
Evaluation of Gallic Acid-Coated Gold Nanoparticles as an Anti-Aging Ingredient


Abstract
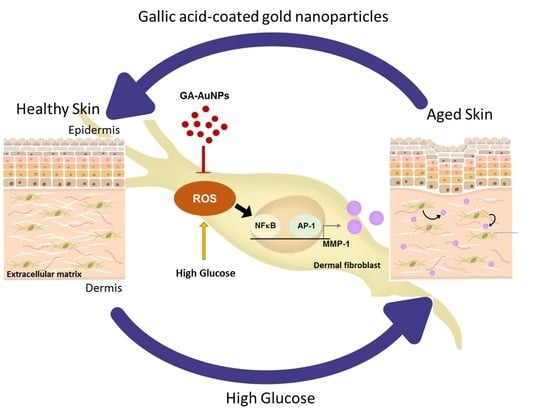
:
1. Introduction
2. Results
2.1. Characterization of Synthesized GA–AuNPs
2.2. GA–AuNPs Prevents High Glucose-Induced MMP-1 Expression in Human Dermal Fibroblasts
2.3. GA–AuNPs Inhibited MMP-1 Expression in Glucose-Induced Fibroblast Cells through ASK1, MAPKs, NFκB and AP-1 Pathways
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Synthesis of GA–AuNPs
4.3. AuNPs Size and Zeta Potential Measurements
4.4. Preparation of Stock Solutions of GA-AuNPs
4.5. Cell Culture
4.6. Cell Viability Assay
4.7. Evaluation of the Intracellular Reactive Oxygen Species (ROS) Production
4.8. Real-Time Polymerase Chain Reaction
4.9. Western Blot Analysis
4.10. Collagenase Activity—Zymography Assay
4.11. Collagen Zymography
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tobin, D.J. Introduction to skin aging. J. Tissue Viability 2017, 26, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noordam, R.; Gunn, D.A.; Tomlin, C.C.; Maier, A.B.; Mooijaart, S.P.; Slagboom, P.E.; Westendorp, R.G.J.; de Craen, A.J.M.; van Heemst, D. High serum glucose levels are associated with a higher perceived age. AGE 2013, 35, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Makrantonaki, E.; Jiang, D.; Hossini, A.M.; Nikolakis, G.; Wlaschek, M.; Scharffetter-Kochanek, K.; Zouboulis, C.C. Diabetes mellitus and the skin. Rev. Endocr. Metab. Disord. 2016, 17, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Kruse, C.R.; Singh, M.; Sørensen, J.A.; Eriksson, E.; Nuutila, K. The effect of local hyperglycemia on skin cells in vitro and on wound healing in euglycemic rats. J. Surg. Res. 2016, 206, 418–426. [Google Scholar] [CrossRef] [Green Version]
- Rolo, A.P.; Palmeira, C.M. Diabetes and mitochondrial function: Role of hyperglycemia and oxidative stress. Toxicol. Appl. Pharmacol. 2006, 212, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Vanessa Fiorentino, T.; Prioletta, A.; Zuo, P.; Folli, F. Hyperglycemia-induced oxidative stress and its role in diabetes mellitus related cardiovascular diseases. Curr. Pharm. Des. 2013, 19, 5695–5703. [Google Scholar] [CrossRef]
- Matsumoto, N.; Omagari, D.; Ushikoshi-Nakayama, R.; Yamazaki, T.; Inoue, H.; Saito, I. Hyperglycemia induces generation of reactive oxygen species and accelerates apoptotic cell death in salivary gland cells. Pathobiology 2021, 88, 234–241. [Google Scholar] [CrossRef]
- Senthil, K.K.; Gokila, V.M.; Mau, J.L.; Lin, C.C.; Chu, F.H.; Wei, C.C.; Liao, V.H.C.; Wang, S.Y. A steroid like phytochemical Antcin M is an anti-aging reagent that eliminates hyperglycemia-accelerated premature senescence in dermal fibroblasts by direct activation of Nrf2 and SIRT-1. Oncotarget 2016, 7, 62836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, S.; Hopley, B.; Genovese, L.; Sibilla, S.; Laight, D.; Shute, J. Effects of collagen-derived bioactive peptides and natural antioxidant compounds on proliferation and matrix protein synthesis by cultured normal human dermal fibroblasts. Sci. Rep. 2018, 8, 10474. [Google Scholar] [CrossRef]
- Poljsak, B. Strategies for reducing or preventing the generation of oxidative stress. Oxid. Med. Cell. Longev. 2011, 2011, 194586. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wang, X.; Du, J.; Gu, Z.; Zhao, Y. Reactive oxygen species-regulating strategies based on nanomaterials for disease treatment. Adv. Sci. 2021, 8, 2002797. [Google Scholar] [CrossRef] [PubMed]
