


Monitoring of Indoor Farming of Lettuce Leaves for 16 Hours Using Electrical Impedance Spectroscopy (EIS) and Double-Shell Model (DSM)


Abstract
:1. Introduction
2. Theoretical Background
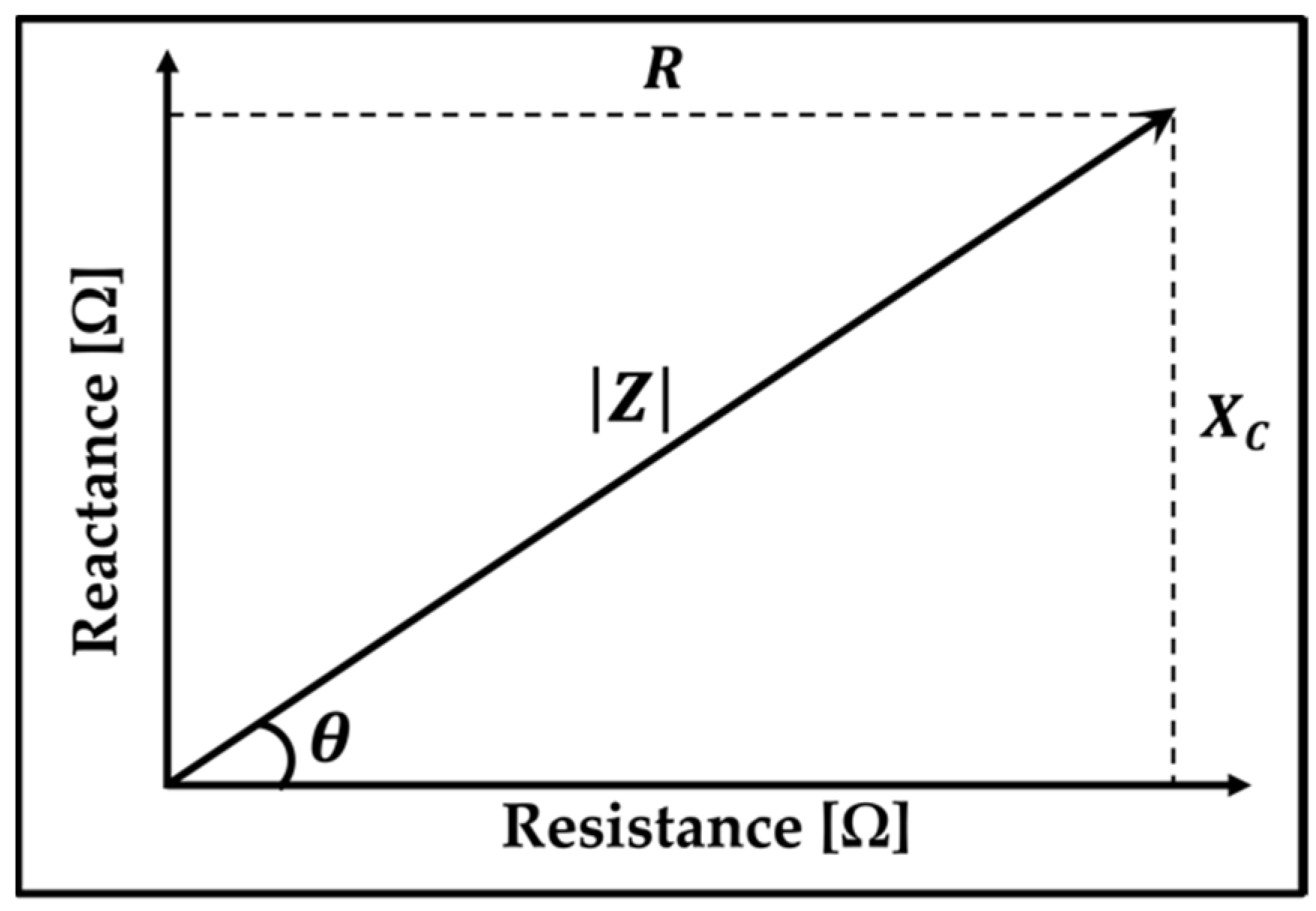
2.1. Electrical Impedance
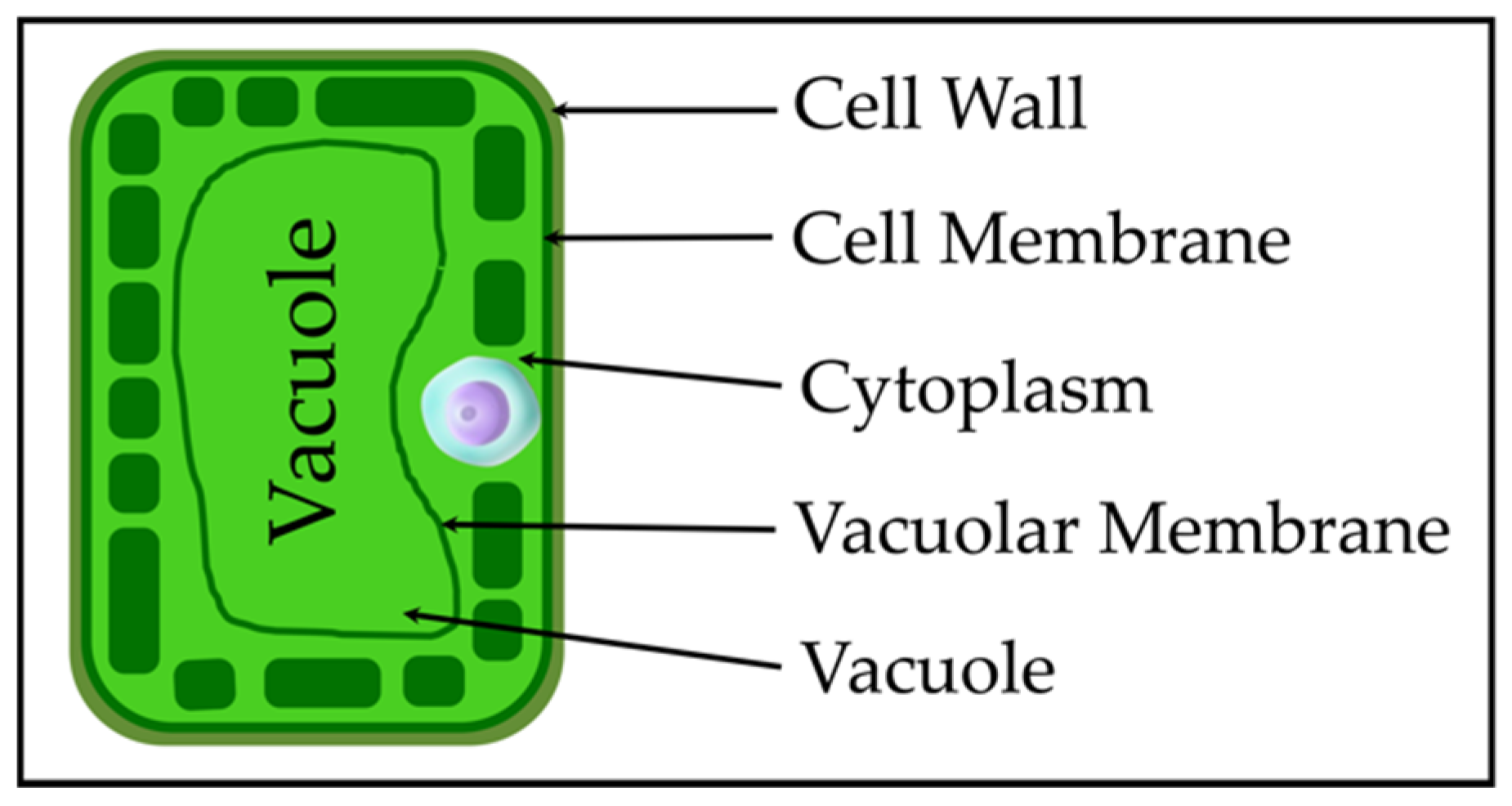
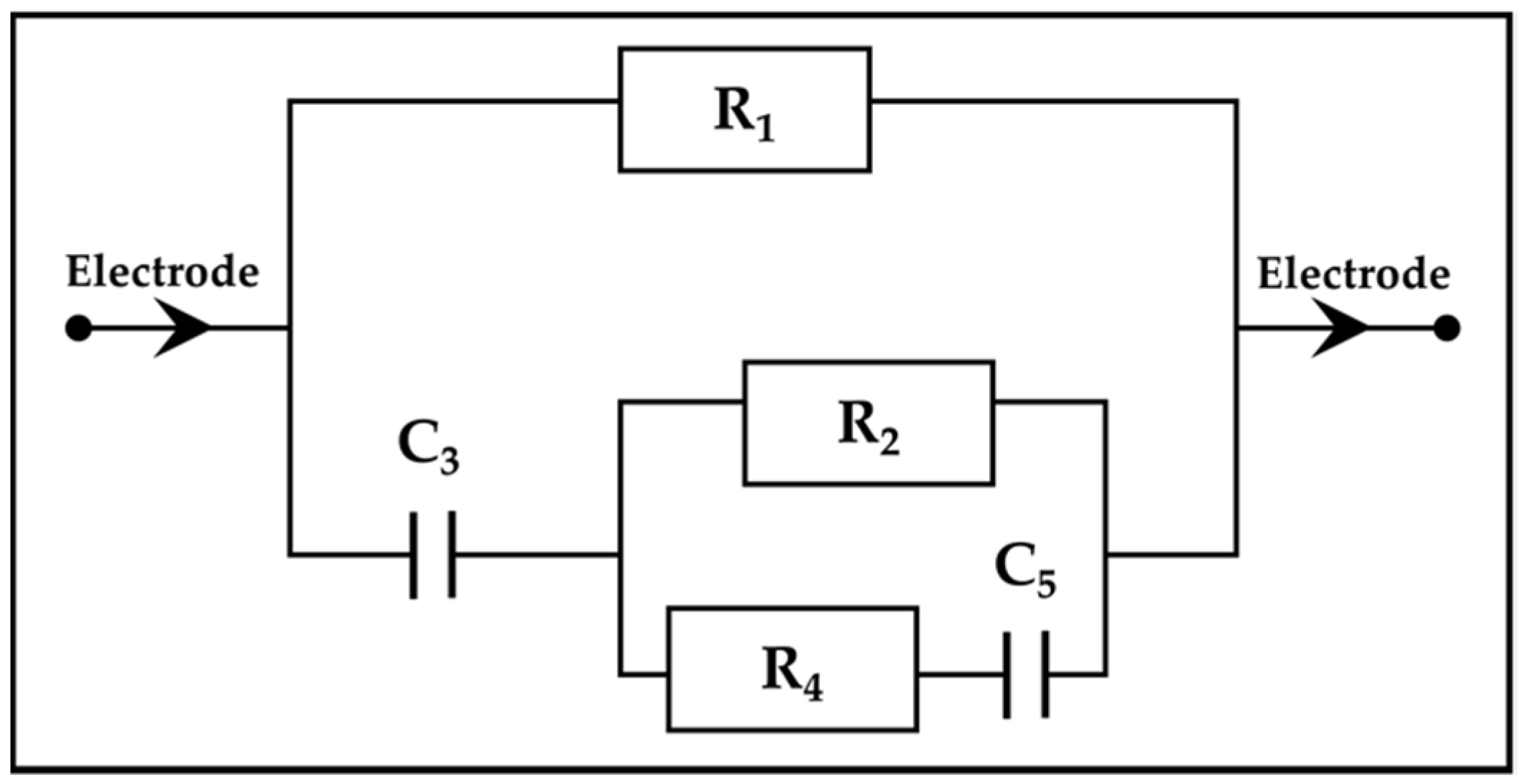
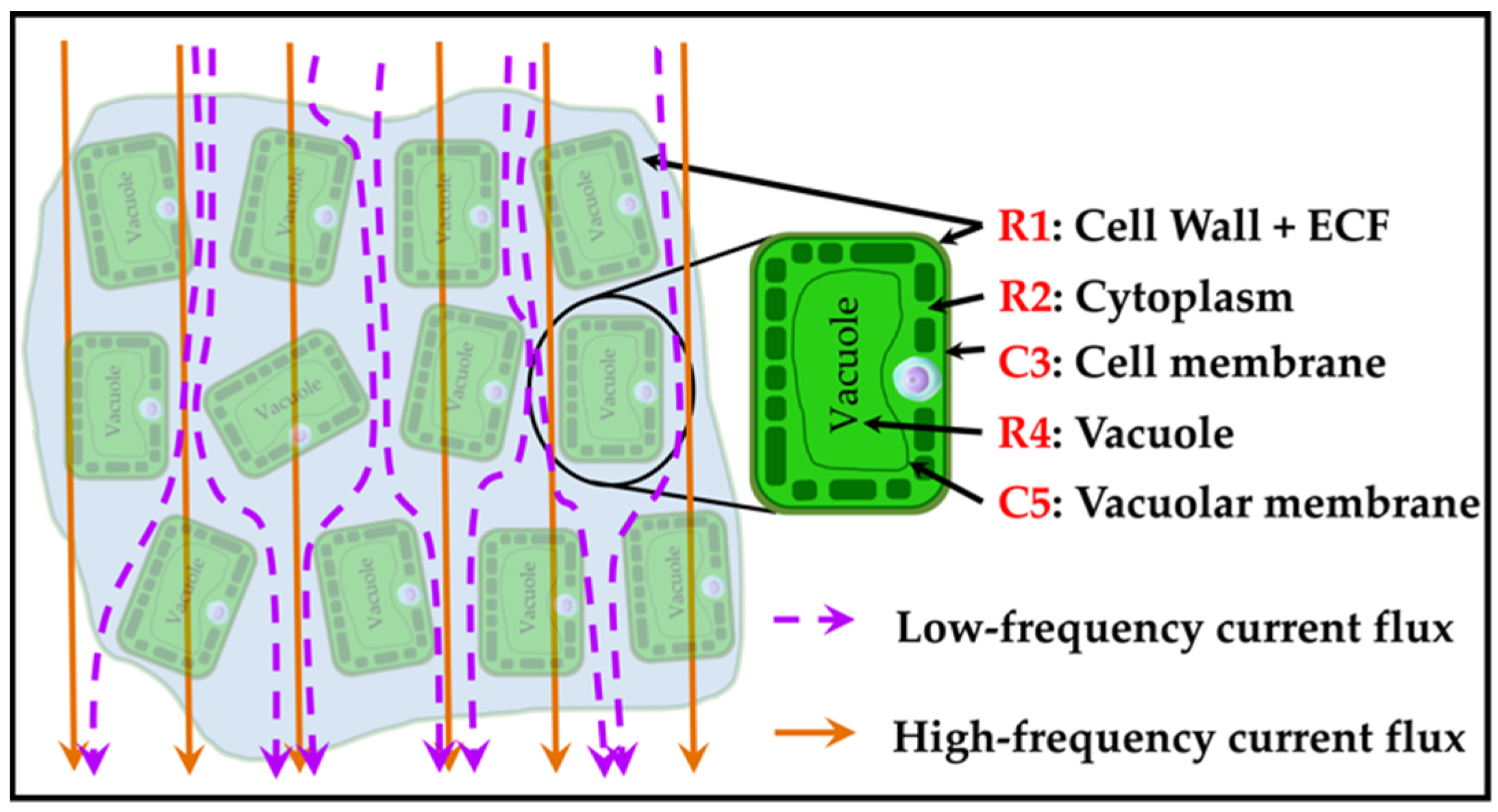
2.2. Plant Cell Anatomy and Double-Shell Model
2.3. LEDs and Indoor Plant Cultivation
3. Materials and Methods
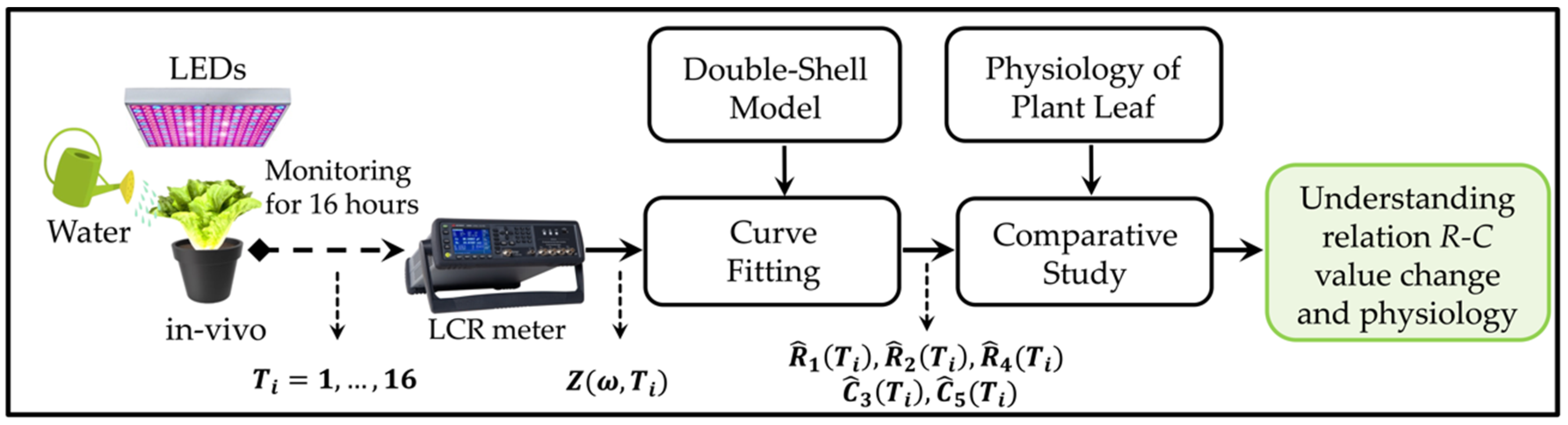
3.1. Experimental Research Overview
- ▪ Lettuce leaves were measured in vivo using an LCR meter for 16 h.
- ▪ The output of the LCR meter is the impedance,
- ▪ A curve-fitting method was used to estimate the R and C values of the DSM.
- ▪ Comparative studies between changes in R-C values and leaf physiology were performed.

3.2. Indoor Growing Space Setup and Lighting
3.3. Impedance Measurement Method and Procedure
- The designed experiment process was as follows. Open-circuit calibration and short-circuit calibration were performed before measurements to calibrate the LCR equipment. Using a pair of electrodes adhered to the lettuce leaves, the impedance of the leaves was measured without harming the plants. Throughout all measurement periods, the two electrodes were kept 3 cm apart. After the measurement, the electrodes were washed, dried, and placed on new leaves to prevent unreliable results.
- The impedance values recorded by the precision LCR equipment were applied to the equivalent circuit, reflecting the growth process of plant tissues. The process of plant growth monitoring was then analyzed and figured out by using a computer to fit the values of the measured impedance parameters to the electrical model. Data were processed using Microsoft Excel 2021 (Microsoft Corporation, Redmond, WA, USA) and graphically analyzed using MATLAB R2022a (The MathWorks, Inc., Natick, MA, USA).
4. Results
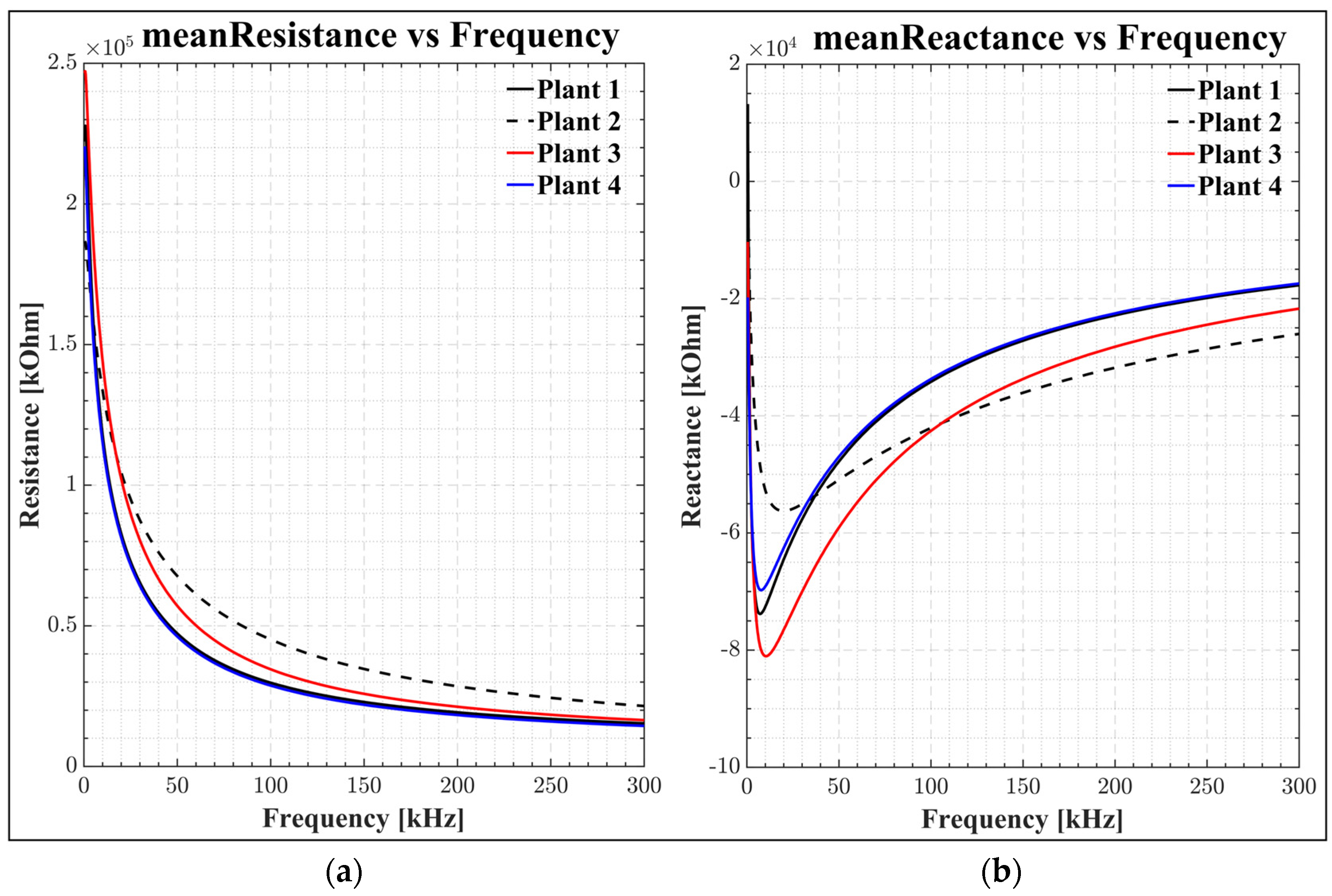
4.1. Frequency-Dependent Resistance and Reactance
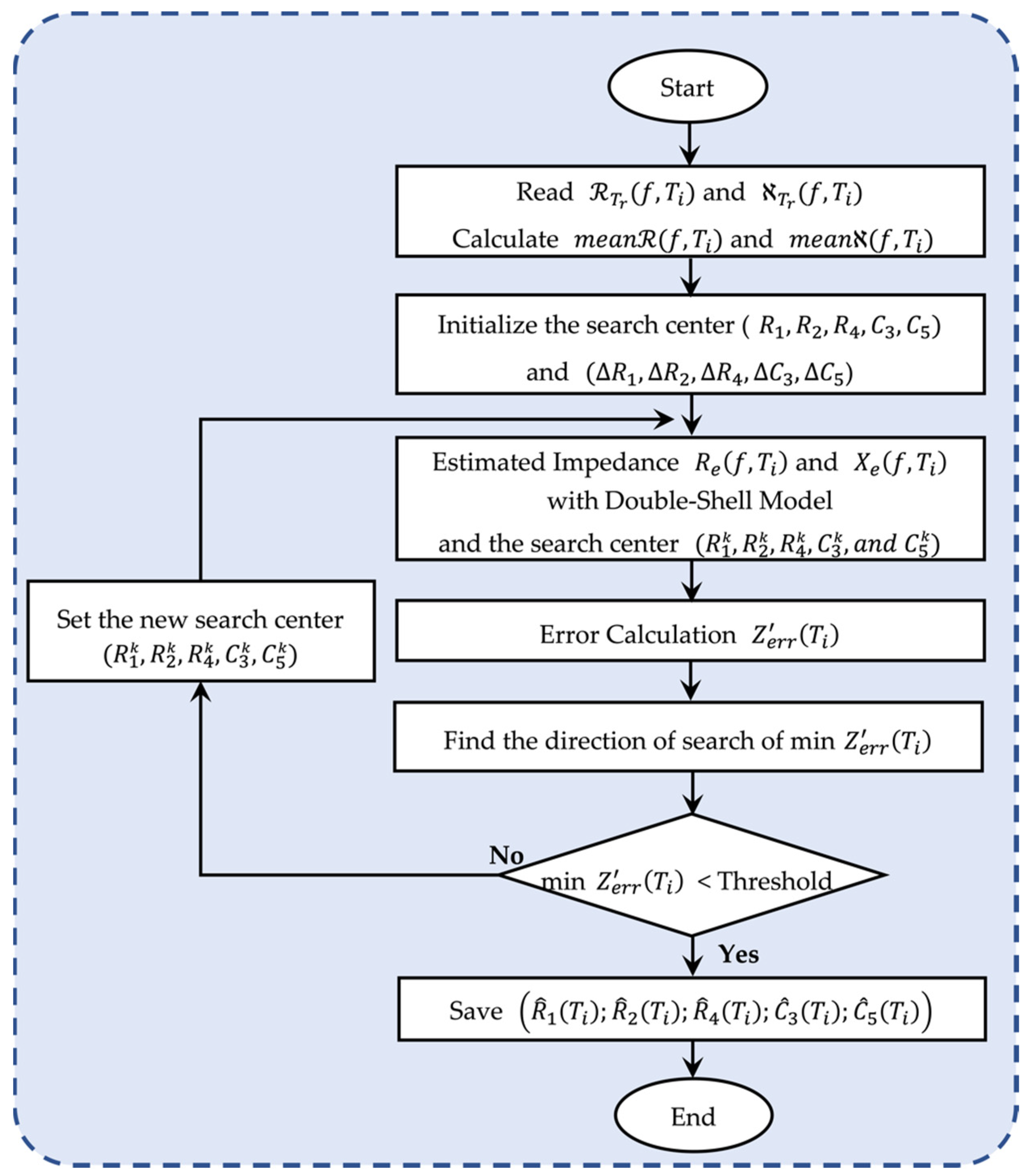
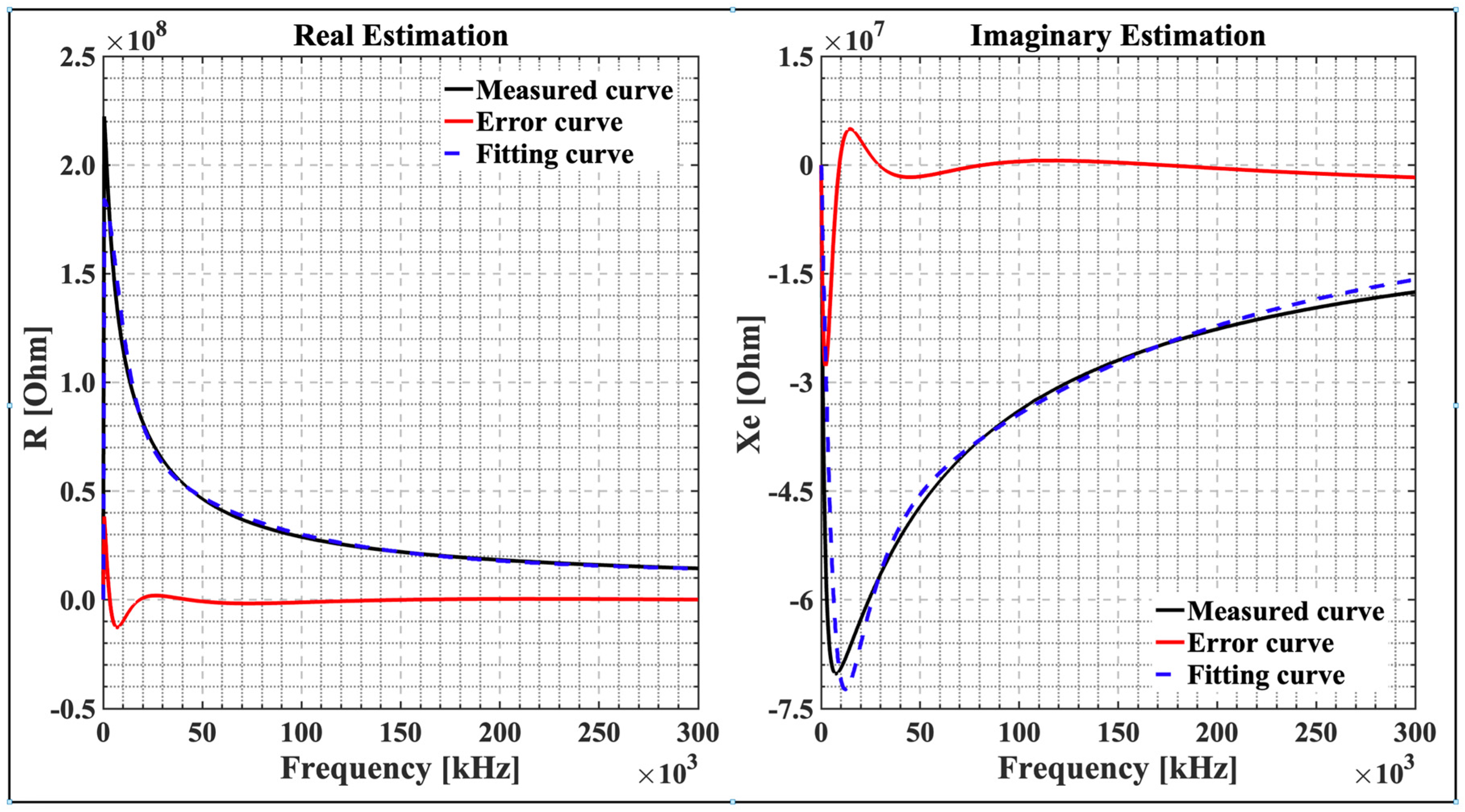
4.2. The Proposed Optimization Flowchart
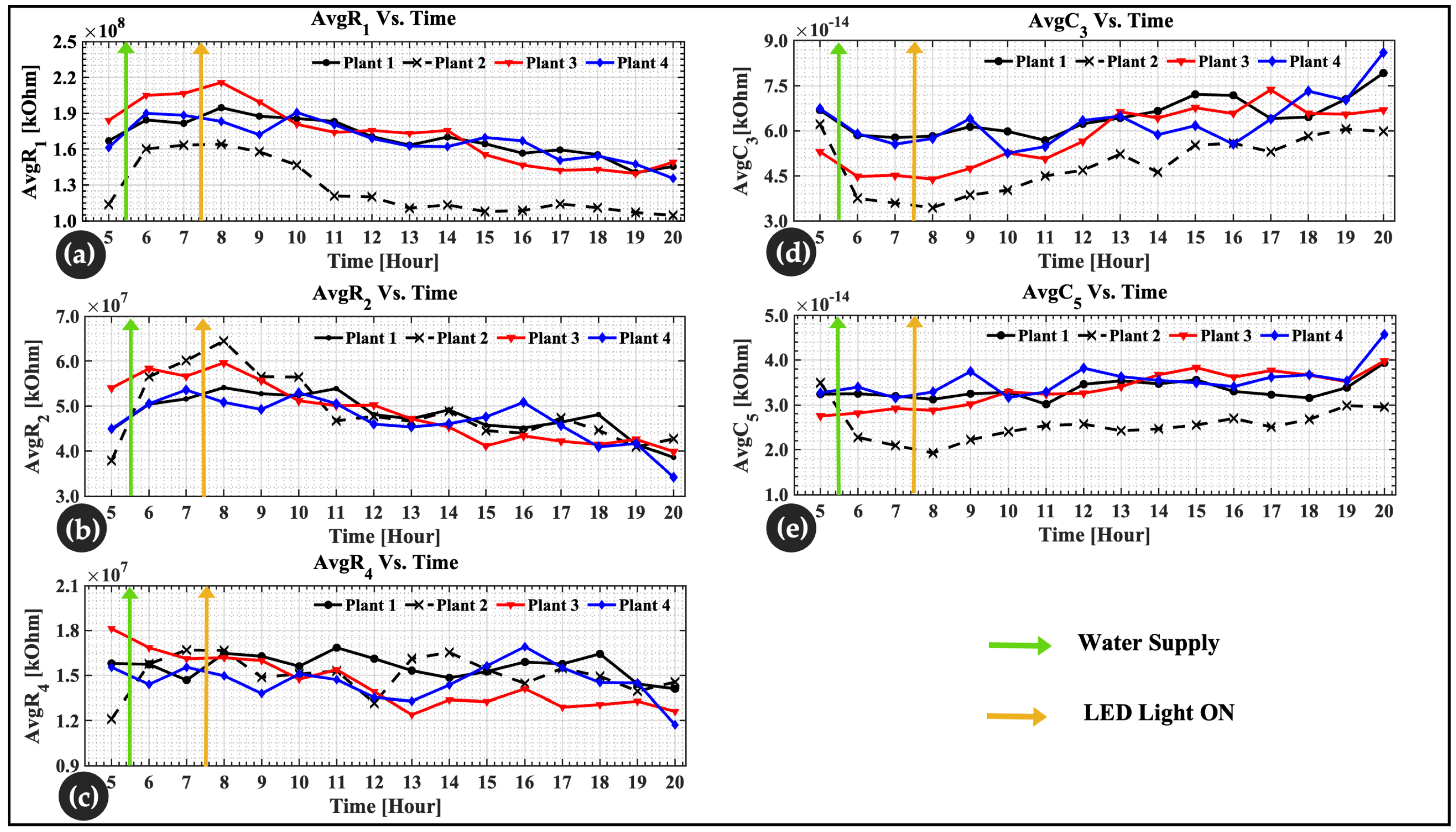
4.3. Physiological Changes in Lettuce Leaves
5. Discussion
6. Conclusions and Future Work
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
Abbreviations and Symbols
| EIS | Electrical impedance spectroscopy |
| R-C values | Resistance and capacitance values |
| DSM | Double-shell model |
| ECF | Extracellular fluid |
| ICF | Intracellular fluid |
| Capacitive reactance | |
| R | Resistance |
| Measured resistance | |
| Measured reactance | |
| Mean value of measured resistance for | |
| Mean value of measured reactance for | |
| Plant index = 1, 2…, 4 | |
| Try index = 1, 2…, 5 | |
| Total number of measurements (5) | |
| Time index = 1, 2…, 16 | |
| Angular frequency | |
| Frequency- and time-dependent impedance | |
| Cell wall + ECF resistance | |
| Cell membrane resistance | |
| Cell membrane capacitance | |
| Vacuole resistance | |
| Vacuolar membrane capacitance | |
