
Distinguishing Nanoparticle Aggregation from Viscosity Changes in MPS/MSB Detection of Biomarkers
 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Modeling the Fraction of Bound mNPs after Biomarker-Induced Aggregation
2.2. Simulations of mNP Rotations Using the Effective Field Model
2.3. Dimer-Only Model
2.4. In Vitro MPS/MSB Apparatus
2.5. Signal Scaling
2.6. In Vitro Reagents
2.7. Numerical Solution
3. Results
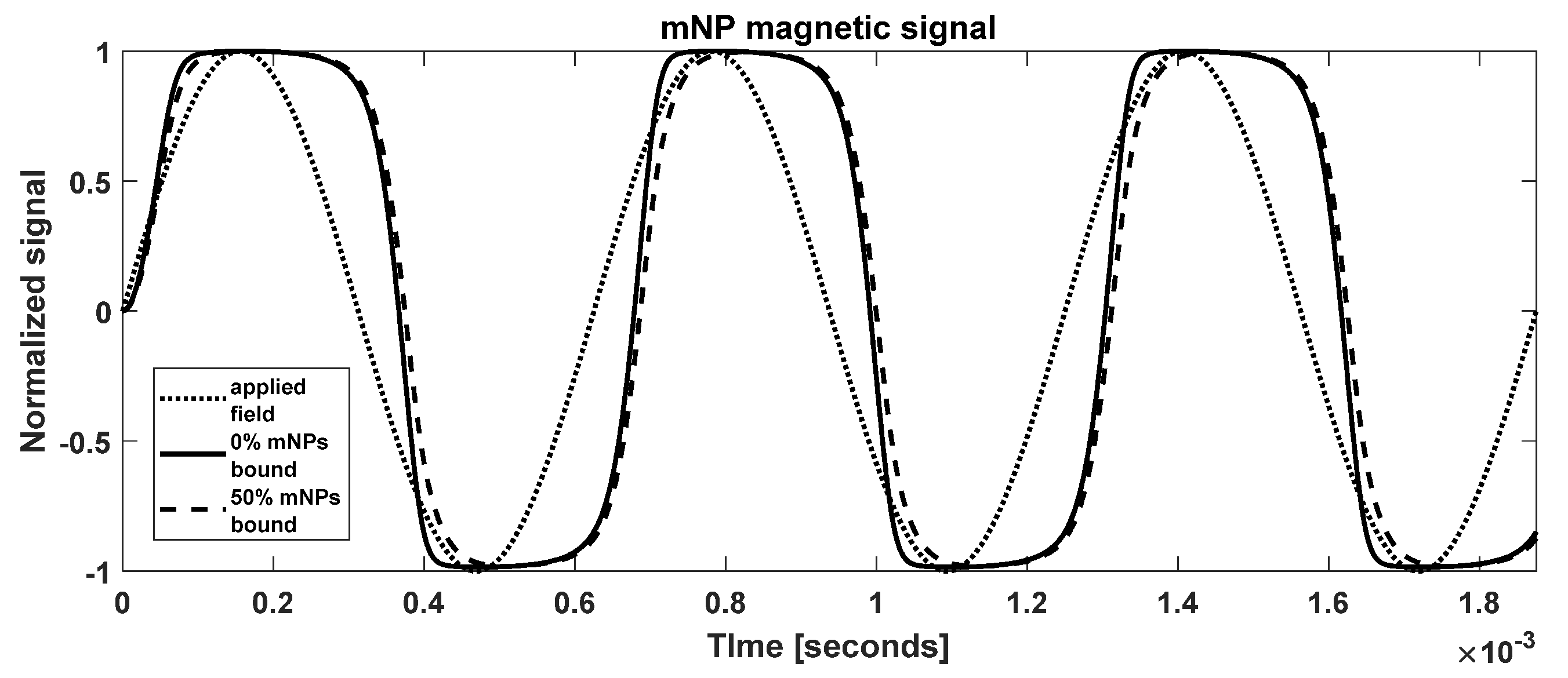
3.1. MNP Magnetic Signal
3.2. Biomarker-Induced mNP Aggregation
3.3. Ratio of Harmonics and Scaling
3.4. Aggregation Versus Viscosity Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gleich, B.; Weizenecker, J. Tomographic Imaging Using the Nonlinear Response of Magnetic Particles. Nature 2005, 435, 1214–1217. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.L.C.; Zhang, X.Y.; Steinberg, X.G.; Qu, X.H.; Huang, X.S.; Cheng, X.M.; Bliss, X.T.; Du, X.F.; Rao, X.J.; Song, X.G.; et al. A Review of Magnetic Particle Imaging and Perspectives on Neuroimaging. Am. J. Neuroradiol. 2019, 40, 206–212. [Google Scholar] [CrossRef]
- Knopp, T.; Buzug, T.M. Magnetic Particle Imaging: An Introduction to Imaging Principles and Scanner Instrumentation; Springer: Berlin/Heidelberg, Germany, 2012; ISBN 9783642041990. [Google Scholar]
- Talebloo, N.; Gudi, M.; Robertson, N.; Wang, P. Magnetic Particle Imaging: Current Applications in Biomedical Research. J. Magn. Reson. Imaging 2020, 51, 1659–1668. [Google Scholar] [CrossRef] [PubMed]
- Graeser, M.; Thieben, F.; Szwargulski, P.; Werner, F.; Gdaniec, N.; Boberg, M.; Griese, F.; Möddel, M.; Ludewig, P.; Van De Ven, D.; et al. Human-Sized Magnetic Particle Imaging for Brain Applications. Nat. Commun. 2019, 10, 1936. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Li, J.; Peng, P.; Nassab, N.H.; Smith, B.R. Quantitative Drug Release Monitoring in Tumors of Living Subjects by Magnetic Particle Imaging Nanocomposite. Nano Lett. 2019, 19, 6725–6733. [Google Scholar] [CrossRef] [PubMed]
- Herz, S.; Vogel, P.; Kampf, T.; Dietrich, P.; Veldhoen, S.; Rückert, M.A.; Kickuth, R.; Behr, V.C.; Bley, T.A. Magnetic Particle Imaging–Guided Stenting. J. Endovasc. Ther. 2019, 26, 512–519. [Google Scholar] [CrossRef]
- Wu, K.; Su, D.; Liu, J.; Saha, R.; Wang, J.-P. Magnetic Nanoparticles in Nanomedicine: A Review of Recent Advances. Nanotechnology 2019, 30, 502003. [Google Scholar] [CrossRef]
- Rauwerdink, A.M.; Giustini, A.J.; Weaver, J.B. Simultaneous Quantification of Multiple Magnetic Nanoparticles. Nanotechnology 2010, 21, 45. [Google Scholar] [CrossRef]
- Zhang, X.; Reeves, D.; Shi, Y.; Gimi, B.; Nemani, K.; Perreard, I.M.; Toraya-Brown, S.; Fiering, S.; Weaver, J.B. Toward Localized In Vivo Biomarker Concentration Measurements. IEEE Trans. Magn. 2015, 51, 1–4. [Google Scholar] [CrossRef]
- Shi, Y.; Jyoti, D.; Gordon-Wylie, S.W.; Weaver, J.B. Quantification of Magnetic Nanoparticles by Compensating for Multiple Environment Changes Simultaneously. Nanoscale 2020, 12, 195–200. [Google Scholar] [CrossRef]
