.; Ho, Y.L. Quality assessment on Polygoni Multiflori Caulis using HPLC/UV/MS combined with principle component analysis. Chem. Cent. J. 2013, 7, 106. [Google Scholar] [CrossRef]
- Fattorusso, R.; Frutos, S.; Sun, X.; Sucher, N.J.; Pellecchia, M. Traditional Chinese medicines with caspase-inhibitory activity. Phytomed. Int. J. Phytother. Phytopharm. 2006, 13, 16–22. [Google Scholar] [CrossRef]
- Meng, X.; Xiong, X.J. Traditional Chinese medicine insights of newly-diagnosed and young hypertension and clinical practice of Tianma Gouteng Decoction for hypertension treatment. Zhongguo Zhong Yao Za Zhi Zhongguo Zhongyao Zazhi China J. Chin. Mater. Med. 2020, 45, 2752–2759. [Google Scholar]
- Feng, S.; Song, L.; Liu, Y.; Lai, F.; Zuo, G.; He, G.; Chen, M.; Huang, D. Hypoglycemic activities of commonly-used traditional Chinese herbs. Am. J. Chin. Med. 2013, 41, 849–864. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia Commission. Chinese Pharmacopoeia, 11th ed.; China Medical Science Press: Beijing, China, 2020; pp. 277–278. [Google Scholar]
- Nanjing University of Chinese Medicine. Zhong Yao Daci Dian, 2nd ed.; Shanghai Scientific & Technical Publishers: Shanghai, China, 2015; pp. 2033–2034. [Google Scholar]
- Zhang, Z.; Zhang, Y.; Gao, M.; Cui, X.; Yang, Y.; van Duijn, B.; Wang, M.; Hu, Y.; Wang, C.; Xiong, Y. Steamed Panax notoginseng Attenuates Anemia in Mice With Blood Deficiency Syndrome via Regulating Hematopoietic Factors and JAK-STAT Pathway. Front. Pharmacol. 2019, 10, 1578. [Google Scholar] [CrossRef]
- Akin, S.; Haznedaroglu, I.C. Clinical variations between three different causes of thrombocytopenia. Niger. J. Clin. Pract. 2021, 24, 17–20. [Google Scholar]
- Almiş, H.; Bucak, I.H.; Çelik, V.; Tekin, M.; Karakoç, F.; Konca, Ç.; Turgut, M. Mean platelet volume in hepatitis A. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 2310–2314. [Google Scholar]
- Mac Manus, M.; Lamborn, K.; Khan, W.; Varghese, A.; Graef, L.; Knox, S. Radiotherapy-associated neutropenia and thrombocytopenia: Analysis of risk factors and development of a predictive model. Blood 1997, 89, 2303–2310. [Google Scholar] [CrossRef]
- Kuter, D.J. Managing thrombocytopenia associated with cancer chemotherapy. Oncology 2015, 29, 282–294. [Google Scholar]
- Sun, X.; Zhao, Y.N.; Qian, S.; Gao, R.L.; Yin, L.M.; Wang, L.P.; Chong, B.H.; Zhang, S.Z. Ginseng-Derived Panaxadiol Saponins Promote Hematopoiesis Recovery in Cyclophosphamide-Induced Myelosuppressive Mice: Potential Novel Treatment of Chemotherapy-Induced Cytopenias. Chin. J. Integr. Med. 2018, 24, 200–206. [Google Scholar] [CrossRef]
- Alitalo, R. Induced differentiation of K562 leukemia cells: A model for studies of gene expression in early megakaryoblasts. Leuk. Res. 1990, 14, 501–514. [Google Scholar] [CrossRef]
- Schweinfurth, N.; Hohmann, S.; Deuschle, M.; Lederbogen, F.; Schloss, P. Valproic acid and all trans retinoic acid differentially induce megakaryopoiesis and platelet-like particle formation from the megakaryoblastic cell line MEG-01. Platelets 2010, 21, 648–657. [Google Scholar] [CrossRef]
- Lanza, F.; Gachet, C.; Eckly, A. In Vitro and In Vivo Methods to Explore Megakaryopoiesis. J. Vis. Exp. JoVE 2021, 177. [Google Scholar] [CrossRef]
- Inra, C.N.; Zhou, B.O.; Acar, M.; Murphy, M.M.; Richardson, J.; Zhao, Z.; Morrison, S.J. A perisinusoidal niche for extramedullary haematopoiesis in the spleen. Nature 2015, 527, 466–471. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, W.Z.; Liu, T.; Feng, X.; Yang, N.; Zhou, H.F. Signaling pathway of MAPK/ERK in cell proliferation, differentiation, migration, senescence and apoptosis. J. Recept. Signal Transduct. Res. 2015, 35, 600–604. [Google Scholar] [CrossRef]
