1. Introduction
Delayed systems play an important role in chemical kinetics [
1,
2]. Even for the complex network of the first order reactions, it can be described by relatively simple system of delayed differential equations, in which the effects of intermediates are replaced by time lags [
3,
4].
The interaction between two chemicals A and B, forming the product C, is not instant but is during some time interval
. Hence, the law of mass action as the fundamental law of chemical kinetics should be reformulated schematically as:
leading us to the differential equation with delay:
which is called the law of delayed mass action.
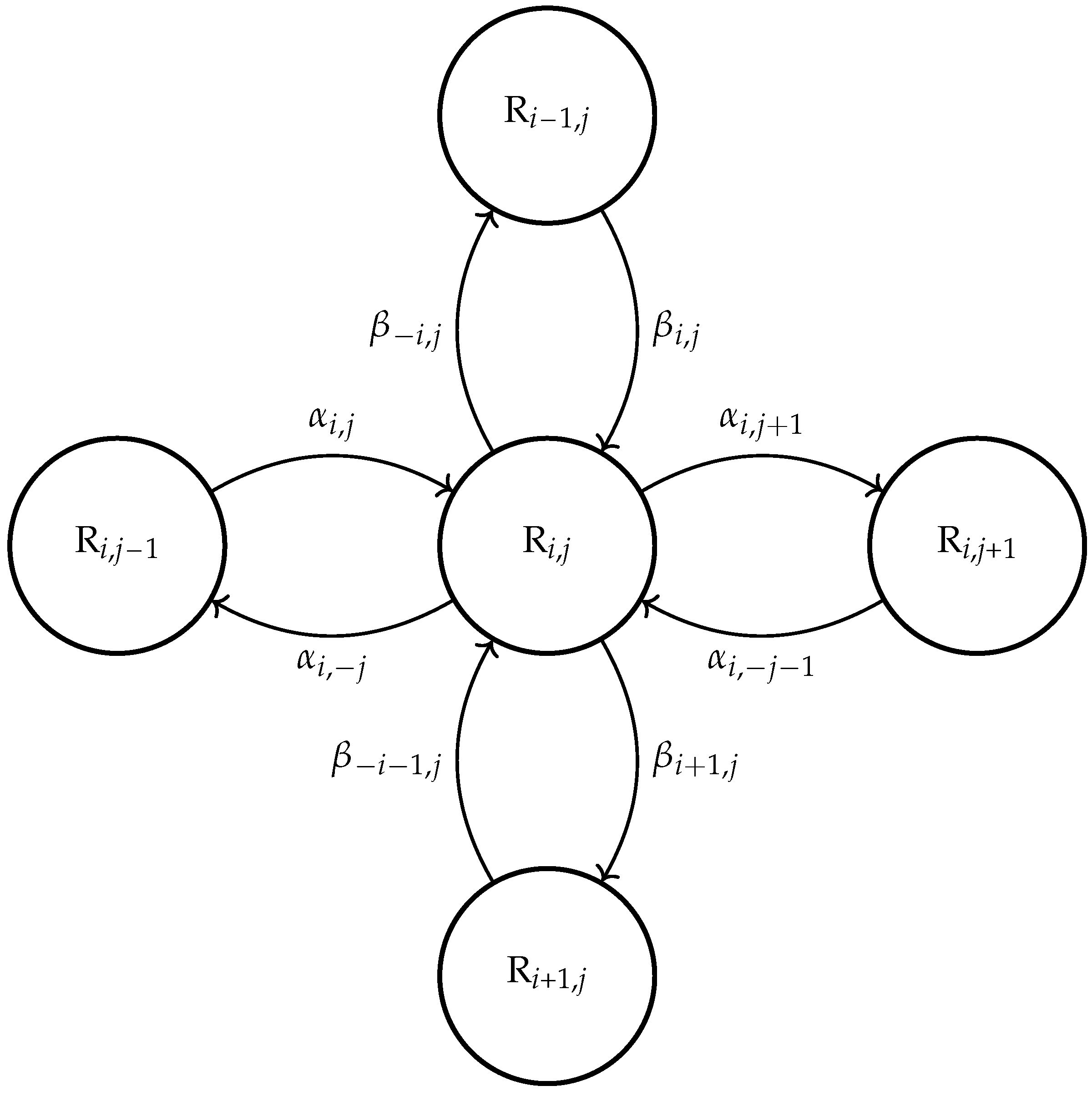
Since delay
is rather a random variable than a deterministic one and can accept various values according to some distribution laws, here, we offer a considerably more advanced model which includes continuously distributed delays (or simply, distributed delays), which can be described by the differential equation with distributed delay:
where
is the probability density function of time delay
. The model (
2) can be referred to as the law of mass action with distributed delay. In practice, we can consider an integral with limited bounds in (
2), using the properties of the distribution of random variables such as Chebyshev’s inequality, as it will be shown in
Section 3.1.