| Estimated resistance | |
| Estimated reactance | |
| New search-point candidate from old center point | |
| New search-point candidate from old center point | |
| New search-point candidate from old center point | |
| New search-point candidate from old center point | |
| New search-point candidate from old center point | |
| Defined as iteration | |
| Vector search direction | |
| Positive scalar denoting the distance moved along the search direction | |
| Vector search direction for point candidate | |
| Vector search direction for point candidate | |
| Vector search direction for point candidate | |
| Vector search direction for point candidate | |
| Vector search direction for point candidate | |
| Vector from to | |
| Vector from to | |
| Vector from | |
| Vector from to | |
| Vector from to | |
| Impedance error as a function of frequency | |
| Minimum impedance error as a function of time | |
| Measured impedance as a function of frequency and time | |
| Estimated impedance as a function of frequency and time | |
| Estimated value after curve fitting corresponding to cell wall + ECF resistance | |
| Estimated value after curve fitting corresponding to cell membrane resistance | |
| Estimated value after curve fitting corresponding to vacuole resistance | |
| Estimated value after curve fitting corresponding to cell membrane capacitance | |
| Estimated value after curve fitting corresponding to vacuolar membrane capacitance |
References
- Chaerle, L.; Straeten, D.V.D. Imaging Techniques and the Early Detection of Plant Stress. Trends Plant Sci. 2000, 5, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Mahlein, A.-K. Plant Disease Detection by Imaging Sensors—Parallels and Specific Demands for Precision Agriculture and Plant Phenotyping. Plant Dis. 2016, 100, 241–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolhar-Nordenkampf, H.R.; Long, S.P.; Baker, N.R.; Oquist, G.; Schreiber, U.; Lechner, E.G. Chlorophyll Fluorescence as a Probe of the Photosynthetic Competence of Leaves in the Field: A Review of Current Instrumentation. Funct. Ecol. 1989, 3, 497–514. [Google Scholar] [CrossRef]
- Zhang, J.; Pu, R.; Huang, W.; Yuan, L.; Luo, J.; Wang, J. Using In-Situ Hyperspectral Data for Detecting and Discriminating Yellow Rust Disease from Nutrient Stresses. Field Crops Res. 2012, 134, 165–174. [Google Scholar] [CrossRef]
- Montanha, G.S.; Rodrigues, E.S.; Marques, J.P.R.; de Almeida, E.; dos Reis, A.R.; Pereira de Carvalho, H.W. X-ray Fluorescence Spectroscopy (XRF) Applied to Plant Science: Challenges towards in Vivo Analysis of Plants. Metallomics 2020, 12, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Jarquín, S.; Herrera-Ubaldo, H.; de Folter, S.; Winkler, R. In Vivo Monitoring of Nicotine Biosynthesis in Tobacco Leaves by Low-Temperature Plasma Mass Spectrometry. Talanta 2018, 185, 324–327. [Google Scholar] [CrossRef] [PubMed]