- Yu, J.S.; Cui, W. Proliferation, survival and metabolism: The role of PI3K/AKT/mTOR signalling in pluripotency and cell fate determination. Development 2016, 143, 3050–3060. [Google Scholar] [CrossRef]
- Gabbouj, S.; Ryhänen, S.; Marttinen, M.; Wittrahm, R.; Takalo, M.; Kemppainen, S.; Martiskainen, H.; Tanila, H.; Haapasalo, A.; Hiltunen, M.; et al. Altered Insulin Signaling in Alzheimer’s Disease Brain—Special Emphasis on PI3K-Akt Pathway. Front. Neurosci. 2019, 13, 629. [Google Scholar] [CrossRef]
- Yang, J.; Nie, J.; Ma, X.; Wei, Y.; Peng, Y.; Wei, X. Targeting PI3K in cancer: Mechanisms and advances in clinical trials. Mol. Cancer 2019, 18, 26. [Google Scholar] [CrossRef]
- Qu, T.; Gao, Y.; Li, A.; Li, Z.; Qin, X. Systems biology analysis of the effect and mechanism of total flavonoids of Astragali Radix against cyclophosphamide-induced leucopenia in mice. J. Pharm. Biomed. Anal. 2021, 205, 114357. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Choi, E.J. Pathological roles of MAPK signaling pathways in human diseases. Biochim. Biophys. Acta 2010, 1802, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Kirito, K.; Kaushansky, K. Transcriptional regulation of megakaryopoiesis: Thrombopoietin signaling and nuclear factors. Curr. Opin. Hematol. 2006, 13, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, T.; Liu, S.; Mo, Q.; Jiang, N.; Chen, Q.; Yang, J.; Han, Y.W.; Chen, J.P.; Huang, F.H.; et al. Discovery of a novel megakaryopoiesis enhancer, ingenol, promoting thrombopoiesis through PI3K-Akt signaling independent of thrombopoietin. Pharmacol. Res. 2022, 177, 106096. [Google Scholar] [CrossRef]
- Mortazavi Farsani, S.S.; Sadeghizadeh, M.; Gholampour, M.A.; Safari, Z.; Najafi, F. Nanocurcumin as a novel stimulator of megakaryopoiesis that ameliorates chemotherapy-induced thrombocytopenia in mice. Life Sci. 2020, 256, 117840. [Google Scholar] [CrossRef]
- Noetzli, L.J.; French, S.L.; Machlus, K.R. New Insights Into the Differentiation of Megakaryocytes From Hematopoietic Progenitors. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1288–1300. [Google Scholar] [CrossRef]
- Burstein, S.A. Effects of interleukin 6 on megakaryocytes and on canine platelet function. Stem Cells 1994, 12, 386–393. [Google Scholar] [CrossRef]
- Baatout, S. Interleukin-6 and megakaryocytopoiesis: An update. Ann. Hematol. 1996, 73, 157–162. [Google Scholar] [CrossRef]
- Beaulieu, L.M.; Lin, E.; Mick, E.; Koupenova, M.; Weinberg, E.O.; Kramer, C.D.; Genco, C.A.; Tanriverdi, K.; Larson, M.G.; Benjamin, E.J.; et al. Interleukin 1 receptor 1 and interleukin 1β regulate megakaryocyte maturation, platelet activation, and transcript profile during inflammation in mice and humans. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 552–564. [Google Scholar] [CrossRef]
- Orkin, S.H.; Shivdasani, R.A.; Fujiwara, Y.; McDevitt, M.A. Transcription factor GATA-1 in megakaryocyte development. Stem Cells 1998, 16 (Suppl. 2), 79–83. [Google Scholar]
- Muntean, A.G.; Pang, L.; Poncz, M.; Dowdy, S.F.; Blobel, G.A.; Crispino, J.D. Cyclin D-Cdk4 is regulated by GATA-1 and required for megakaryocyte growth and polyploidization. Blood 2007, 109, 5199–5207. [Google Scholar] [CrossRef]
- Chagraoui, H.; Kassouf, M.; Banerjee, S.; Goardon, N.; Clark, K.; Atzberger, A.; Pearce, A.C.; Skoda, R.C.; Ferguson, D.J.; Watson, S.P.; et al. SCL-mediated regulation of the cell-cycle regulator p21 is critical for murine megakaryopoiesis. Blood 2011, 118, 723–735. [Google Scholar] [CrossRef]