The reactions which are used in electrochemical biosensing come from the reactions that are catalyzed by an enzyme. They are commonly known as reversible [
5] or irreversible [
6] reactions. The irreversible one-complex Michaelis–Menthen (IR1CMM) mechanism is a keystone in modeling enzyme kinetics. Its reaction scheme
represents a two-step process [
7,
8,
9], where the enzyme E combines with the substrate S to form a complex C, which then breaks down into the product P, releasing E in the process. The mechanism IR1CMM is described with the help of ordinary differential equations:
Here, for any substance A, we denote its concentration at instant t as .
An application of the delayed mass action law to enzyme kinetics was inspired by Brown’s model, formulated in [
10], where complex C has a lifetime
before being decayed. We call the following reaction scheme:
the irreversible one-complex Brown’s (IR1CB) mechanism, which can be described with the following system of delayed differential equations:
When comparing mechanisms IR1CMM and IR1CB, Roussel, M.R has introduced the notion of a chemically acceptable model [
1]. While evidencing that the model using delayed mass action law is more adequate, the most significant failure of the lag model was that the solutions of these equations oscillate around the equilibrium point, which is forbidden by the law of microscopic reversibility. Oscillatory enzyme reactions are found in a number of enzymatic systems. Goldbeter, A. investigated the influence of Michaelis–Menten kinetics on the oscillatory behavior in an enzyme system [
11].
Albornoz, J.M. and Parravano, A. have also shown that the models based on delayed differential equations (DDE) oscillate at small values of the Michaelis–Menthen constant, which cannot be seen with the help of ordinary differential equations (ODE) [
12]. Piephoff, D.E. et al. has described the conformation of non-equilibrium enzyme kinetics, where a traditional Michaelis–Menten model is extended to a generalized form, which includes corrections coming from informational currents within combined cyclic kinetics loops [
13].
Hinch, R. and Schnell, S. studied the conditions of equivalence of enzyme–substrate reaction mechanisms involving multiple complexes with a distributed delay system without complexes [
14]. It was shown that the distribution of the delay is determined by the number of intermediate complexes and the rates of the individual reaction mechanisms.
In continuing the research [
14] and applying the mass action law with the distributed delay (
2), we offer the following continuously distributed delay model:
where
is density function of delay distribution. In the manuscript [
15], theoretical knowledge about kinetics of single molecule associated with the Michaelis–Menten model has been given.
This study was motivated by the desire to describe all possible complexes created during enzyme–substrate interaction with the help of the delay density function. Moreover, we offer an effective method for its parameter estimation in one special case.
4. Experimental Study
4.1. Enzyme–Substrate Interaction
Enzyme biosensors consist of enzymes immobilized at the surface of the transducer. Hence, an immobilization step with the help of Bovine Serum Albumin (BSA) is very important as it affects the sensitivity and selectivity of biosensors. Enzyme products may be electroactive, meaning their activity may be followed by amperometry.
In our study, Acetylcholinesterase (AChE) as an enzyme has been used, since it has a very high catalytic activity. AChE is often used to design biosensors based on the inhibition analysis.
Since conductivity (specifically, electrolytic conductivity) is the ability of a substance to conduct an electric current, in solvents where electrical conductivity is present, particularly water, ionisation will provide the necessary carriers. The electrical conductivity of a solution is determined by both the physical characteristics of the carriers and the medium. The measured conductivity comes from the ions of dissolved substances. The preliminary measurement has been conducted in aqueous solutions.
The studies were carried out in aqueous solutions using the following substances: protein—BSA; enzyme—enzyme acetylcholinesterase (AChE); and substrate—Acetylcholine chloride (AChCl). The substances were purchased from Sigma Aldrich.