- Altangerel, N.; Ariunbold, G.O.; Gorman, C.; Alkahtani, M.H.; Borrego, E.J.; Bohlmeyer, D.; Hemmer, P.; Kolomiets, M.V.; Yuan, J.S.; Scully, M.O. In Vivo Diagnostics of Early Abiotic Plant Stress Response via Raman Spectroscopy. Proc. Natl. Acad. Sci. USA 2017, 114, 3393–3396. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.; Jeon, E.; Park, K.S.; Yeo, K.-H.; Lee, J. Monitoring of Water Transportation in Plant Stem With Microneedle Sap Flow Sensor. J. Microelectromech. Syst. 2018, 27, 440–447. [Google Scholar] [CrossRef]
- Daskalakis, S.N.; Goussetis, G.; Assimonis, S.D.; Tentzeris, M.M.; Georgiadis, A. A UW Backscatter-Morse-Leaf Sensor for Low-Power Agricultural Wireless Sensor Networks. IEEE Sens. J. 2018, 18, 7889–7898. [Google Scholar] [CrossRef] [Green Version]
- Coppedè, N.; Janni, M.; Bettelli, M.; Maida, C.L.; Gentile, F.; Villani, M.; Ruotolo, R.; Iannotta, S.; Marmiroli, N.; Marmiroli, M.; et al. An In Vivo Biosensing, Biomimetic Electrochemical Transistor with Applications in Plant Science and Precision Farming. Sci. Rep. 2017, 7, 16195. [Google Scholar] [CrossRef] [PubMed]
- AD5933 Datasheet and Product Info | Analog Devices. Available online: https://www.analog.com/en/products/ad5933.html (accessed on 1 September 2022).
- Kernbach, S. Device for Measuring the Plant Physiology and Electrophysiology. arXiv 2022. [Google Scholar] [CrossRef]
- Jócsák, I.; Végvári, G.; Vozáry, E. Electrical Impedance Measurement on Plants: A Review with Some Insights to Other Fields. Theor. Exp. Plant Physiol. 2019, 31, 359–375. [Google Scholar] [CrossRef] [Green Version]
- Repo, T.; Laukkanen, J.; Silvennoinen, R. Measurement of the Tree Root Growth Using Electrical Impedance Spectroscopy. Silva Fenn. 2005, 39, 380. [Google Scholar] [CrossRef] [Green Version]
- Väinölä, A.; Repo, T. Impedance Spectroscopy in Frost Hardiness Evaluation of Rhododendron Leaves. Ann. Bot. 2000, 86, 799–805. [Google Scholar] [CrossRef] [Green Version]
- Bera, T.K.; Bera, S.; Chowdhury, A.; Ghoshal, D.; Chakraborty, B. Electrical Impedance Spectroscopy (EIS) Based Fruit Characterization; Informa UK Ltd.: London, UK, 2017. [Google Scholar]
- Harker, F.R.; Maindonald, J.H. Ripening of Nectarine Fruit (Changes in the Cell Wall, Vacuole, and Membranes Detected Using Electrical Impedance Measurements). Plant Physiol. 1994, 106, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Kuson, P.; Terdwongworakul, A. Minimally-Destructive Evaluation of Durian Maturity Based on Electrical Impedance Measurement. J. Food Eng. 2013, 116, 50–56. [Google Scholar] [CrossRef]
- Żywica, R.; Banach, J.K. Simple Linear Correlation between Concentration and Electrical Properties of Apple Juice. J. Food Eng. 2015, 158, 8–12. [Google Scholar] [CrossRef]
- Zheng, L.; Wang, Z.; Sun, H.; Zhang, M.; Li, M. Real-Time Evaluation of Corn Leaf Water Content Based on the Electrical Property of Leaf. Comput. Electron. Agric. 2014, 112, 102–109. [Google Scholar] [CrossRef]
- Hayden, R.I.; Moyse, C.A.; Calder, F.W.; Crawford, D.P.; Fensom, D.S. Electrical Impedance Studies on Potato and Alfalfa Tissue. J. Exp. Bot. 1969, 20, 177–200. [Google Scholar] [CrossRef]
- Paszewski, A.; Bulanda, W.; Dziubiñska, H.; Trębacz, K. Resistance and Capacity in the Thallus of Marchantia Polymorpha. Physiol. Plant. 1982, 54, 213–220. [Google Scholar] [CrossRef]