- Cullmann, K.; Jahn, M.; Spindler, M.; Schenk, F.; Manukjan, G.; Mucci, A.; Steinemann, D.; Boller, K.; Schulze, H.; Bender, M.; et al. Forming megakaryocytes from murine-induced pluripotent stem cells by the inducible overexpression of supporting factors. Res. Pract. Thromb. Haemost. 2021, 5, 111–124. [Google Scholar] [CrossRef]
- DiMartino, J.F.; Selleri, L.; Traver, D.; Firpo, M.T.; Rhee, J.; Warnke, R.; O’Gorman, S.; Weissman, I.L.; Cleary, M.L. The Hox cofactor and proto-oncogene Pbx1 is required for maintenance of definitive hematopoiesis in the fetal liver. Blood 2001, 98, 618–626. [Google Scholar] [CrossRef]
- Tang, C.; Shi, X.; Wang, W.; Zhou, D.; Tu, J.; Xie, X.; Ge, Q.; Xiao, P.F.; Sun, X.; Lu, Z. Global analysis of in vivo EGR1-binding sites in erythroleukemia cell using chromatin immunoprecipitation and massively parallel sequencing. Electrophoresis 2010, 31, 2936–2943. [Google Scholar] [CrossRef]
- Yen, J.H.; Lin, C.Y.; Chuang, C.H.; Chin, H.K.; Wu, M.J.; Chen, P.Y. Nobiletin Promotes Megakaryocytic Differentiation through the MAPK/ERK-Dependent EGR1 Expression and Exerts Anti-Leukemic Effects in Human Chronic Myeloid Leukemia (CML) K562 Cells. Cells 2020, 9, 877. [Google Scholar] [CrossRef]
- Gao, Q.; Jeon, S.J.; Jung, H.A.; Lee, H.E.; Park, S.J.; Lee, Y.; Lee, Y.; Ko, S.Y.; Kim, B.; Choi, J.S.; et al. Nodakenin Enhances Cognitive Function and Adult Hippocampal Neurogenesis in Mice. Neurochem. Res. 2015, 40, 1438–1447. [Google Scholar] [CrossRef]
- Lim, W.; Yang, C.; Bazer, F.W.; Song, G. Chrysophanol Induces Apoptosis of Choriocarcinoma Through Regulation of ROS and the AKT and ERK1/2 Pathways. J. Cell. Physiol. 2017, 232, 331–339. [Google Scholar] [CrossRef]
- Hsieh, M.H.; Tsai, J.P.; Yang, S.F.; Chiou, H.L.; Lin, C.L.; Hsieh, Y.H.; Chang, H.R. Fisetin Suppresses the Proliferation and Metastasis of Renal Cell Carcinoma through Upregulation of MEK/ERK-Targeting CTSS and ADAM9. Cells 2019, 8, 948. [Google Scholar] [CrossRef]
- Zhang, S.; Xue, R.; Geng, Y.; Wang, H.; Li, W. Fisetin Prevents HT22 Cells From High Glucose-Induced Neurotoxicity via PI3K/Akt/CREB Signaling Pathway. Front. Neurosci. 2020, 14, 241. [Google Scholar] [CrossRef]
- Bae, H.J.; Kim, J.; Kim, J.; Goo, N.; Cai, M.; Cho, K.; Jung, S.Y.; Kwon, H.; Kim, D.H.; Jang, D.S.; et al. The effect of maslinic acid on cognitive dysfunction induced by cholinergic blockade in mice. Br. J. Pharmacol. 2020, 177, 3197–3209. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Schriml, L.M.; Arze, C.; Nadendla, S.; Chang, Y.W.; Mazaitis, M.; Felix, V.; Feng, G.; Kibbe, W.A. Disease Ontology: A backbone for disease semantic integration. Nucleic Acids Res. 2012, 40, D940–D946. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Wang, L.; Zeng, J.; Wu, A.; Qin, M.; Wen, M.; Zhang, T.; Chen, W.; Mei, Q.; Qin, D.; et al. Caulis Polygoni Multiflori Accelerates Megakaryopoiesis and Thrombopoiesis via Activating PI3K/Akt and MEK/ERK Signaling Pathways. Pharmaceuticals 2022, 15, 1204. https://doi.org/10.3390/ph15101204
Yang X, Wang L, Zeng J, Wu A, Qin M, Wen M, Zhang T, Chen W, Mei Q, Qin D, et al. Caulis Polygoni Multiflori Accelerates Megakaryopoiesis and Thrombopoiesis via Activating PI3K/Akt and MEK/ERK Signaling Pathways. Pharmaceuticals. 2022; 15(10):1204. https://doi.org/10.3390/ph15101204
Chicago/Turabian StyleYang, Xin, Long Wang, Jing Zeng, Anguo Wu, Mi Qin, Min Wen, Ting Zhang, Wang Chen, Qibing Mei, Dalian Qin, and et al. 2022. "Caulis Polygoni Multiflori Accelerates Megakaryopoiesis and Thrombopoiesis via Activating PI3K/Akt and MEK/ERK Signaling Pathways" Pharmaceuticals 15, no. 10: 1204. https://doi.org/10.3390/ph15101204