- Badhani, B.; Sharma, N.; Kakkar, R. Gallic acid: A versatile antioxidant with promising therapeutic and industrial applications. RSC Adv. 2015, 5, 27540–27557. [Google Scholar] [CrossRef]
- Chen, J.; Yang, J.; Ma, L.; Li, J.; Shahzad, N.; Kim, C.K. Structure-antioxidant activity relationship of methoxy, phenolic hydroxyl, and carboxylic acid groups of phenolic acids. Sci. Rep. 2020, 10, 2611. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Alam, M.B.; Baek, M.E.; Kwon, Y.G.; Lim, J.Y.; Lee, S.H. Protection against UVB-induced photoaging by Nypa fruticans via inhibition of MAPK/AP-1/MMP-1 signaling. Oxid. Med. Cell. Longev. 2020, 2020, 2905362. [Google Scholar] [CrossRef]
- Moldovan, M.; Lahmar, A.; Bogdan, C.; Părăuan, S.; Tomuţă, I.; Crişan, M. Formulation and evaluation of a water-in-oil cream containing herbal active ingredients and ferulic acid. Clujul Med. 2017, 90, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, S.; Cho, S.H.; Park, D.; Jung, E. Anti-skin aging properties of proto-catechuic acid in vitro and in vivo. J. Cosmet. Dermatol. 2020, 19, 977–984. [Google Scholar] [CrossRef]
- Mahato, K.; Nagpal, S.; Shah, M.A.; Srivastava, A.; Maurya, P.K.; Roy, S.; Jaiswal, A.; Singh, R.; Chandra, P. Gold nanoparticle surface engineering strategies and their applications in biomedicine and diagnostics. 3 Biotech 2019, 9, 57. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, Y.; Ding, T.; Liu, J.; Zhao, H. Multifunctional gold nanoparticles: A novel nanomaterial for various medical applications and biological activities. Front. Bioeng. Biotechnol. 2020, 8, 990. [Google Scholar] [CrossRef]
- Amini, S.M.; Akbari, A. Metal nanoparticles synthesis through natural phenolic acids. IET Nanobiotechnol. 2019, 13, 771–777. [Google Scholar] [CrossRef]
- Punithavathi, V.R.; Prince, P.S.M.; Kumar, R.; Selvakumari, J. Antihyperglycaemic, antilipid peroxidative and antioxidant effects of gallic acid on streptozotocin induced diabetic Wistar rats. Eur. J. Pharmacol. 2011, 650, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.H.S.; Yen, J.H.; Wu, H.T.; Huang, S.T. Gallic acid inhibited matrix invasion and AP-1/ETS-1-mediated MMP-1 transcription in human nasopharyngeal carcinoma cells. Int. J. Mol. Sci. 2017, 18, 1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, E.; Park, S.Y.; Lee, H.J.; Lee, T.Y.; Sun, Z.W.; Yi, T.H. Gallic acid regulates skin photoaging in UVB-exposed fibroblast and hairless mice. Phytother. Res. 2014, 28, 1778–1788. [Google Scholar] [CrossRef]
- Chen, Y.J.; Lee, Y.C.; Huang, C.H.; Chang, L.S. Gallic acid-capped gold nanoparticles inhibit EGF-induced MMP-9 expression through suppression of p300 stabilization and NFκB/c-Jun activation in breast cancer MDA-MB-231 cells. Toxicol. Appl. Pharmacol. 2016, 310, 98–107. [Google Scholar] [CrossRef]
- Patungwasa, W.; Hodak, J.H. pH tunable morphology of the gold nanoparticles produced by citrate reduction. Mater. Chem. Phys. 2008, 108, 45–54. [Google Scholar] [CrossRef]
- Kaasalainen, M.; Aseyev, V.; von Haartman, E.; Karaman, D.Ş.; Mäkilä, E.; Tenhu, H.; Rosenholm, J.; Salonen, J. Size, stability, and porosity of mesoporous nanoparticles characterized with light scattering. Nanoscale Res. Lett. 2017, 12, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; El-Sayed, M.A. Gold nanoparticles: Optical properties and implementations in cancer diagnosis and photothermal therapy. J. Adv. Res. 2021, 1, 13–28. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.P.; Ma, B.Y.; Wei, X.W.; Qian, Z.Y. The in vitro and in vivo toxicity of gold nanoparticles. Chin. Chem. Lett. 2017, 28, 691–702. [Google Scholar] [CrossRef]
