
Optimising Multispectral Active Fluorescence to Distinguish the Photosynthetic Variability of Cyanobacteria and Algae
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
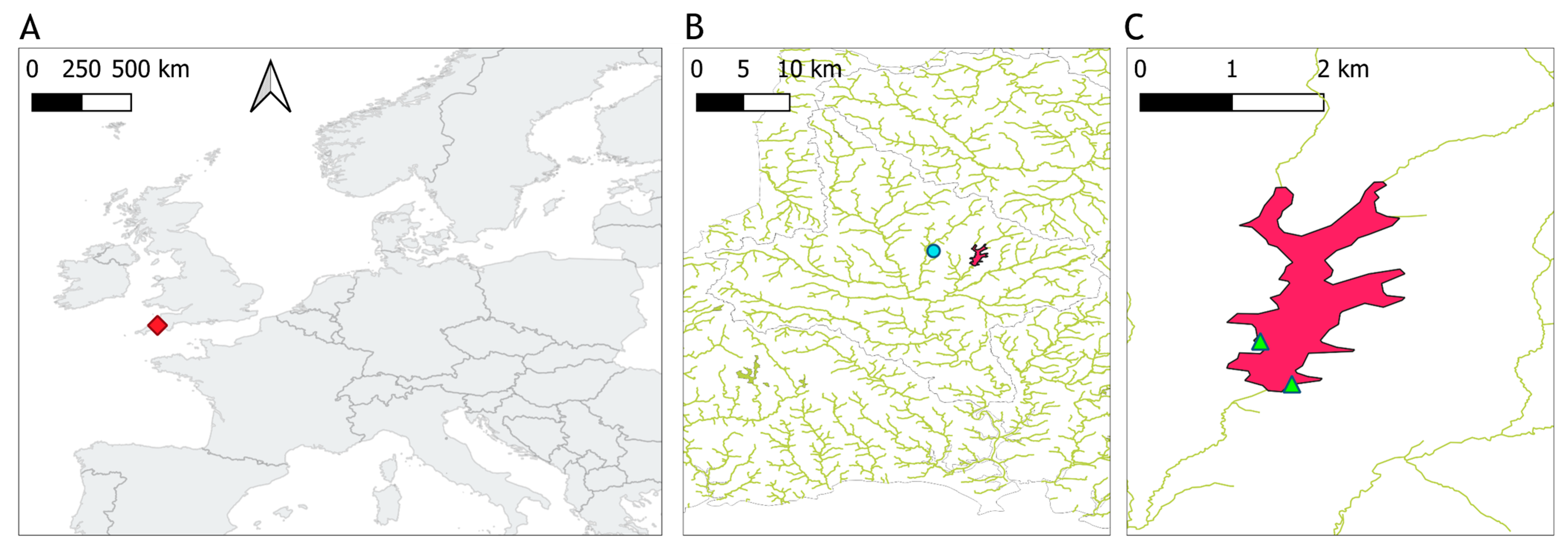
2.1. Study Site
2.2. Single-Turnover Fluorescence
2.3. Microscopy
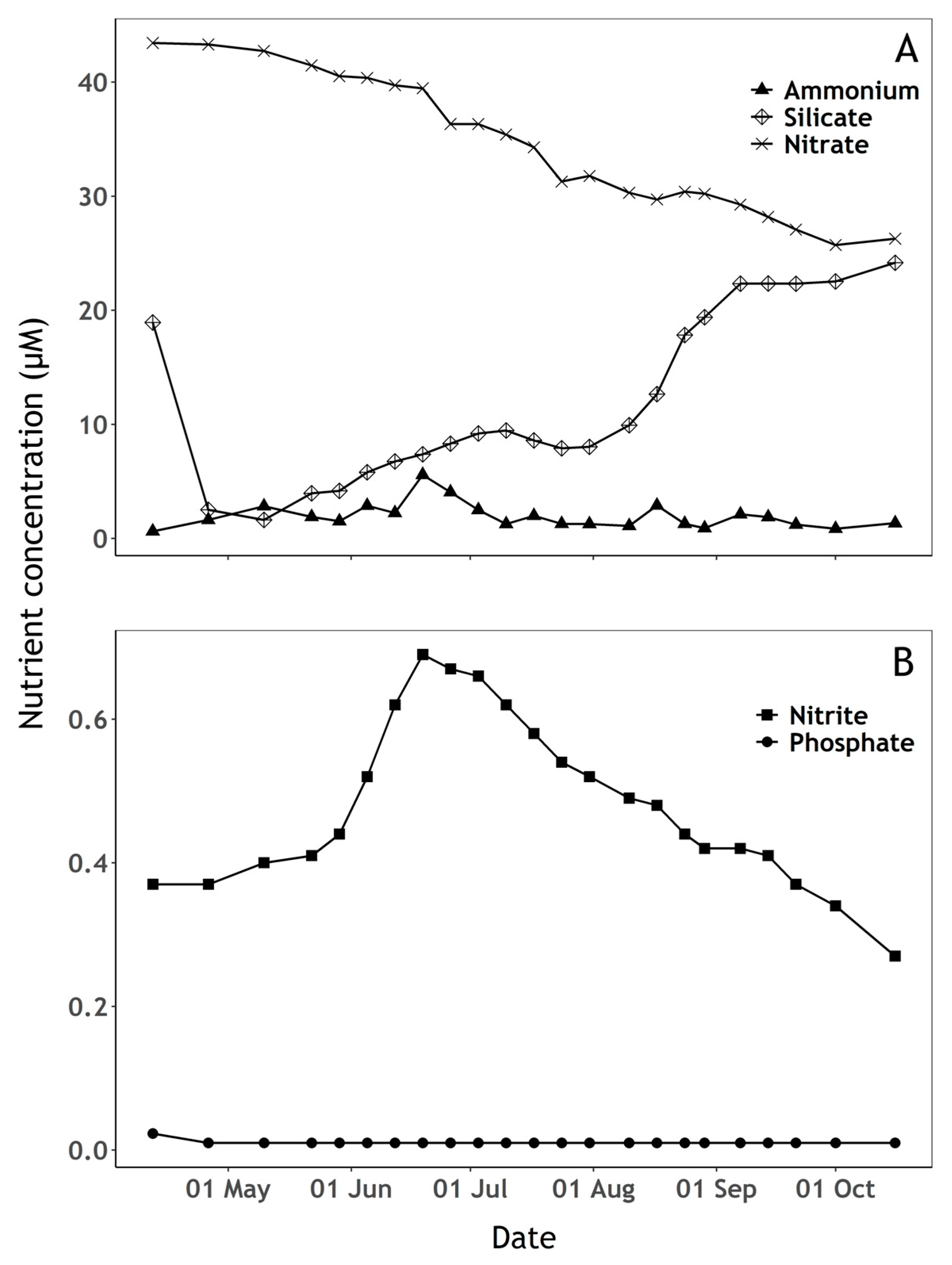
2.4. Nutrients
3. Results
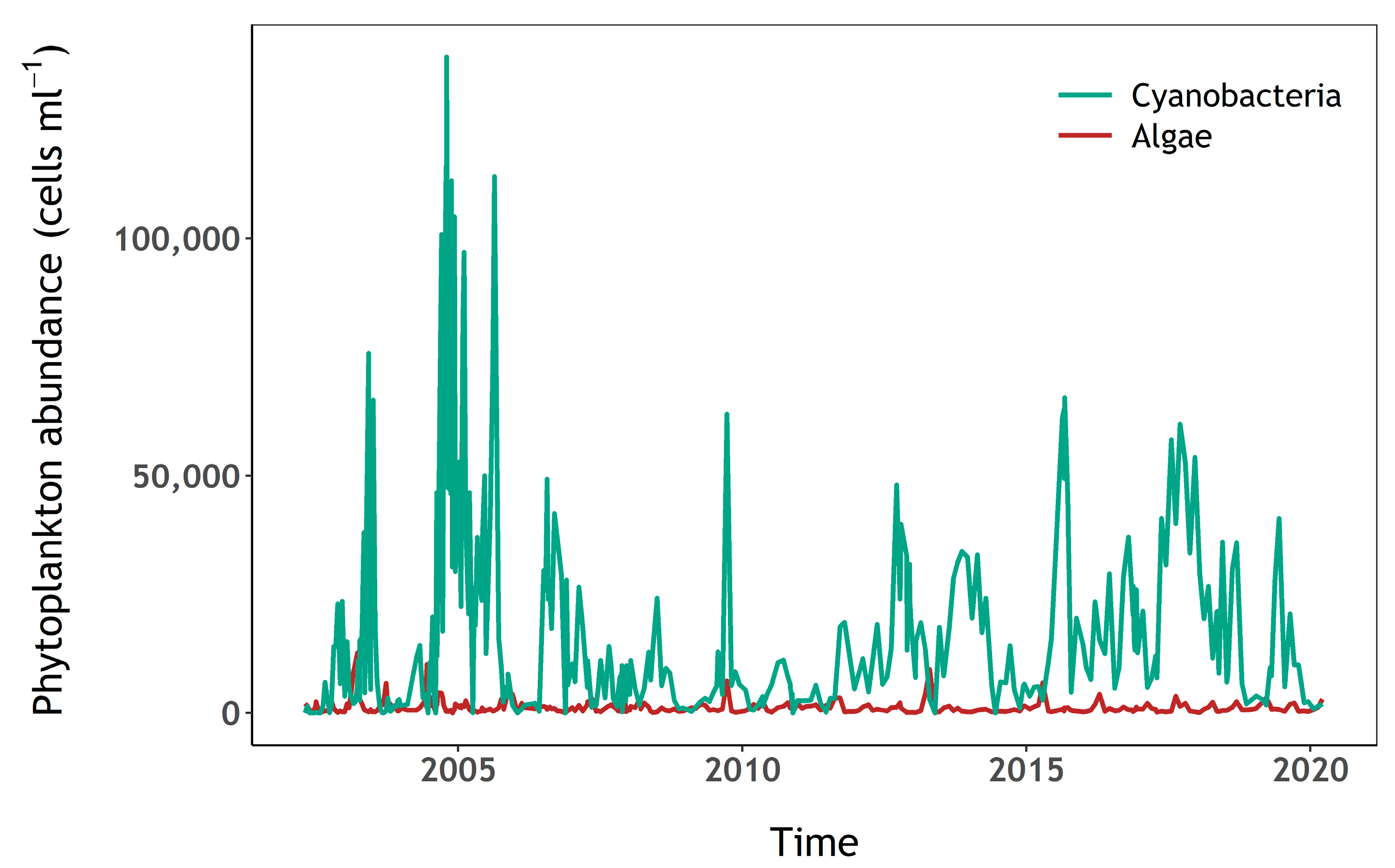
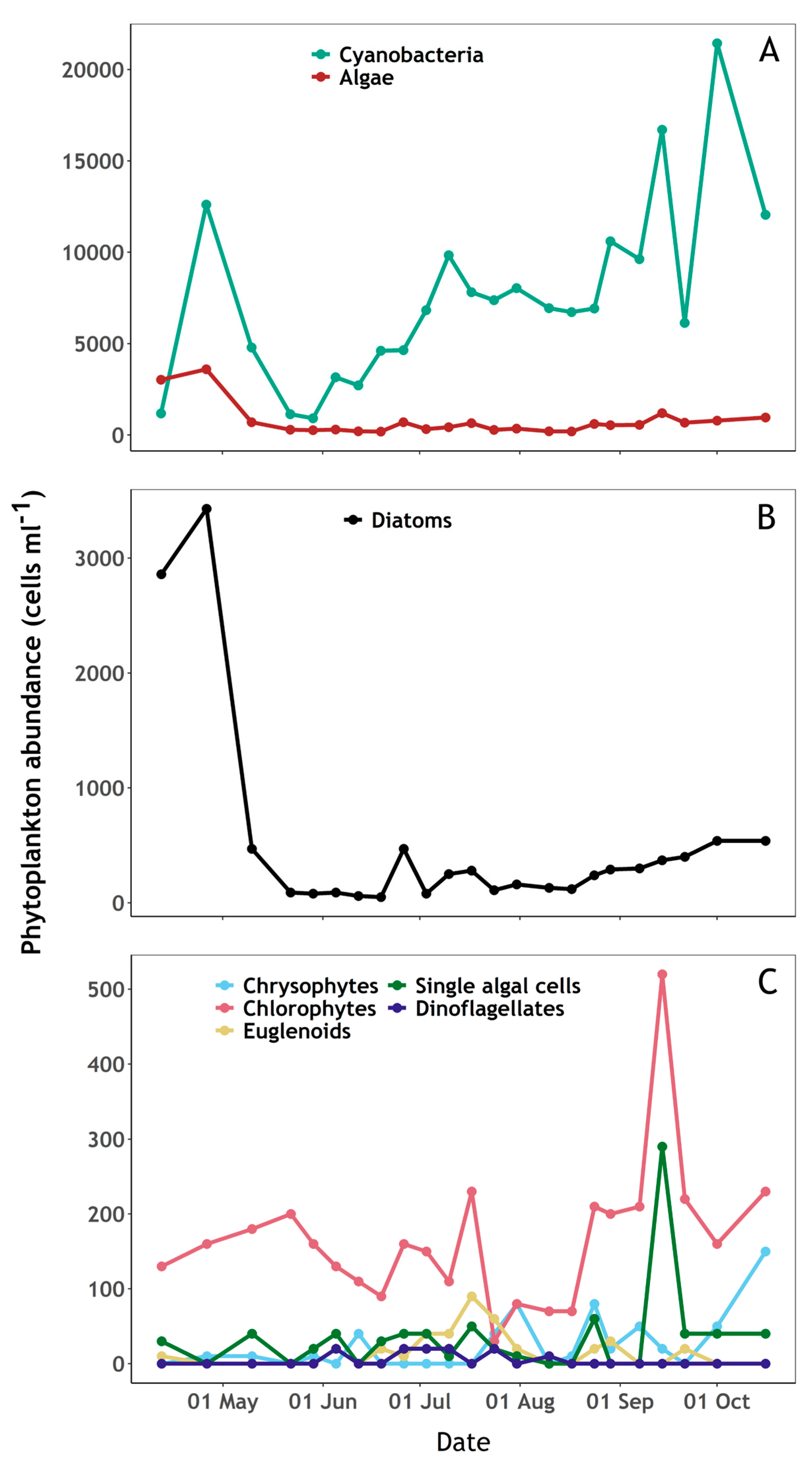
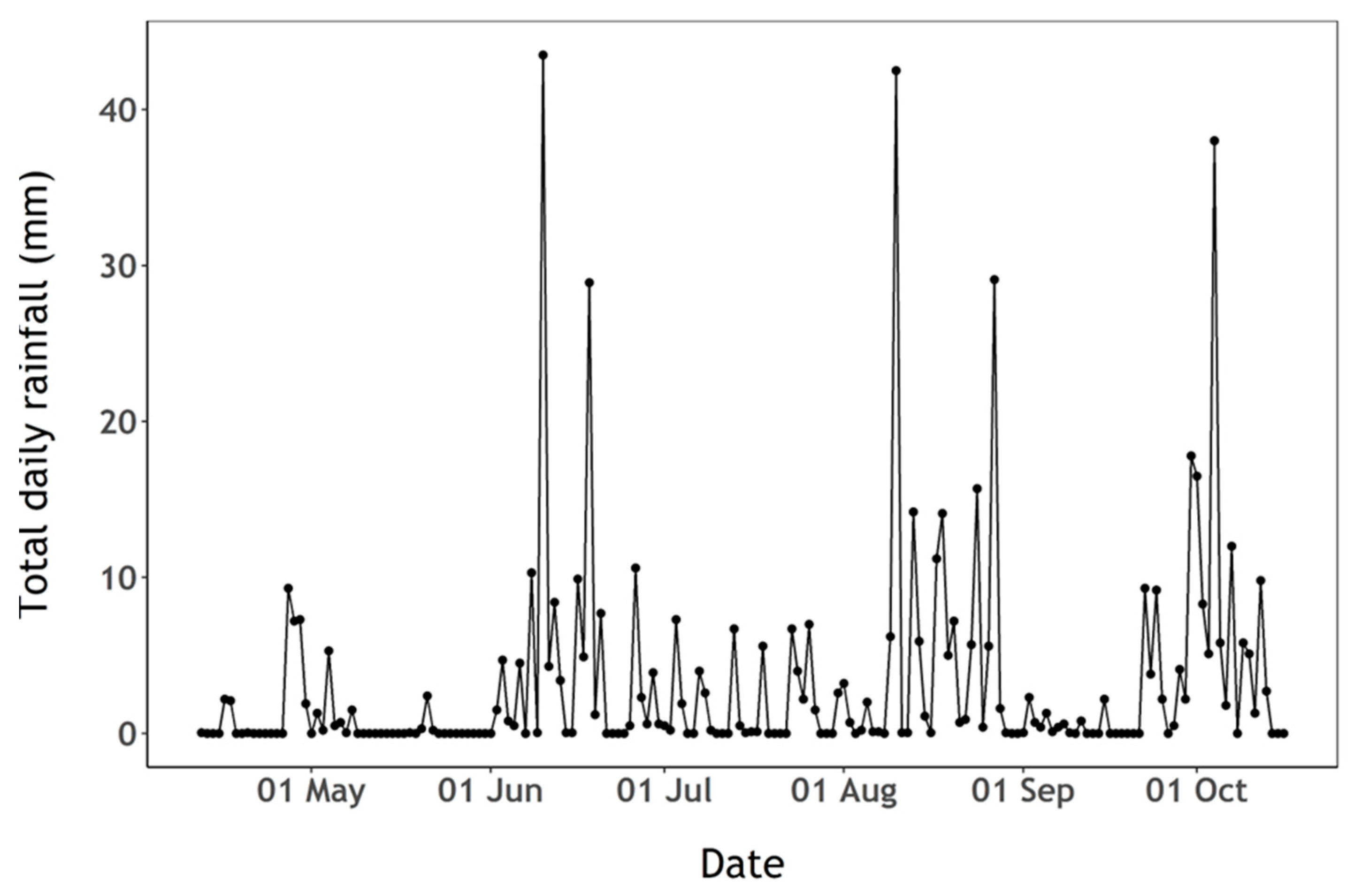
3.1. Phytoplankton and Nutrient Dynamics
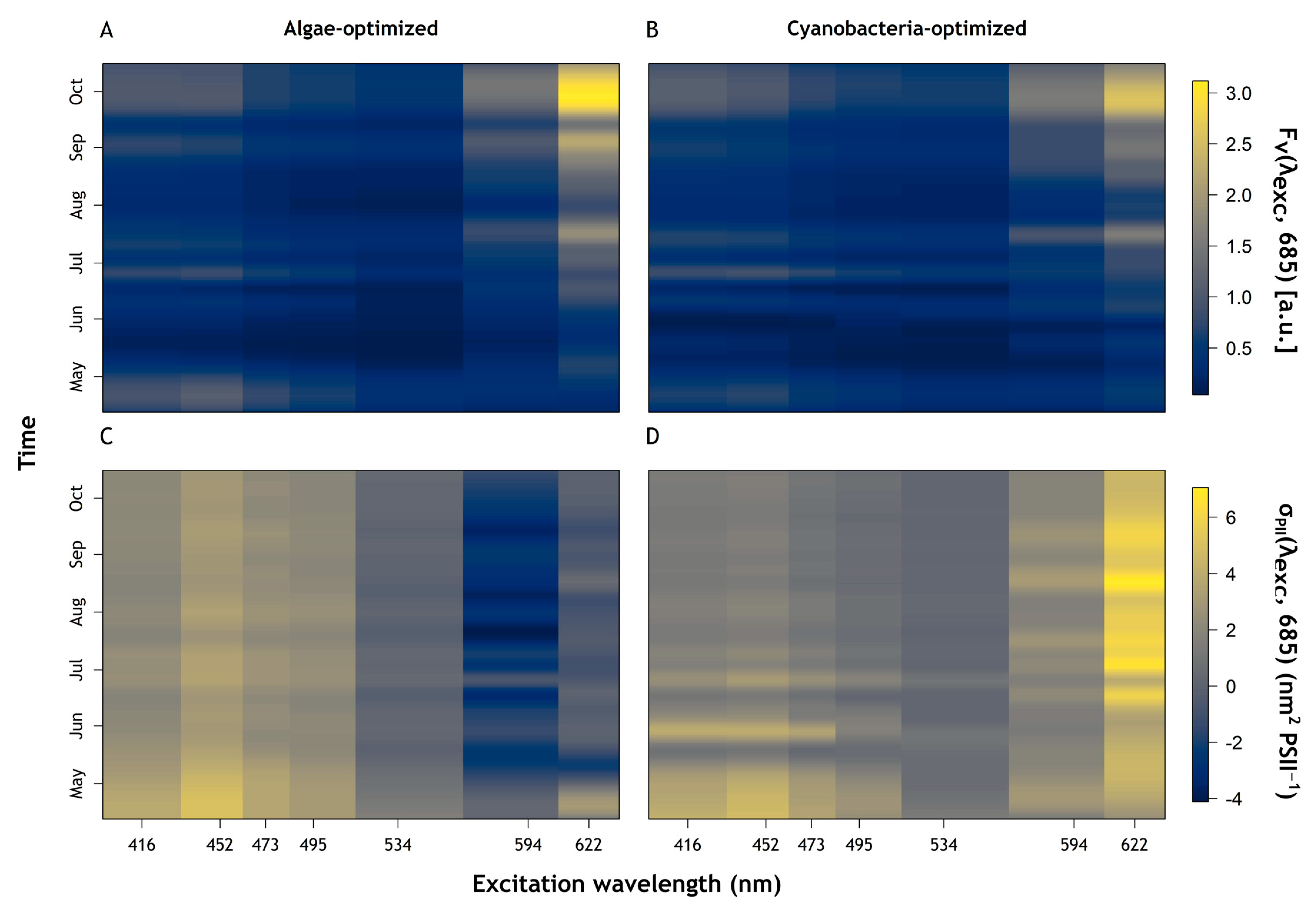
3.2. Fluorescence Dynamics
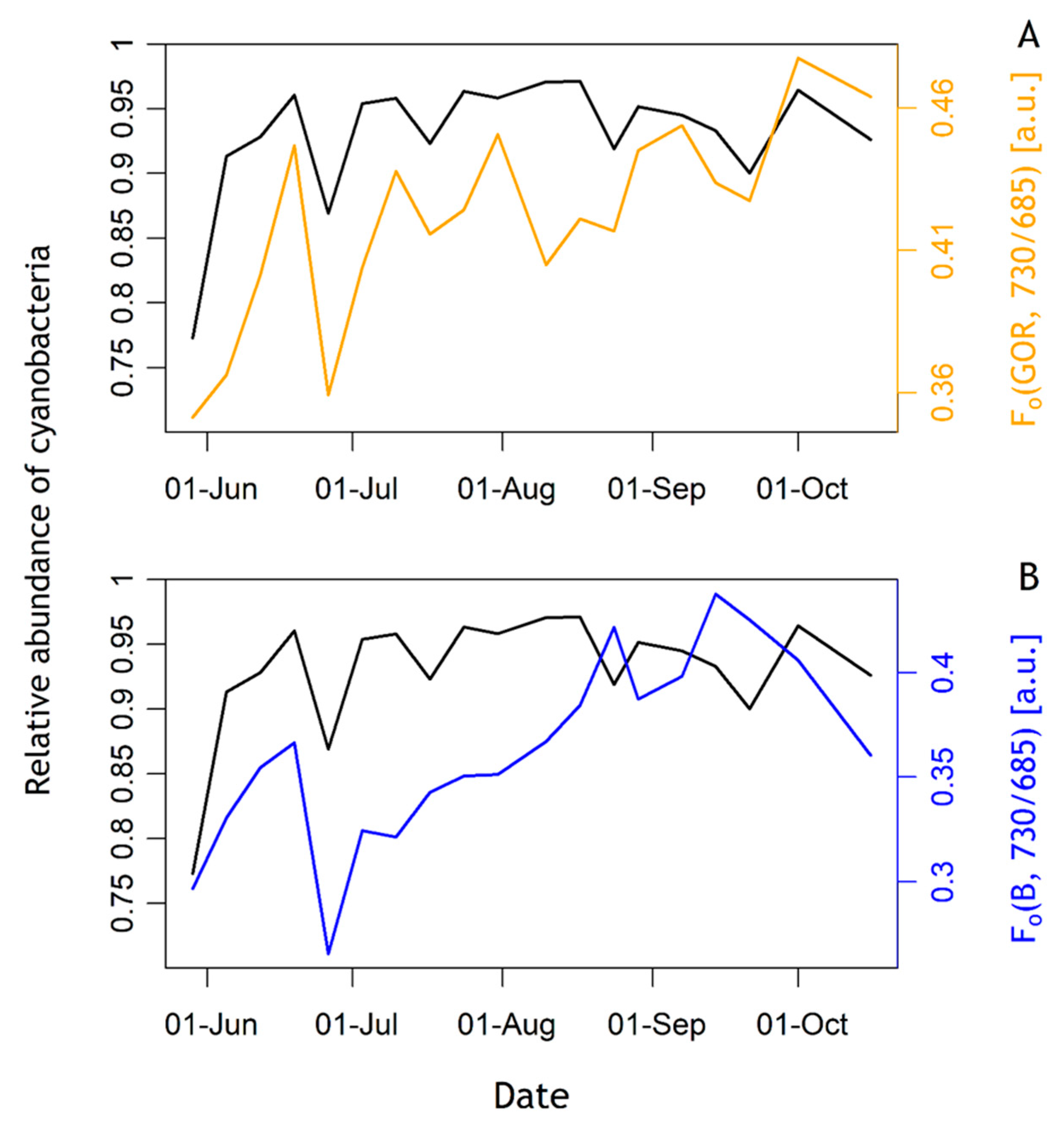
3.3. Relative Cyanobacteria Abundance and Fo Dynamics
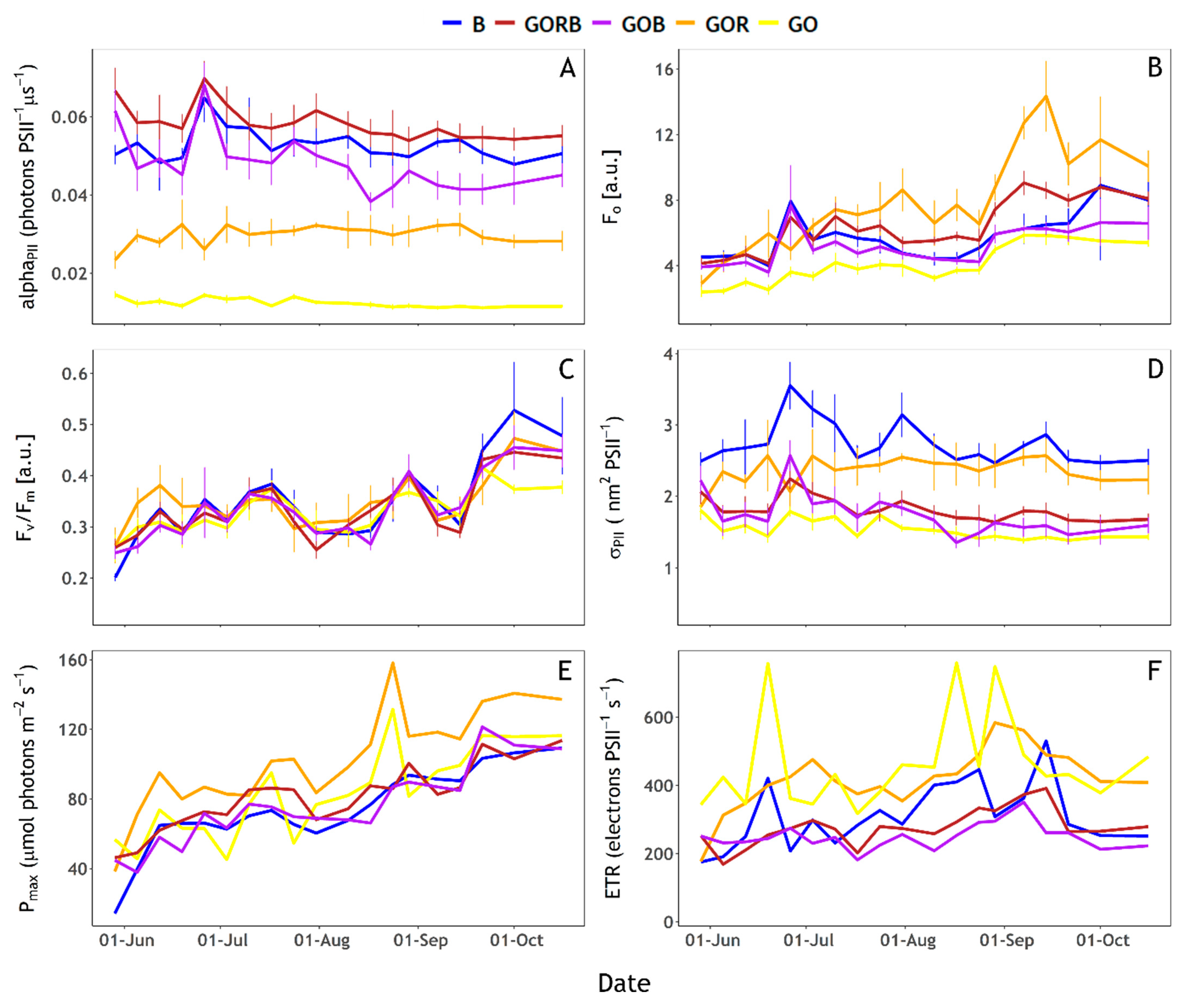
3.4. Photophysiological Characterisation
3.5. Emission Ratio of 730 nm over 685 nm