To the complex formed with BSA of 2 mg/mL (volume of 4 mL) and enzyme of 2 mg/mL (volume of 0.1 mL) (the complex is known as cross-linked enzyme aggregate (CLEA) [
22]), substrate of 2 mg/mL in different volumes (0.1 mL, 0.3 mL, 0.9 mL, 1.5 mL) was added. As a result, four samples were obtained:
Sample 1. BSA + enzyme + 0.1 mL of substrate;
Sample 2. BSA + enzyme + 0.3 mL of substrate;
Sample 3. BSA + enzyme + 0.9 mL of substrate;
Sample 4. BSA + enzyme + 1.5 mL of substrate.
The conductivity of the BSA + enzyme + substrate complex was tested by using a specially constructed measuring setup, during 500 s with signal sampling every 5 s. There were 100 conductivity measurements made during 500 s.
4.1.1. Experimental Data
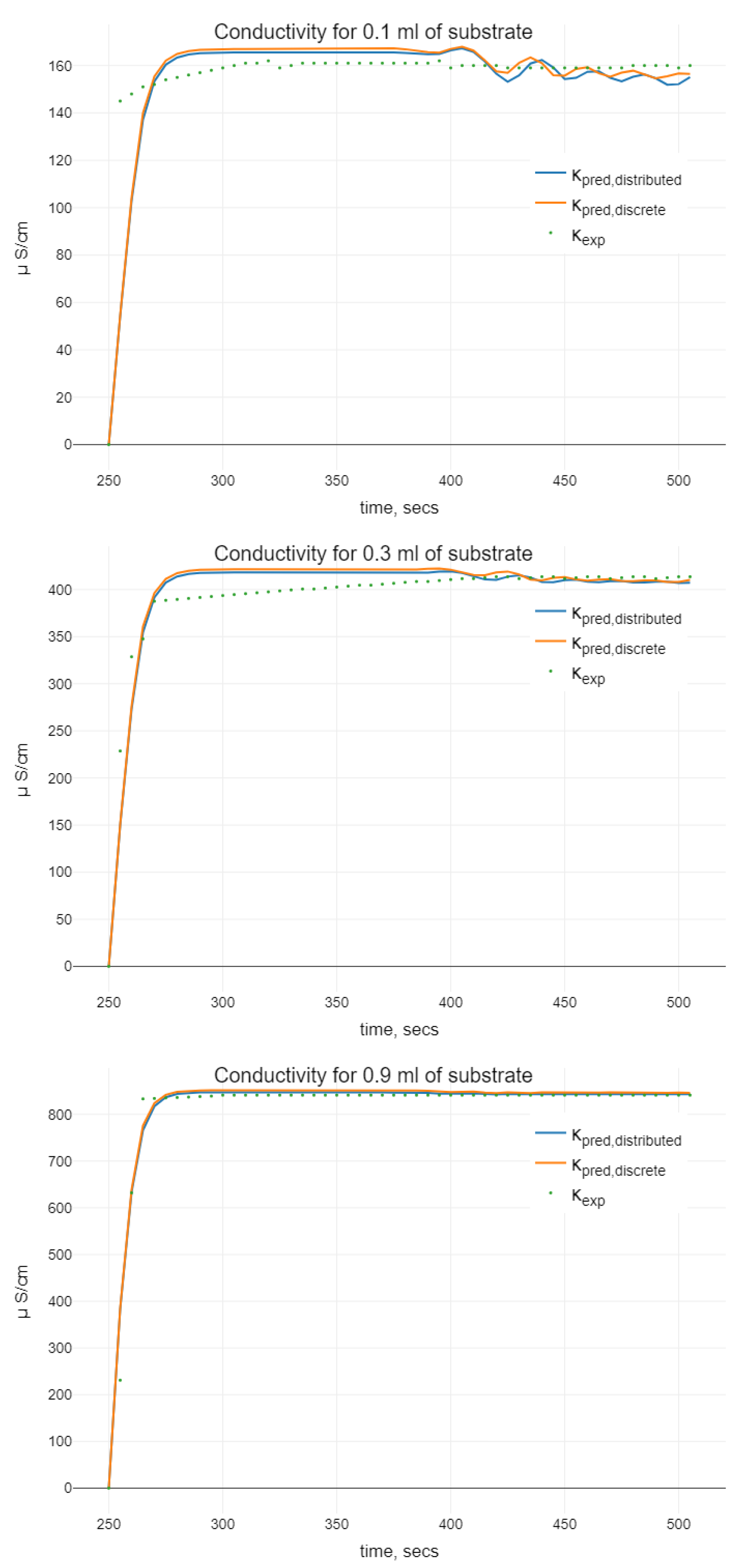
As a result of the experiments, the dependence of the conductivity changes in time was obtained. In the initial stage (0–249 s), the conductivity of the BSA–enzyme complex was tested. When the conductivity remained constant, in the 250th second of the measurement, a substrate of different concentration was added to the BSA–enzyme complex. The addition of a substrate rapidly changed the conductivity of the resulting complex, which may indicate a rapid chemical reaction. By comparing the changes in conductivity as a function of the volume of the substrate added, it can be said that the larger the volume of the substrate added, the greater the change in conductivity was and the more dynamic the increase in conductivity was.