- Cao, C.; Xiao, Z.; Wu, Y.; Ge, C. Diet and skin aging—From the perspective of food nutrition. Nutrients 2020, 12, 870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, L.; Dixon, C.L.; Aguilera-Aguirre, L.; Menon, R. Oxidative stress-induced TGF-beta/TAB1-mediated p38 MAPK activation in human amnion epithelial cells. Biol. Reprod. 2018, 99, 1100–1112. [Google Scholar] [CrossRef]
- Jung, Y.R.; Kim, D.H.; Kim, S.R.; An, H.J.; Lee, E.K.; Tanaka, T.; Kim, N.D.; Yokozawa, T.; Park, J.N.; Chung, H.Y. Anti-wrinkle effect of magnesium lithospermate B from Salvia miltiorrhiza bunge: Inhibition of MMPs via NF-κB signaling. PLoS ONE 2014, 9, e102689. [Google Scholar] [CrossRef] [Green Version]
- Wiraguna, A.A.G.P.; Dianasari, R.; Pangkahila, W. The topical skin application of purple corn extract (Zea mays) inhibited the increase in MMP-1 levels and decreased collagen in wistar rats (Rattus norvegicus) exposed to UV-B Rays. Biomed. Pharmacol. J. 2019, 35, 297–304. [Google Scholar] [CrossRef]
- Lu, Y.E.; Chen, Y.J. Resveratrol inhibits matrix metalloproteinase-1 and-3 expression by suppressing of p300/NFκB acetylation in TNF-α-treated human dermal fibroblasts. Chem. Biol. Interact. 2021, 337, 109395. [Google Scholar] [CrossRef]
- Jiménez, Z.; Kim, Y.J.; Mathiyalagan, R.; Seo, K.H.; Mohanan, P.; Ahn, J.C.; Kim, Y.J.; Yang, D.C. Assessment of radical scavenging, whitening and moisture retention activities of Panax ginseng berry mediated gold nanoparticles as safe and efficient novel cosmetic material. Artif. Cells Nanomed. Biotechnol. 2018, 46, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, N.; Muddapur, U. Biosynthesis of metal nanoparticles: A review. J. Nanotechnol. 2014, 2014, 510246. [Google Scholar] [CrossRef] [Green Version]
- Teodoro, A.J. Bioactive compounds of food: Their role in the prevention and treatment of diseases. Oxid. Med. Cell. 2019, 2019, 3765986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, J.; Wang, Y. MDI 301, a synthetic retinoid, depressed levels of matrix metalloproteinases and oxidative stress in diabetic dermal fibroblasts. Oncotarget 2017, 8, 43889. [Google Scholar] [CrossRef] [Green Version]
- Argyropoulos, A.J.; Robichaud, P.; Balimunkwe, R.M.; Fisher, G.J.; Hammerberg, C.; Yan, Y.; Quan, T. Alterations of dermal connective tissue collagen in diabetes: Molecular basis of aged-appearing skin. PLoS ONE 2016, 11, e0153806. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Jackson, C.J. Extracellular matrix reorganization during wound healing and its impact on abnormal scarring. Adv. Wound Caref. 2015, 4, 119–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, W.; Wang, Y.S.; Hwang, E.; Lin, P.; Bae, J.; Seo, S.A.; Yi, T.H. Rubus idaeus L. (red raspberry) blocks UVB-induced MMP production and promotes type I pro-collagen synthesis via inhibition of MAPK/AP-1, NF-κB and stimulation of TGF-β/Smad, Nrf2 in normal human dermal fibroblasts. J. Photochem. Photobiol. B Biol. 2018, 185, 241–253. [Google Scholar] [CrossRef]
- Riedlinger, T.; Liefke, R.; Meier-Soelch, J.; Jurida, L.; Nist, A.; Stiewe, T.; Lienhard Schmitz, M. NF-κB p65 dimerization and DNA-binding is important for inflam-matory gene expression. FASEB J. 2019, 33, 4188–4202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, B.; Baldwin, A.S., Jr.; Ballard, D.W.; Greene, W.C.; Angel, P.; Herrlich, P. Cross-coupling of the NF-kappa B p65 and Fos/Jun transcription factors produces potentiated biological function. EMBO J. 1993, 12, 3879–3891. [Google Scholar] [CrossRef]