4. Discussion
4.1. Phytoplankton, Nutrient and Fluorescence Dynamics
4.2. Emission Ratio of 730 nm over 685 nm
4.3. Photophysiological Characterisation
4.4. Suitability of LabSTAF for In Situ Assessment of Cyanobacteria
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Blooms Like It Hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paerl; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Hall, N.S.; Calandrino, E.S. Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic-induced change. Sci. Total Environ. 2011, 409, 1739–1745. [Google Scholar] [CrossRef]
- Paerl, H.W.; Paul, V.J. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2012, 46, 1349–1363. [Google Scholar] [CrossRef]
- Wurtsbaugh, W.A.; Paerl, H.W.; Dodds, W.K. Nutrients, eutrophication and harmful algal blooms along the freshwater to marine continuum. Wiley Interdiscip. Rev. Water 2019, 6, e1373. [Google Scholar] [CrossRef]
- Codd, G.A.; Lindsay, J.; Young, F.M.; Morrison, L.F.; Metcalf, J.S. Harmful Cyanobacteria: From mass mortalities to management measures. In Harmful Cyanobacteria; Springer: Dordrecht, The Netherlands; Berlin, Germany, 2005; pp. 1–23. [Google Scholar]
- Codd, G.A.; Morrison, L.F.; Metcalf, J.S. Cyanobacterial toxins: Risk management for health protection. Toxicol. Appl. Pharmacol. 2005, 203, 264–272. [Google Scholar] [CrossRef]
- Whitton, B.A.; Potts, M. The Ecology of Cyanobacteria: Their Diversity in Time and Space; Kluwer Acad: Dordrecht, The Netherlands, 2000. [Google Scholar]