4.1.2. Parameter Estimation for the Experimental Study
Algorithm 1 was implemented in R package. For the integration of (
11), Julia calling was used. Initial parameter values together with the parameter bounds are presented in
Table 1. The solution of the optimization problem (
17) is displayed in the column
. The root mean squared error of the prediction with the obtained model throughout all data series (i.e., for different initial substrate volumes) is 48.74105. The number of iterations was 50.
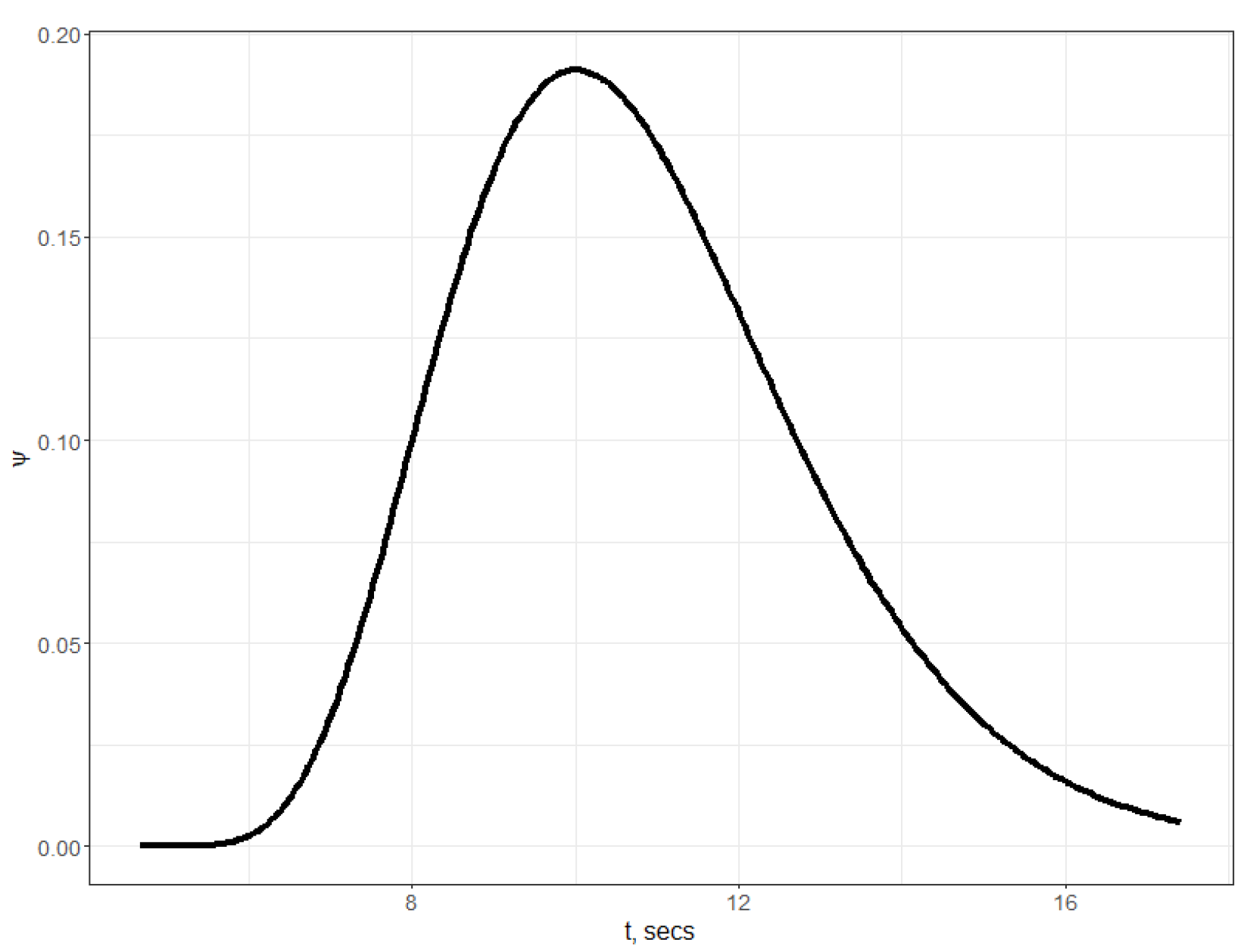
The density function
for estimated parameters is shown on
Figure 2. In
Figure 3, the results of integration of (
11) and (
4) for different initial volumes of substrate which are based on the values of parameters estimated are shown.