- Rauwerdink, A.M.; Hansen, E.W.; Weaver, J.B. Nanoparticle Temperature Estimation in Combined Ac and Dc Magnetic Fields. Phys. Med. Biol. 2009, 54, L51–L55. [Google Scholar] [CrossRef] [PubMed]
- Rauwerdink, A.; Weaver, J. Viscous Effects on Nanoparticle Magnetization Harmonics. J. Magn. Magn. Mater. 2010, 322, 609–613. [Google Scholar] [CrossRef]
- Wu, K.; Liu, J.; Wang, Y.; Ye, C.; Feng, Y.; Wang, J.-P. Superparamagnetic Nanoparticle-Based Viscosity Test. Appl. Phys. Lett. 2015, 107, 53701. [Google Scholar] [CrossRef]
- Weaver, J.B.; Rauwerdink, A.M.; Perreard, I.; Kilfoyle, R. Micro-rheology: Evaluating the rigidity of the microenvironment surrounding antibody binding sites. In Proceedings of the Medical Imaging 2010: Biomedical Applications in Molecular, Structural, and Functional Imaging, San Diego, CA, USA, 1 March 2010; Volume 7626, p. 76261J. [Google Scholar]
- Weaver, J.B.; Rauwerdink, K.M.; Rauwerdink, A.M.; Perreard, I.M. Magnetic Spectroscopy of Nanoparticle Brownian Motion Measurement of Microenvironment Matrix Rigidity. Biomed. Tech. Eng. 2013, 58, 547–550. [Google Scholar] [CrossRef] [PubMed]
- Weaver, J.; Zhang, X.; Toraya-Brown, S.; Reeves, D.; Perreard, I.; Fiering, S. TU-G-217A-07: Magnetic Nanoparticle Quantitation: Compensating for Relaxation Effects. Med. Phys. 2012, 39, 3927. [Google Scholar] [CrossRef]
- Shi, Y.; Weaver, J.B. Concurrent Quantification of Magnetic Nanoparticles Temperature and Relaxation Time. Med. Phys. 2019, 46, 4070–4076. [Google Scholar] [CrossRef]
- Gordon-Wylie, S.W.; Grüttner, C.; Teller, H.; Weaver, J.B. Using Magnetic Nanoparticles and Protein–Protein Interactions to Measure PH at the Nanoscale. IEEE Sensors Lett. 2020, 4, 1–3. [Google Scholar] [CrossRef]
- Weaver, J.B.; Shi, Y.; Ness, D.B.; Khurshid, H.; Samia, A.C.S. Sensitivity Limits for in Vivo ELISA Measurements of Molecular Biomarker Concentrations. Int. J. Magn. Part. Imaging 2017, 3, 1706003. [Google Scholar] [CrossRef]
- Perreard, I.M.; Reeves, D.B.; Zhang, X.; Weaver, J.B. Magnetic Nanoparticles Temperature Measurements. In Proceedings of the 2013 International Workshop on Magnetic Particle Imaging (IWMPI), Berkeley, CA, USA, 23–24 March 2013; IEEE: New York, NY, USA, 2013; p. 1. [Google Scholar]
- Engvall, E.; Perlmann, P. Enzyme-Linked Immunosorbent Assay, Elisa. J. Immunol. 1972, 109, 129–135. [Google Scholar]
- Ogawa, M.; Kosaka, N.; Choyke, P.L.; Kobayashi, H. H-Type Dimer Formation of Fluorophores: A Mechanism for Activatable, in Vivo Optical Molecular Imaging. ACS Chem. Biol. 2009, 4, 535–546. [Google Scholar] [CrossRef]