- Carmichael, W.W.; An, J. Using an enzyme linked immunosorbent assay (ELISA) and a protein phosphatase inhibition assay (PPIA) for the detection of microcystins and nodularins. Nat. Toxins 1999, 7, 377–385. [Google Scholar] [CrossRef]
- Baker, J.A.; Entsch, B.; Neilan, B.A.; McKay, D.B. Monitoring changing toxigenicity of a cyanobacterial bloom by molecular methods. Appl. Environ. Microbiol. 2002, 68, 6070–6076. [Google Scholar] [CrossRef] [Green Version]
- Oehrle, S.A.; Southwell, B.; Westrick, J. Detection of various freshwater cyanobacterial toxins using ultra-performance liquid chromatography tandem mass spectrometry. Toxicon 2010, 55, 965–972. [Google Scholar] [CrossRef]
- Pearson, L.A.; Neilan, B.A. The molecular genetics of cyanobacterial toxicity as a basis for monitoring water quality and public health risk. Curr. Opin. Biotechnol. 2008, 19, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Singh, S.; Ahn, C.-Y.; Oh, H.-M.; Asthana, R.K. Monitoring approaches for a toxic cyanobacterial bloom. Environ. Sci. Technol. 2013, 47, 8999–9013. [Google Scholar] [CrossRef]
- Beutler, M.; Wiltshire, K.H.; Meyer, B.; Moldaenke, C.; Lüring, C.; Meyerhöfer, M.; Hansen, U.-P.; Dau, H. A fluorometric method for the differentiation of algal populations in vivo and in situ. Photosynth. Res. 2002, 72, 39–53. [Google Scholar] [CrossRef]
- Becker, A.; Meister, A.; Wilhelm, C. Flow cytometric discrimination of various phycobilin-containing phytoplankton groups in a hypertrophic reservoir. Cytometry Suppl. 2002, 48, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.; Henrichs, D.W.; Olson, R.J.; Sosik, H.M. Continuous automated imaging-in-flow cytometry for detection and early warning of Karenia brevis blooms in the Gulf of Mexico. Environ. Sci. Pollut. Res. 2013, 20, 6896–6902. [Google Scholar] [CrossRef]
- Huot, Y.; Babin, M. Overview of Fluorescence Protocols: Theory, Basic Concepts, and Practice. In Chlorophyll a Fluorescence in Aquatic Sciences: Methods and Applications; Suggett, D.J., Prášil, O., Borowitzka, M.A., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 31–74. [Google Scholar]
- Suggett, D.J.; Moore, C.M.; Oxborough, K.; Geider, R.J. Fast Repetition Rate (FRR) Chlorophyll a Fluorescence Induction Measurements; Chelsea Technologies Group, Ltd.: West Molesey, UK, 2006; 53p, Available online: https://data.imas.utas.edu.au/attachments/fc64fc7b-29b0-4607-9527-899dfa991d68/FRRFmethodsManual.pdf (accessed on 23 December 2022).