4.2. Enzyme–Substrate–Inhibitor Interaction
The second experimental study used butyryl cholinesterase (BuChE) (EC 3.1.1.8, from Horse Serum) with a specific activity of 13 U/mg solid bovine albumin (fraction V, 98% purity), butyryl choline chloride (BuChCl) (98% purity), a-chaconine (95% purity) from potato sprouts, and glutaraldehyde (grade II, 25% aqueous solution), which were purchased from Sigma-Aldrich Chemie GmbH (Steinheim, Germany). All other reagents were of analytical grade and were used without any further treatment.
Biologically active membranes were formed by cross-linking butyryl cholinesterase with BSA on the transducer surface in a saturated glutaraldehyde vapour. The mixture containing 5% (w/v) butyryl cholinesterase, 5% (w/v) BSA, and 10% (w/v) glycerol in 20 mM phosphate buffer (pH 7.2) was deposited on the sensitive surface of one transducer by the drop method, while the mixture of 10% (w/v) BSA and 10 % (w/v) glycerol in 20 mM phosphate buffer (pH 7.2) was placed on the surface of a reference transducer. The sensor chip was then placed in a saturated glutaraldehyde vapor. After a 30 min exposure in glutaraldehyde, the membranes were dried at room temperature for 15 min.
All measurements were performed in daylight at room temperature in an open glass vessel filled with a vigorously stirred 5 mM phosphate buffer solution, pH 7.2. The 200 mM stock solution of BuChCl in deionised HO, and 2 mM stock solution of the a-chaconine in 5 mM acetic acid were prepared. The concentrations of substrates and inhibitors were adjusted by adding defined volumes of the stock solution of proper concentration. The differential output signal between the measuring and reference Ion-Sensitive Field Effect Transistors (ISFETs) was registered using portable device. After the response measurement (determination of enzyme inhibition), the initial enzyme activity was restored by washing out the biosensor enzymatic membrane in the working buffer solution for 10 to 15 min.
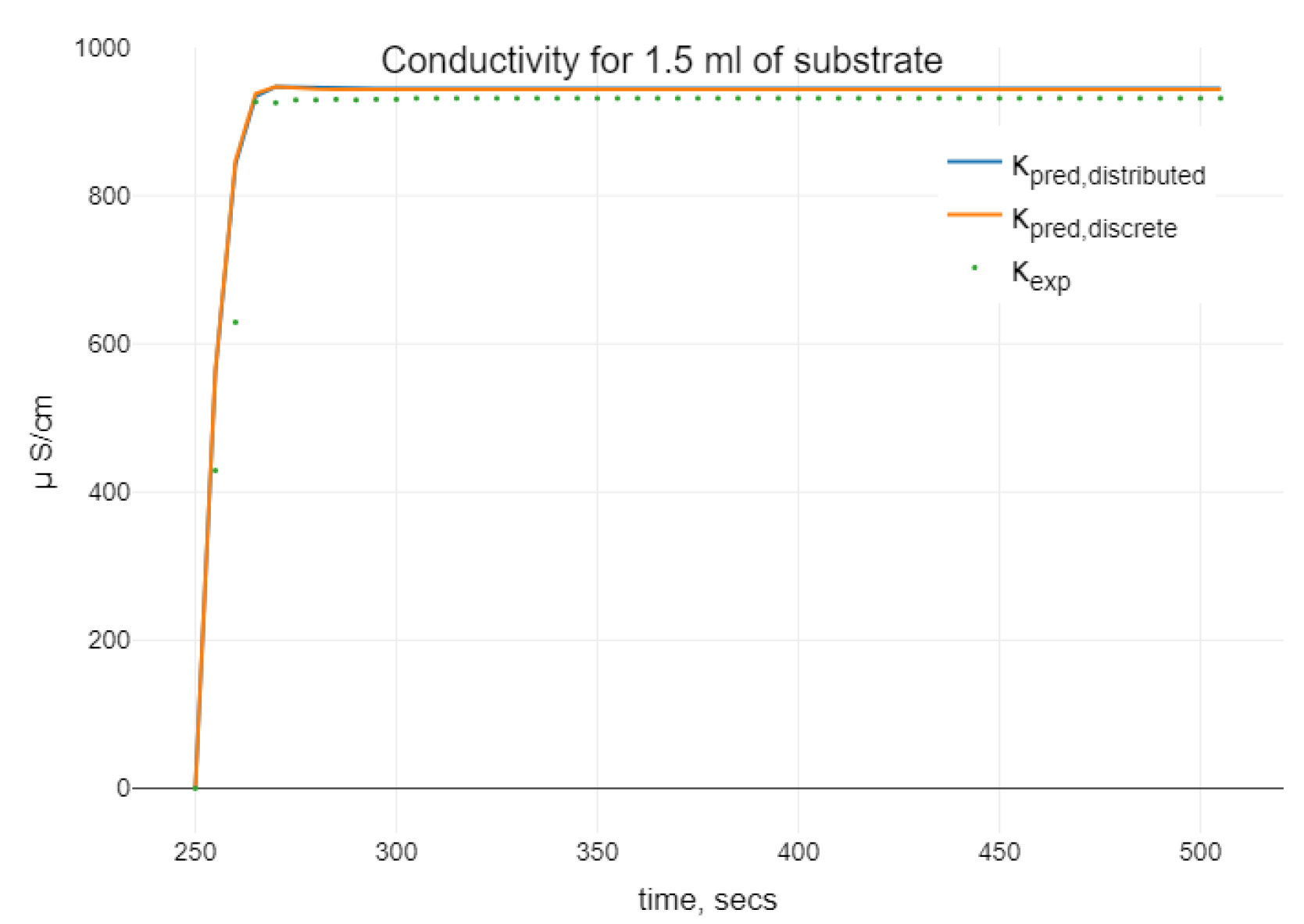
4.3. Numerical Simulation of Enzyme–Substrate–Inhibitor Model
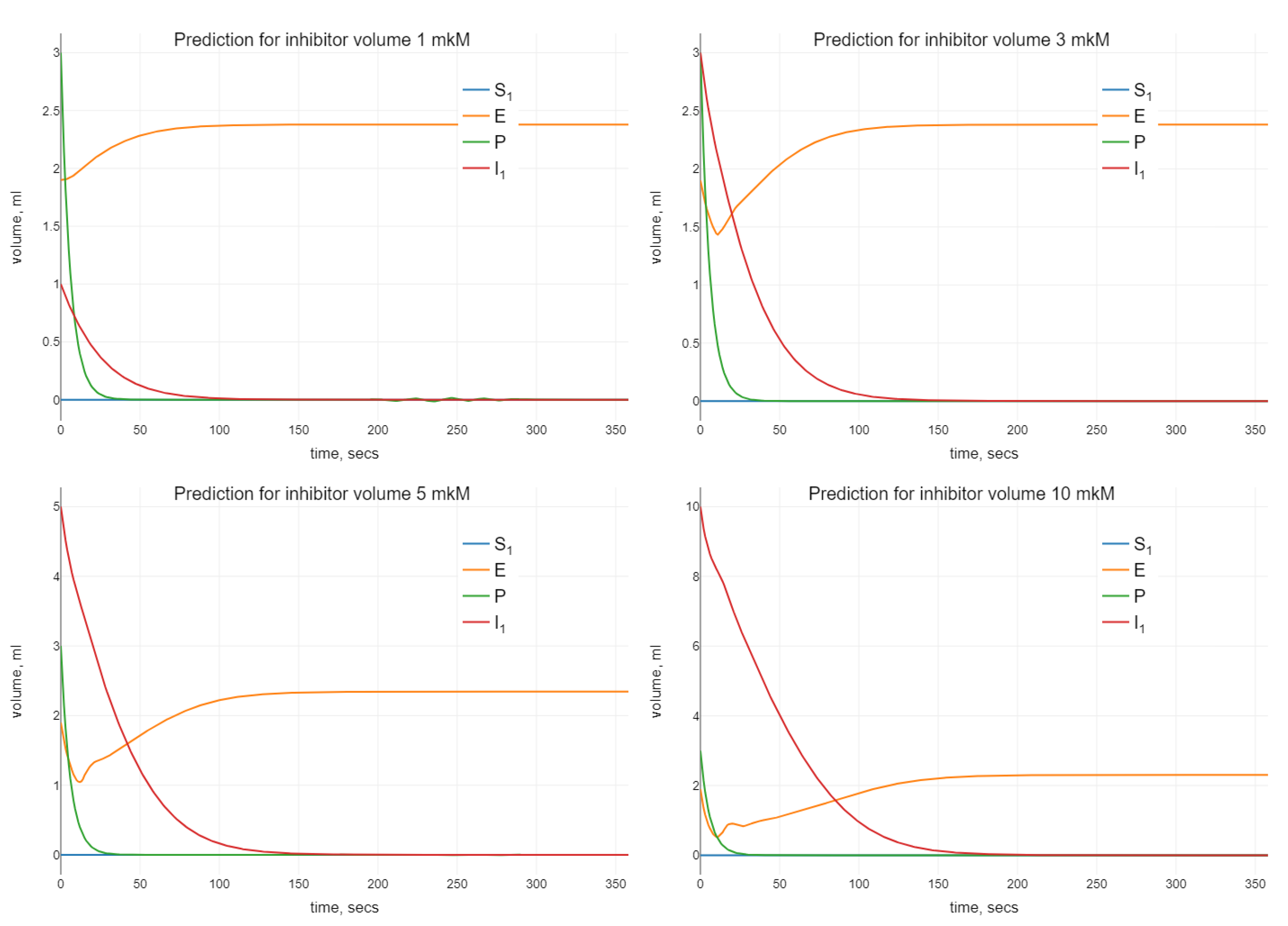
Algorithm 1 was modified for the estimation of the parameters of the model (24). The optimal values of the parameters were used for the simulation using different initial values for the inhibitor. The visualization of the simulations results is presented in
Figure 4.
5. Discussion and Conclusions
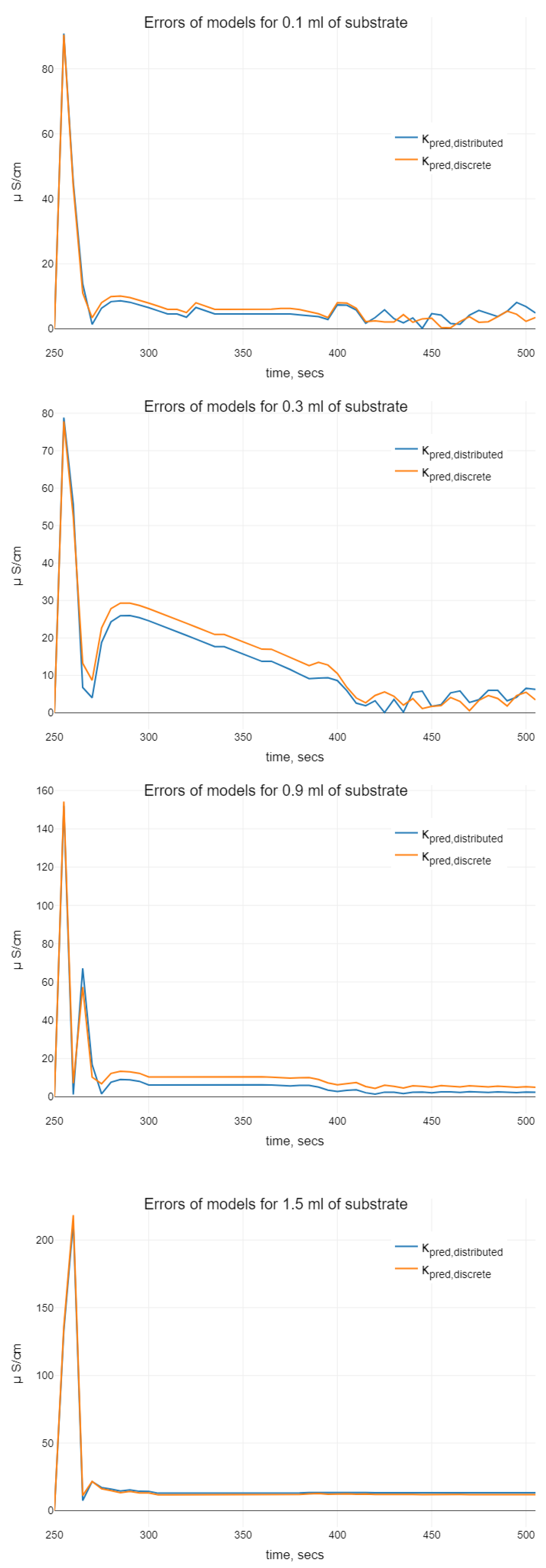
While analyzing the models of (
11) and (
4), with the help of plots on
Figure 3 and
Figure 5, we can conclude the following. It is clearly seen that “undesirable” oscillations in the trajectories are appearing at the smaller initial volumes of the substrate. Then, oscillations disappear for bigger ones. There are some differences between trajectories of models with distributed and discrete delay (we mean that the model with discrete delay (
4) is constructed for the mean value of delay
obtained using the density function
, i.e., for
) observable on the plots. Namely, we see differences in the phases of oscillations. Using the distributed delay, the oscillation starts later. Moreover, for the bigger values of initial substrate volume, the oscillations for the distributed delay model disappear (as it can be seen in case of 1.5 mL of substrate). The amplitudes of the oscillations at the smaller values
are similar, whereas by increasing the initial substrate amount, the oscillations for distributed delay models are not seen. Hence, when comparing two kinds of delay models, which both demonstrate oscillations in certain sense, we conclude that (
11) is more chemically adequate for bigger values of the initial substrate dose than the discrete delay model.