- MacDougall, R.G.; Thompson, R.G.; Piene, H. Stem Electrical Capacitance and Resistance Measurements as Related to Total Foliar Biomass of Balsam Fir Trees. Can. J. For. Res. 1987, 17, 1071–1074. [Google Scholar] [CrossRef]
- Zhang, M.I.N.; Willison, J.H.M. Electrical Impedance Analysis in Plant Tissues11. J. Exp. Bot. 1991, 42, 1465–1475. [Google Scholar] [CrossRef]
- SharathKumar, M.; Heuvelink, E.; Marcelis, L.F.M. Vertical Farming: Moving from Genetic to Environmental Modification. Trends Plant Sci. 2020, 25, 724–727. [Google Scholar] [CrossRef] [PubMed]
- Caballero, B.; Finglas, P.; Toldrá, F. Encyclopedia of Food and Health, 1st ed.; Academic Press: Cambridge, MA, USA, 2015; ISBN 978-0-12-384947-2. [Google Scholar]
- Landrein, B.; Hamant, O. How Mechanical Stress Controls Microtubule Behavior and Morphogenesis in Plants: History, Experiments and Revisited Theories. Plant J. Cell Mol. Biol. 2013, 75, 324–338. [Google Scholar] [CrossRef]
- Camargo, A.; Smith, J.S. Image Pattern Classification for the Identification of Disease Causing Agents in Plants. Comput. Electron. Agric. 2009, 66, 121–125. [Google Scholar] [CrossRef]
- Bock, C.H.; Parker, P.E.; Cook, A.Z.; Gottwald, T.R. Visual Rating and the Use of Image Analysis for Assessing Different Symptoms of Citrus Canker on Grapefruit Leaves. Plant Dis. 2008, 92, 530–541. [Google Scholar] [CrossRef] [Green Version]
- Wijekoon, C.P.; Goodwin, P.H.; Hsiang, T. Quantifying Fungal Infection of Plant Leaves by Digital Image Analysis Using Scion Image Software. J. Microbiol. Methods 2008, 74, 94–101. [Google Scholar] [CrossRef]
- Dean, D.A.; Ramanathan, T.; Machado, D.; Sundararajan, R. Electrical Impedance Spectroscopy Study of Biological Tissues. J. Electrost. 2008, 66, 165–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macdonald, J.R.; Barsoukov, E. Impedance Spectroscopy: Theory, Experiment, and Applications; Wiley.com; John Wiley & Sons: Hoboken, NJ, USA, 2018; ISBN 978-1-119-33317-3. [Google Scholar]
- Hall, J.L.; Flowers, T.J.; Roberts, R.M. Plant Cell Structure and Metabolism; Longman: Harlow, UK, 1984. [Google Scholar]
- Zhang, M.I.N.; Stout, D.G.; Willison, J.H.M. Electrical Impedance Analysis in Plant Tissues3. J. Exp. Bot. 1990, 41, 371–380. [Google Scholar] [CrossRef]
- Harker, F.R.; Forbes, S.K. Ripening and Development of Chilling Injury in Persimmon Fruit: An Electrical Impedance Study. N. Z. J. Crops Hortic. Sci. 1997, 25, 149–157. [Google Scholar] [CrossRef]
- Bauchot, A.D.; Harker, F.R.; Arnold, W.M. The Use of Electrical Impedance Spectroscopy to Assess the Physiological Condition of Kiwifruit. Postharvest Biol. Technol. 2000, 18, 9–18. [Google Scholar] [CrossRef]
- Zhang, M.I.N.; Willison, J.H.M. Electrical Impedance Analysis in Plant Tissues8. J. Exp. Bot. 1993, 44, 1369–1375. [Google Scholar] [CrossRef]