- Babin, M. Phytoplankton fluorescence: Theory, current literature and in situ measurement. In Real-Time Coastal Observing Systems for Marine Ecosystem Dynamics and Harmful Algal Blooms; UNESCO: Paris, France, 2008; pp. 237–280. [Google Scholar]
- Suggett, D.J.; Oxborough, K.; Baker, N.R.; MacIntyre, H.L.; Kana, T.M.; Geider, R.J. Fast repetition rate and pulse amplitude modulation chlorophyll a fluorescence measurements for assessment of photosynthetic electron transport in marine phytoplankton. Eur. J. Phycol. 2003, 38, 371–384. [Google Scholar] [CrossRef] [Green Version]
- Houliez, E.; Simis, S.; Nenonen, S.; Ylöstalo, P.; Seppälä, J. Basin-scale spatio-temporal variability and control of phytoplankton photosynthesis in the Baltic Sea: The first multiwavelength fast repetition rate fluorescence study operated on a ship-of-opportunity. J. Mar. Syst. 2017, 169, 40–51. [Google Scholar] [CrossRef]
- Kazama, T.; Hayakawa, K.; Kuwahara, V.S.; Shimotori, K.; Imai, A.; Komatsu, K. Development of photosynthetic carbon fixation model using multi-excitation wavelength fast repetition rate fluorometry in Lake Biwa. PLoS ONE 2021, 16, e0238013. [Google Scholar] [CrossRef]
- Gorbunov, M.Y.; Shirsin, E.; Nikonova, E.; Fadeev, V.V.; Falkowski, P.G. A multi-spectral fluorescence induction and relaxation (FIRe) technique for physiological and taxonomic analysis of phytoplankton communities. Mar. Ecol. Prog. Ser. 2020, 644, 1–13. [Google Scholar] [CrossRef]
- Schreiber, U.; Klughammer, C.; Kolbowski, J. Assessment of wavelength-dependent parameters of photosynthetic electron transport with a new type of multi-color PAM chlorophyll fluorometer. Photosynth. Res. 2012, 113, 127–144. [Google Scholar] [CrossRef]
- Schuback, N.; Tortell, P.D.; Berman-Frank, I.; Campbell, D.A.; Ciotti, A.; Courtecuisse, E.; Erickson, Z.K.; Fujiki, T.; Halsey, K.; Hickman, A.E. Single-turnover variable chlorophyll fluorescence as a tool for assessing phytoplankton photosynthesis and primary productivity: Opportunities, caveats and recommendations. Front. Mar. Sci. 2021, 8, 690607. [Google Scholar] [CrossRef]
- Gregor, J.; Maršálek, B. A simple in vivo fluorescence method for the selective detection and quantification of freshwater cyanobacteria and eukaryotic algae. Acta Hydrochim. Hydrobiol. 2005, 33, 142–148. [Google Scholar] [CrossRef]
- Gregor, J.; Maršálek, B.; Šípková, H. Detection and estimation of potentially toxic cyanobacteria in raw water at the drinking water treatment plant by in vivo fluorescence method. Water Res. 2007, 41, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Seppälä, J.; Ylöstalo, P.; Kaitala, S.; Hällfors, S.; Raateoja, M.; Maunula, P. Ship-of-opportunity based phycocyanin fluorescence monitoring of the filamentous cyanobacteria bloom dynamics in the Baltic Sea. Estuar. Coast. Shelf Sci. 2007, 73, 489–500. [Google Scholar] [CrossRef]
- Dodds, W. Freshwater Ecology: Concepts and Environmental Applications; Academic Press: San Diego, CA, USA, 2002. [Google Scholar]
- Dittami, S.M.; Heesch, S.; Olsen, J.L.; Collén, J. Transitions between marine and freshwater environments provide new clues about the origins of multicellular plants and algae. J. Phycol. 2017, 53, 731–745. [Google Scholar] [CrossRef] [Green Version]
- Price, D.C.; Steiner, J.M.; Yoon, H.S.; Bhattacharya, D.; Löffelhardt, W. Glaucophyta. In Handbook of the Protists, 2nd ed.; Springer: Cham, Switzerland, 2017; pp. 23–87. [Google Scholar] [CrossRef]
- Mimuro, M.; Fujita, Y. Estimation of chlorophyll a distribution in the photosynthetic pigment systems I and II of the blue-green alga Anabaena variabilis. Biochim. Biophys. Acta-Bioenerg. 1977, 459, 376–389. [Google Scholar] [CrossRef]
- Johnsen, G.; Sakshaug, E. Biooptical characteristics of PSII and PSI in 33 species (13 pigment groups) of marine phytoplankton, and the relevance for pulse-amplitude-modulated and fast-repetition-rate fluorometry. J. Phycol. 2007, 43, 1236–1251. [Google Scholar] [CrossRef]
- Courtecuisse, E.; Oxborough, K.; Tilstone, G.H.; Spyrakos, E.; Hunter, P.D.; Simis, S.G.H. Determination of optical markers of cyanobacterial physiology from fluorescence kinetics. J. Plankton Res. 2022, 44, 365–385. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Raven, J.A. Aquatic Photosynthesis, 2nd ed.; Princeton University Press: Princeton, NJ, USA, 2013. [Google Scholar]
- Oxborough, K. LabSTAF and RunSTAF Handbook: 2408-014-HB | Issue F; Chelsea Technologies, Ltd.: West Molesey, UK, 2022; 171p. [Google Scholar] [CrossRef]
- Boatman, T.G.; Geider, R.J.; Oxborough, K. Improving the accuracy of single turnover active fluorometry (STAF) for the estimation of phytoplankton primary productivity (PhytoPP). Front. Mar. Sci. 2019, 6, 319. [Google Scholar] [CrossRef] [Green Version]
- Lawson, J.; Sambrook, H.; Solomon, D.; Weilding, G. The Roadford scheme: Minimizing environmental impact on affected catchments. Water Environ. J. 1991, 5, 671–681. [Google Scholar] [CrossRef]
- Bryant, D.A.; Guglielmi, G.; de Marsac, N.T.; Castets, A.-M.; Cohen-Bazire, G. The structure of cyanobacterial phycobilisomes: A model. Arch. Microbiol. 1979, 123, 113–127. [Google Scholar] [CrossRef]
- Johnsen, G.; Sakshaug, E. Light harvesting in bloom-forming marine phytoplankton: Species specificity and photoacclimation. Sci. Mar. 1996, 60, 47–56. [Google Scholar]