- Chen, Y.J.; Chang, L.S. NFκB-and AP-1-mediated DNA looping regulates matrix metalloproteinase-9 transcription in TNF-α-treated human leukemia U937 cells. Biochim. Biophys. Acta Gene Regul. Mech. 2015, 1849, 1248–1259. [Google Scholar] [CrossRef]
- Chen, Y.J.; Lin, K.N.; Jhang, L.M.; Huang, C.H.; Lee, Y.C.; Chang, L.S. Gallic acid abolishes the EGFR/Src/Akt/Erk-mediated expression of matrix metalloprotein-ase-9 in MCF-7 breast cancer cells. Chem. Biol. Interact. 2016, 252, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Raju, G.; Katiyar, N.; Vadukumpully, S.; Shankarappa, S.A. Penetration of gold nanoparticles across the stratum corneum layer of thick-skin. J. Dermatol. Sci. 2018, 89, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Román, R.; Naik, A.; Kalia, Y.N.; Guy, R.H.; Fessi, H. Skin penetration and distribution of polymeric nanoparticles. J. Control. Release 2004, 99, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, V.; Mitragotri, S. Nanoparticles for topical drug delivery: Potential for skin cancer. Adv. Drug Deliv. Rev. 2020, 153, 87–108. [Google Scholar] [CrossRef]
- Vogt, A.; Combadiere, B.; Hadam, S.; Stieler, K.M.; Lademann, J.; Schaefer, H.; Autran, B.; Sterry, W.; Blume-Peytavi, U. 40 nm, but not 750 or 1500 nm, nanoparticles enter epidermal CD1a+ cells after transcutaneous application on human skin. J. Invest. Dermatol. 2006, 126, 1316–1322. [Google Scholar] [CrossRef] [Green Version]
- Koeppl, S.; Solenthaler, C.; Caseri, W.; Spolenak, R. Towards a reproducible synthesis of high aspect ratio gold nanorods. J. Nanomater. 2011, 2011, 515049. [Google Scholar] [CrossRef]
- Eslami, A.C.; Pasanphan, W.; Wagner, B.A.; Buettner, G.R. Free radicals produced by the oxidation of gallic acid: An electron paramagnetic resonance study. Chem. Cent. J. 2010, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Zuorro, A.; Iannone, A.; Natali, S.; Lavecchia, R. Green synthesis of silver nanoparticles using bilberry and red currant waste extracts. Processes 2019, 7, 193. [Google Scholar] [CrossRef] [Green Version]
- Suchaoin, W.; de Sousa, I.P.; Netsomboon, K.; Lam, H.T.; Laffleur, F.; Bern-kop-Schnürch, A. Development and in vitro evaluation of zeta potential changing self-emulsifying drug delivery systems for enhanced mucus permeation. Int. J. Pharm. 2016, 510, 255–262. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Median Size (nm) | Mean Size (nm) ± SD | Particle Numbers |
|---|---|---|---|
| 6 | 36.43 | 36.02 ± 5.80 | n = 129 |
| 9 | 26.02 | 26.23 ± 4.16 | n = 172 |
| 11 | 29.88 | 30.30 ± 3.98 | n = 119 |
| pH | Mean Size (nm) ± SD | Zeta Potential (mV) ± SD | Polydispersity Index |
|---|---|---|---|
| 6 | 52.0 ± 18.1 | 61.30 ± 2.84 | 0.259 |
| 9 | 32 ± 20.8 | 48.26 ± 1.84 | 0.311 |
| 11 | 40.8 ± 15.4 | –49.63 ± 2.11 | 0.141 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.-Z.; Tsai, Y.-Y.; Chang, L.-S.; Chen, Y.-J. Evaluation of Gallic Acid-Coated Gold Nanoparticles as an Anti-Aging Ingredient. Pharmaceuticals 2021, 14, 1071. https://doi.org/10.3390/ph14111071
Wu Y-Z, Tsai Y-Y, Chang L-S, Chen Y-J. Evaluation of Gallic Acid-Coated Gold Nanoparticles as an Anti-Aging Ingredient. Pharmaceuticals. 2021; 14(11):1071. https://doi.org/10.3390/ph14111071
Chicago/Turabian StyleWu, Yun-Zhen, Yen-Yu Tsai, Long-Sen Chang, and Ying-Jung Chen. 2021. "Evaluation of Gallic Acid-Coated Gold Nanoparticles as an Anti-Aging Ingredient" Pharmaceuticals 14, no. 11: 1071. https://doi.org/10.3390/ph14111071