- Duong, T.N.; Takamura, T.; Watanabe, H.; Tanaka, M. Light Emitting Diodes (LEDs) as a Radiation Source for Micropropagation of Strawberry. In Transplant Production in the 21st Century: Proceedings of the International Symposium on Transplant Production in Closed System for Solving the Global Issues on Environmental Conservation, Food, Resources and Energy; Kubota, C., Chun, C., Eds.; Springer: Dordrecht, The Netherland, 2000; pp. 114–118. ISBN 978-94-015-9371-7. [Google Scholar]
- Xu, H.; Xu, Q.; Li, F.; Feng, Y.; Qin, F.; Fang, W. Applications of Xerophytophysiology in Plant Production—LED Blue Light as a Stimulus Improved the Tomato Crop. Sci. Hortic. 2012, 148, 190–196. [Google Scholar] [CrossRef]
- Han, T.; Vaganov, V.; Cao, S.; Li, Q.; Ling, L.; Cheng, X.; Peng, L.; Zhang, C.; Yakovlev, A.N.; Zhong, Y.; et al. Improving “Color Rendering” of LED Lighting for the Growth of Lettuce. Sci. Rep. 2017, 7, 45944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.-H.; Wheeler, R.M.; Sager, J.C.; Yorio, N.C.; Goins, G.D. Light-Emitting Diodes as an Illumination Source for Plants: A Review of Research at Kennedy Space Center. Habitation 2005, 10, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Everitt, B.S.; Skrondal, A. The Cambridge Dictionary of Statistics, 4th ed.; Cambridge University Press: Cambridge, UK, 2010; ISBN 978-0-511-78827-7. [Google Scholar]
- Kotz, S.; Read, C.B.; Balakrishnan, N. Brani Vidakovic Encyclopedia of Statistical Sciences, 16 Volume Set, 2nd Edition | Wiley. Available online: https://www.wiley.com/en-us/Encyclopedia+of+Statistical+Sciences%2C+16+Volume+Set%2C+2nd+Edition-p-9780471150442 (accessed on 22 November 2022).
- Edgar, T.F.; Himmelblau, D.M.; Lasdon, L.S. Optimization of Chemical Processes, 2nd, illustrated ed.; McGraw-Hill: New York, NY, USA, 2001; ISBN 978-0-07-039359-2. [Google Scholar]
- Ackmann, J.J.; Seitz, M.A. Methods of Complex Impedance Measurements in Biologic Tissue. Crit. Rev. Biomed. Eng. 1984, 11, 281–311. [Google Scholar]
- Wilner, J.; Brach, E.J. Utilization of Bioelectric Tests in Biological Research. Rep. Agric. Can. Eng. Stat. Res. Inst. Can. 1979, 40, 630–637. [Google Scholar]
- Ings, J.; Mur, L.A.J.; Robson, P.R.H.; Bosch, M. Physiological and Growth Responses to Water Deficit in the Bioenergy Crop Miscanthus x Giganteus. Front. Plant Sci. 2013, 4, 468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Ario, M.; Sablowski, R. Cell Size Control in Plants. Annu. Rev. Genet. 2019, 53, 45–65. [Google Scholar] [CrossRef]
- Chen, P.; Jung, N.U.; Giarola, V.; Bartels, D. The Dynamic Responses of Cell Walls in Resurrection Plants During Dehydration and Rehydration. Front. Plant Sci. 2020, 10, 1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Responses of Plants to Environmental Stress, 2nd ed.; Volume 1: Chilling, Freezing, and High Temperature Stresses. Available online: https://www.cabdirect.org/cabdirect/abstract/19802605739 (accessed on 3 December 2022).
- Phillips, J.R.; Fischer, E.; Baron, M.; van den Dries, N.; Facchinelli, F.; Kutzer, M.; Rahmanzadeh, R.; Remus, D.; Bartels, D. Lindernia Brevidens: A Novel Desiccation-Tolerant Vascular Plant, Endemic to Ancient Tropical Rainforests. Plant J. Cell Mol. Biol. 2008, 54, 938–948. [Google Scholar] [CrossRef] [PubMed]