- Silsbe, G.M.; Oxborough, K.; Suggett, D.J.; Forster, R.M.; Ihnken, S.; Komárek, O.; Lawrenz, E.; Prášil, O.; Röttgers, R.; Šicner, M. Toward autonomous measurements of photosynthetic electron transport rates: An evaluation of active fluorescence-based measurements of photochemistry. Limnol. Oceanogr. Methods 2015, 13, 138–155. [Google Scholar] [CrossRef] [Green Version]
- Woodward, E.; Rees, A. Nutrient distributions in an anticyclonic eddy in the northeast Atlantic Ocean, with reference to nanomolar ammonium concentrations. Deep Sea Res. Part II: Top. Stud. Oceanogr. 2001, 48, 775–793. [Google Scholar] [CrossRef]
- Wetzel, R.G. Limnology: Lake and River Ecosystems, 3rd ed.; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- de Figueiredo, D.R.; Reboleira, A.S.; Antunes, S.C.; Abrantes, N.; Azeiteiro, U.; Goncalves, F.; Pereira, M.J. The effect of environmental parameters and cyanobacterial blooms on phytoplankton dynamics of a Portuguese temperate lake. Hydrobiologia 2006, 568, 145–157. [Google Scholar] [CrossRef]
- Carey, C.C.; Ibelings, B.W.; Hoffmann, E.P.; Hamilton, D.P.; Brookes, J.D. Eco-physiological adaptations that favour freshwater cyanobacteria in a changing climate. Water Res. 2012, 46, 1394–1407. [Google Scholar] [CrossRef]
- Yoshida, M.; Horiuchi, T.; Nagasawa, Y. Situ Multi-Excitation Chlorophyll Fluorometer for Phytoplankton Measurements: Technologies and Applications beyond Conventional Fluorometers; Oceans’11 MTS/IEEE KONA; IEEE: Piscataway, NJ, USA, 2011; pp. 1–4. [Google Scholar]
- Calzadilla, P.I.; Kirilovsky, D. Revisiting cyanobacterial state transitions. Photochem. Photobiol. Sci. 2020, 19, 585–603. [Google Scholar] [CrossRef] [Green Version]
- Luimstra, V.M.; Schuurmans, J.M.; Verschoor, A.M.; Hellingwerf, K.J.; Huisman, J.; Matthijs, H.C. Blue light reduces photosynthetic efficiency of cyanobacteria through an imbalance between photosystems I and II. Photosynth. Res. 2018, 138, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Suggett, D.J.; Moore, C.M.; Hickman, A.E.; Geider, R.J. Interpretation of fast repetition rate (FRR) fluorescence: Signatures of phytoplankton community structure versus physiological state. Mar. Ecol. Prog. Ser. 2009, 376, 1–19. [Google Scholar] [CrossRef]
- Lavergne, J.; Joliot, P. Thermodynamics of the excited states of photosynthesis. In Biophysics Textbook Online; Biophysical Society: Bethesda, MD, USA, 2000; pp. 1–12. [Google Scholar]
- Parésys, G.; Rigart, C.; Rousseau, B.; Wong, A.; Fan, F.; Barbier, J.-P.; Lavaud, J. Quantitative and qualitative evaluation of phytoplankton communities by trichromatic chlorophyll fluorescence excitation with special focus on cyanobacteria. Water Res. 2005, 39, 911–921. [Google Scholar] [CrossRef] [Green Version]
- Raateoja, M.; Seppälä, J.; Ylöstalo, P. Fast repetition rate fluorometry is not applicable to studies of filamentous cyanobacteria from the Baltic Sea. Limnol. Oceanogr. 2004, 49, 1006–1012. [Google Scholar] [CrossRef]
- Suggett, D.J.; MacIntyre, H.L.; Kana, T.M.; Geider, R.J. Comparing electron transport with gas exchange: Parameterising exchange rates between alternative photosynthetic currencies for eukaryotic phytoplankton. Aquat. Microb. Ecol. 2009, 56, 147–162. [Google Scholar] [CrossRef] [Green Version]
- Napoléon, C.; Claquin, P. Multi-parametric relationships between PAM measurements and carbon incorporation, an in situ approach. PLoS ONE 2012, 7, e40284. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protocol | Excitation Wavebands (nm at Centre) | Intensity Range (μmol Photons m−2 s−1 ) | Pulse Length (μs) | Photosynthetic Pigment Groups Targeted | Phytoplankton Group Targeted |
|---|---|---|---|---|---|
| B | 452, 452 | 28,175–33,538 | 100 | Chlorophylls a/b/c, carotenoids | Algae, weak signal from cyanobacteria possible |
| GOR | 534, 594, 622 | 20,990 | 200 | Phycobilipigments Chlorophylls a/b/c and carotenoids | Cyanobacteria with weaker signal from algae likely |
| GORB | 452, 452, 534, 594, 622 | 49,513–54,528 | 200 | Chlorophylls a/b/c, carotenoids and phycobilipigments | Whole community |
| GOB | 452, 452, 534, 594 | 41,919–46,933 | 200 | Chlorophylls a/b/c, carotenoids and phycobilipigments | Whole community with a slower saturation response compared to GORB |
| GO | 534, 594 | 13,395 | 200 | Phycobilipigments, Chlorophyll c, carotenoids | Cyanobacteria, cryptophytes and rhodophytes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Courtecuisse, E.; Marchetti, E.; Oxborough, K.; Hunter, P.D.; Spyrakos, E.; Tilstone, G.H.; Simis, S.G.H. Optimising Multispectral Active Fluorescence to Distinguish the Photosynthetic Variability of Cyanobacteria and Algae. Sensors 2023, 23, 461. https://doi.org/10.3390/s23010461
Courtecuisse E, Marchetti E, Oxborough K, Hunter PD, Spyrakos E, Tilstone GH, Simis SGH. Optimising Multispectral Active Fluorescence to Distinguish the Photosynthetic Variability of Cyanobacteria and Algae. Sensors. 2023; 23(1):461. https://doi.org/10.3390/s23010461
Chicago/Turabian StyleCourtecuisse, Emilie, Elias Marchetti, Kevin Oxborough, Peter D. Hunter, Evangelos Spyrakos, Gavin H. Tilstone, and Stefan G. H. Simis. 2023. "Optimising Multispectral Active Fluorescence to Distinguish the Photosynthetic Variability of Cyanobacteria and Algae" Sensors 23, no. 1: 461. https://doi.org/10.3390/s23010461