- Jones, K.M.; Pollard, A.C.; Pagel, M.D. Clinical Applications of Chemical Exchange Saturation Transfer (CEST) MRI. J. Magn. Reson. Imaging 2018, 47, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Ward, K.M.; Aletras, A.H.; Balaban, R.S. A New Class of Contrast Agents for MRI Based on Proton Chemical Exchange Dependent Saturation Transfer (CEST). J. Magn. Reson. 2000, 143, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-J.; Yang, J.-Y.; Shu, T.-Y.; Lin, T.-Y.; Chen, Y.-Y.; Su, M.-Y.; Li, W.-J.; Liu, M.-Y. Detection of C-Reactive Protein Based on Magnetic Nanoparticles and Capillary Zone Electrophoresis with Laser-Induced Fluorescence Detection. J. Chromatogr. A 2013, 1315, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Heneka, M.T.; Golenbock, D.T.; Latz, E. Innate Immunity in Alzheimer’s Disease. Nat. Immunol. 2015, 16, 229–236. [Google Scholar] [CrossRef]
- Vezzani, A.; Aronica, E.; Mazarati, A.; Pittman, Q.J. Epilepsy and Brain Inflammation. Exp. Neurol. 2013, 244, 11–21. [Google Scholar] [CrossRef]
- Na, K.-S.; Jung, H.-Y.; Kim, Y.-K. The Role of Pro-Inflammatory Cytokines in the Neuroinflammation and Neurogenesis of Schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 48, 277–286. [Google Scholar] [CrossRef]
- Tucsek, Z.; Toth, P.; Sosnowska, D.; Gautam, T.; Mitschelen, M.; Koller, A.; Szalai, G.; Sonntag, W.E.; Ungvari, Z.; Csiszar, A. Obesity in Aging Exacerbates Blood–Brain Barrier Disruption, Neuroinflammation, and Oxidative Stress in the Mouse Hippocampus: Effects on Expression of Genes Involved in Beta-Amyloid Generation and Alzheimer’s Disease. J. Gerontol. Ser. A Biomed. Sci. Med. Sci. 2014, 69, 1212–1226. [Google Scholar] [CrossRef]
- Saleh, A.; Schroeter, M.; Jonkmanns, C.; Hartung, H.; MoÈdder, U.; Jander, S. In Vivo MRI of Brain Inflammation in Human Ischaemic Stroke. Brain 2004, 127, 1670–1677. [Google Scholar] [CrossRef]
- Kim, H.-W.; Rapoport, S.I.; Rao, J.S. Altered Expression of Apoptotic Factors and Synaptic Markers in Postmortem Brain from Bipolar Disorder Patients. Neurobiol. Dis. 2010, 37, 596–603. [Google Scholar] [CrossRef]
- Fenn, A.M.; Gensel, J.C.; Huang, Y.; Popovich, P.G.; Lifshitz, J.; Godbout, J.P. Immune Activation Promotes Depression 1 Month after Diffuse Brain Injury: A Role for Primed Microglia. Biol. Psychiatry 2014, 76, 575–584. [Google Scholar] [CrossRef]
- Atmuri, A.K.; Henson, M.A.; Bhatia, S.R. A Population Balance Equation Model to Predict Regimes of Controlled Nanoparticle Aggregation. Colloids Surf. A Physicochem. Eng. Asp. 2013, 436, 325–332. [Google Scholar] [CrossRef]
- Łepek, M. How initial condition impacts aggregation—A systematic numerical study. arXiv 2021, arXiv:2112.14828. [Google Scholar]
- Grammatikopoulos, P.; Sowwan, M.; Kioseoglou, J. Computational Modeling of Nanoparticle Coalescence. Adv. Theory Simul. 2019, 2, 1900013. [Google Scholar] [CrossRef]
- Basu, S.; Ghosh, S.K.; Kundu, S.; Panigrahi, S.; Praharaj, S.; Pande, S.; Jana, S.; Pal, T. Biomolecule Induced Nanoparticle Aggregation: Effect of Particle Size on Interparticle Coupling. J. Colloid Interface Sci. 2007, 313, 724–734. [Google Scholar] [CrossRef]