- Jung, N.U.; Giarola, V.; Chen, P.; Knox, J.P.; Bartels, D. Craterostigma Plantagineum Cell Wall Composition Is Remodelled during Desiccation and the Glycine-Rich Protein CpGRP1 Interacts with Pectins through Clustered Arginines. Plant J. Cell Mol. Biol. 2019, 100, 661–676. [Google Scholar] [CrossRef] [PubMed]
- King, G.A.; Henderson, K.G.; Lill, R.E. Ultrastructural Changes in the Nectarine Cell Wall Accompanying Ripening and Storage in a Chilling-Resistant and Chilling-Sensitive Cultivar. N. Z. J. Crop Hortic. Sci. 1989, 17, 337–344. [Google Scholar] [CrossRef]
- Agarwal, U.P.; Ralph, S.A.; Reiner, R.S.; Hunt, C.G.; Baez, C.; Ibach, R.; Hirth, K.C. Production of High Lignin-Containing and Lignin-Free Cellulose Nanocrystals from Wood. Cellulose 2018, 25, 5791–5805. [Google Scholar] [CrossRef]
- Pennisi, G.; Pistillo, A.; Orsini, F.; Cellini, A.; Spinelli, F.; Nicola, S.; Fernandez, J.A.; Crepaldi, A.; Gianquinto, G.; Marcelis, L.F.M. Optimal Light Intensity for Sustainable Water and Energy Use in Indoor Cultivation of Lettuce and Basil under Red and Blue LEDs. Sci. Hortic. 2020, 272, 109508. [Google Scholar] [CrossRef]
- Weaver, G.; van Iersel, M.W. Photochemical Characterization of Greenhouse-Grown Lettuce (Lactuca Sativa L. ‘Green Towers’) with Applications for Supplemental Lighting Control. HortScience 2019, 54, 317–322. [Google Scholar] [CrossRef] [Green Version]
- Palmer, S.; van Iersel, M.W. Increasing Growth of Lettuce and Mizuna under Sole-Source LED Lighting Using Longer Photoperiods with the Same Daily Light Integral. Agronomy 2020, 10, 1659. [Google Scholar] [CrossRef]
- Slatyer, R.O.; Markus, D.K. Plant-Water Relationships. Soil Sci. 1968, 106, 478. [Google Scholar] [CrossRef]
- Jiang, Y.-T.; Yang, L.-H.; Ferjani, A.; Lin, W.-H. Multiple Functions of the Vacuole in Plant Growth and Fruit Quality. Mol. Hortic. 2021, 1, 4. [Google Scholar] [CrossRef]
- Raven, J.A. The Role of Vacuoles. New Phytol. 1987, 106, 357–422. [Google Scholar] [CrossRef]
- Juansah, J.; Budiastra, I.W.; Dahlan, K.; Seminar, K.B. Electrical Behavior of Garut Citrus Fruits During Ripening Changes in Resistance and Capacitance Models of Internal Fruits. Int. J. Eng. 2012, 12, 9. [Google Scholar]
- Briggs, G.E. Some Aspects of Free Space in Plant Tissues. New Phytol. 1957, 56, 305–324. [Google Scholar] [CrossRef]
- Tan, X.; Li, K.; Wang, Z.; Zhu, K.; Tan, X.; Cao, J. A Review of Plant Vacuoles: Formation, Located Proteins, and Functions. Plants 2019, 8, 327. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Research Areas | Technique(s)/References | Research Focus | Finding(s) | Application(s) | Limitation(s) | Author(s) |
|---|---|---|---|---|---|---|
| Image monitoring | Thermal imaging [1] | Nondestructive plant physiology monitoring | Stress at an early stage was alleviated. Irreversible damage and yield loss were prevented. | Scales | A few square centimeters can be studied | Chaerle L. et al., 2000. |