When analyzing the enzyme–substrate–inhibitor model (24), we observe a similar oscillating behavior when increasing the initial amount of inhibitor. Namely, as it can be seen in
Figure 4, small oscillations in
P disappear when we increase
.
In summary, we can conclude that the modeling of the enzyme kinetics based on dynamic systems with distributed delays can make the usage of the delayed differential equations more attractive since it shows more adequate chemical behavior, reducing oscillations when compared with the discrete delays. This is due to more continuous right-sides of the differential equations with distributed delays in comparison to discrete ones. On the other hand, such types of models are complicated enough to demonstrate complex nonlinear behavior, e.g., with the help of using parameters of the density of the delay distribution.
One should bear in mind the computational complexity of the respective models. So, in the case of multi-substrate multi-inhibitor modeling using the model (
6), we need to solve the system of
equations. Using the models with delay (23) or (24), the number of equations determined by the number of phase coordinates is
, which is
smaller. In the case of complicated computing, such as multi-iteration process optimization for the purpose of parameter estimation, this difference in the order of computational complexity is crucial. Even in the case of Julia computing, numerical integration of the systems of differential equations is time-consuming when the system order or accuracy is growing.
Algorithm 1 can be extended to the parameter estimation of
M-substrate
N-inhibitor model (24). In such a case, the parameters to be estimated are reaction rates
,
, three parameters for each density
and
,
,
,
, and
K. That is, we obtain
parameters at the whole, which leads to the corresponding computational complexity. The convergence of such an algorithm is also essential, which is closely dealt with in the search of the initial approximation. We think that in some special cases, the approach presented in
Section 3.2 could be applied for experimental data as well.
The usage of delay models for enzyme kinetics is of importance in the multi-substrate multi-inhibitor cases, as they demonstrate more complex qualitative behavior than the models without delay; for example, outbreaks can repeat in the delayed model, whereas we do not observe this in the case without delay. In the future, we will try to evidence this using experimental data.
Here, we only mention that the application of the delayed models is also preferable from the viewpoint of availability of experimental data. Namely, a typical Michaelis–Menten model is based on the formation of multiple complexes. Their concentrations cannot be assessed from a research point of view. In fact, we measure the product of the reaction through the conductivity of the solution. By using time delays, we do not consider concentrations, but we consider the appearance and disappearance of complexes without having to give concentrations. Moreover, delays can be identified with the help of the algorithm offered above.
In the given work, the study of an enzyme kinetics is based on the chemical response using conductivity. The problem is that in modeling, with the help of mass action law, the exact value of the concentration of the substance is required, whereas the experimental results offer the conductivity. As a rule, they use concentration–conductivity dependencies as linear ones, although it has been shown by Kohlrausch’s studies that the relationships are more complex. In this paper, the assumption for strong electrolytes is used, allowing us to apply the corresponding modification of Kohlrausch law, for which parameters and K were experimental with the help of Algorithm 1. In the future, on the basis of the scheme presented in the paper concerning chemical reactions, we plan to carry out experiments that would take into account conductivity tests depending on many substances, applied one after another and also applied simultaneously. Therefore, the effect of time delays in multi-substrate multi-inhibitor enzyme kinetics modeling is still an open problem, requiring the development of many techniques to apply real experimental data, which offers further possibilities to explore it.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}