| RGB imaging [2,28,29,30] | Identification, quantification, and monitoring of plant diseases | Diseases in Cotton, Apple, Grapefruit, and Canadian goldenrod were identified. | Precision in agriculture, plant phenotyping | Low image quality, low detection accuracy | Mahlein, 2016 Camargo and Smith, 2009; Bock et al., 2008; Wijekoon et al., 2008 | |
| Fluorescent imaging [3] | Leaf damage without visible signs, photosynthesis analysis, and fluorimeter comparison of PSM, MFMS, PAM101 | A significant difference between same-population leaves and photosynthesis changes was observed. Tree and branch damage patterns were identified. | Natural vegetation, ecological research | Application-dependent | Bolhar-Nordenjampf et al., 1989 | |
| Hyperspectral imaging [4] | Ground-based hyperspectral reflectance of yellow rust disease inoculation. Nutrient-stressed treatment to detect and discriminate yellow rust disease from nutrients. | At major growth stages, four vegetation indices clearly responded to disease. Disease and nutrient stress affected most spectral features. The physiological reflectance index was disease-sensitive. | Disease monitoring and mapping | Cost and complexity | Zhang J et al., 2012 | |
| Spectroscopy | X-ray fluorescence [5] | Benchtop XRF to evaluate the elemental distribution change in living plant tissue exposed to X-rays | Higher Zn content than Mn in stems was found. The latter micronutrient presented a higher concentration in leaf veins. | Plant tissue analyses under in vivo conditions | X-rays injure biological tissue | Montanha et al., 2020 |
| Mass [6] | Monitoring the auxin-regulated nicotine biosynthesis in tobacco and evaluating possible biological effects | Rupture of trichomes and cell damage were observed on spots exposed to Low-Temperature Plasma. | Biosynthesis of plant surface in vivo measurement | Destructive to live cell structure | Martínez-Jarquín et al., 2018 | |
| Raman [7] | High-throughput stress phenotyping of plant measurement | Unique negative correlation between concentration levels of anthocyanins and carotenoids was observed. | Plant stress in vivo | Destructive method | Altangerel et al., 2017 | |
| Electrical-based sensor approaches | Microneedle electrodes [8,9] | Measure the xylem sap flow to understand plant physiology | Good adaptation of the microneedle probe in the plant tissue was possible. | Plant physiology (tomato) | Not accurate | Baek et al., 2018; Daskalakis et al., 2018 |
| Organic electrochemical transistor [10] | Real-time monitoring of the electrolyte of tomato plant’s physiological state | A circadian pattern of variation was revealed, which shows the possibility to detect signs of abiotic stress. | Precision farming, plant physiology | Slow response to plant change, low accuracy | Coppedè et al., 2017 | |
| EIS | ZARC-cole and CPE model; CNLS using LEVM7 [14] | Develop EIS for nondestructively evaluating plant root growth of willows | Sum of R1 and R2 in the distributed electric model decreased with an increase in root mass. | Root growth assessment | Minor damage due to insertion of needle’s electrode | Repo et al., 2005 |
| Single-DCE model; CNLS using LEVM v.6 program [15] | Cold acclimation and measurement of frost hardening. | Both quantitative and qualitative changes in cell membranes and water-status were observed. EIS results indicate weaker hardiness than other tests. | Frost hardening capability measurement | System dependent with limited functionality | Väinölä and Repo, 2000 | |
| Different models; nlmin function in S-PLUS [16,17] | Track the electrical change response of fruit physiology and analyze their ripening | Cell wall and vacuole resistance decreased by 60% and 26%, respectively, and membrane capacitance decreased by 9%. | Fruit and vegetable quality measurement | System-dependent, which may limit functionality | Bera et al., 2017; Harker and maindonald, 1994 | |
| Electrical parameter (Z, R, Y, G), with simple linear regression analysis [19] | Determining the effect of Total Soluble Solids (TSS) on the electrical conductivity of reconstituted apple juice. | EIS parameters are good for determining TSS content. Rapid determination of the TSS content in different fruit juices, detection of their adulteration | Food quality measurement | Lack of changes in physicochemical qualities | Żywica and Banach, 2015. | |
| Sensor based on four electrodes and three indexes as indicators of leaf water content [20] | Develop water-saving agriculture and increase water-use efficiency | Negative correlation with all three parameters was observed. Relative water content showed the best correlation with the leaf property. | Crop production (leaf water content) | Existence of a severe fringe effect | Zheng et al., 2014. | |
| Four different models; CNLS method [24] | Determine the best electrical model for plant tissues analysis | Plant tissue conformed better to a double-shell model than others. | Plant tissue analysis | - | Zhang and Willison, 1991. | |
| * Finding R-C with DSM to understand plant physiology | Monitor and understand leaf physiology for 16 h using double-shell model parameters and a comparative analysis. | were obtained. The results confirmed previous studies in the literature of. Rapid changes in R1, R2, C3, and C5 were noticeable after water uptake. Possibility of detecting a plant with slow growth status. | Plant physiology for crop production and precision in agriculture. | System-dependent | Proposed |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nouaze, J.C.; Kim, J.H.; Jeon, G.R.; Kim, J.H. Monitoring of Indoor Farming of Lettuce Leaves for 16 Hours Using Electrical Impedance Spectroscopy (EIS) and Double-Shell Model (DSM). Sensors 2022, 22, 9671. https://doi.org/10.3390/s22249671
Nouaze JC, Kim JH, Jeon GR, Kim JH. Monitoring of Indoor Farming of Lettuce Leaves for 16 Hours Using Electrical Impedance Spectroscopy (EIS) and Double-Shell Model (DSM). Sensors. 2022; 22(24):9671. https://doi.org/10.3390/s22249671
Chicago/Turabian StyleNouaze, Joseph Christian, Jae Hyung Kim, Gye Rok Jeon, and Jae Ho Kim. 2022. "Monitoring of Indoor Farming of Lettuce Leaves for 16 Hours Using Electrical Impedance Spectroscopy (EIS) and Double-Shell Model (DSM)" Sensors 22, no. 24: 9671. https://doi.org/10.3390/s22249671