- Khavani, M.; Izadyar, M.; Housaindokht, M.R. Modeling of the Functionalized Gold Nanoparticle Aggregation in the Presence of Dopamine: A Joint MD/QM Study. J. Phys. Chem. C 2018, 122, 26130–26141. [Google Scholar] [CrossRef]
- Karvelas, E.G.; Lampropoulos, N.K.; Benos, L.T.; Karakasidis, T.; Sarris, I.E. On the Magnetic Aggregation of Fe3O4 Nanoparticles. Comput. Methods Programs Biomed. 2021, 198, 105778. [Google Scholar] [CrossRef]
- Mejías, R.; Hernández Flores, P.; Talelli, M.; Tajada-Herráiz, J.L.; Brollo, M.E.F.; Portilla, Y.; Morales, M.P.; Barber, D.F. Cell-Promoted Nanoparticle Aggregation Decreases Nanoparticle-Induced Hyperthermia under an Alternating Magnetic Field Independently of Nanoparticle Coating, Core Size, and Subcellular Localization. ACS Appl. Mater. Interfaces 2019, 11, 340–355. [Google Scholar] [CrossRef] [PubMed]
- Abu-Bakr, A.F.; Zubarev, A.Y. Effect of Ferromagnetic Nanoparticles Aggregation on Magnetic Hyperthermia. Eur. Phys. J. Spec. Top. 2020, 229, 323–329. [Google Scholar] [CrossRef]
- Bélteky, P.; Rónavári, A.; Zakupszky, D.; Boka, E.; Igaz, N.; Szerencsés, B.; Pfeiffer, I.; Vágvölgyi, C.; Kiricsi, M.; Kónya, Z. Are Smaller Nanoparticles Always Better? Understanding the Biological Effect of Size-Dependent Silver Nanoparticle Aggregation Under Biorelevant Conditions. Int. J. Nanomed. 2021, 16, 3021–3040. [Google Scholar] [CrossRef]
- Yoshimura, K.; Patmawati; Maeda, M.; Kamiya, N.; Zako, T. Protein-Functionalized Gold Nanoparticles for Antibody Detection Using the Darkfield Microscopic Observation of Nanoparticle Aggregation. Anal. Sci. 2021, 37, 507–511. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Su, D.; Saha, R.; Wong, D.; Wang, J.-P. Magnetic Particle Spectroscopy-Based Bioassays: Methods, Applications, Advances, and Future Opportunities. J. Phys. D Appl. Phys. 2019, 52, 173001. [Google Scholar] [CrossRef]
- Smoluchowski, M.V. Drei Vortrage Uber Diffusion, Brownsche Bewegung Und Koagulation von Kolloidteilchen. Z. Fur Phys. 1916, 17, 557–585. [Google Scholar]
- Reeves, D.B.; Weizenecker, J.; Weaver, J.B. Langevin Equation Simulation of Brownian Magnetic Nanoparticles with Experimental and Model Comparisons. In Proceedings of the Medical Imaging 2013: Biomedical Applications in Molecular, Structural, and Functional Imaging; International Society for Optics and Photonics, Lake Buena Vista, Fl, USA, 29 March 2013; Volume 8672, p. 86721C. [Google Scholar]
- Cohen, A. A Pad6 Approximant to the Inverse Langevin Function. Rheol. Acta 1991, 30, 270–273. [Google Scholar] [CrossRef]
- Ahrentorp, F.; Astalan, A.; Blomgren, J.; Jonasson, C.; Wetterskog, E.; Svedlindh, P.; Lak, A.; Ludwig, F.; Van IJzendoorn, L.J.; Westphal, F.; et al. Effective Particle Magnetic Moment of Multi-Core Particles. J. Magn. Magn. Mater. 2015, 380, 221–226. [Google Scholar] [CrossRef]
- Gordon-Wylie, S.W.; Ness, D.B.; Shi, Y.; Mirza, S.K.; Paulsen, K.D.; Weaver, J.B. Measuring Protein Biomarker Concentrations Using Antibody Tagged Magnetic Nanoparticles. Biomed. Phys. Eng. Express 2020, 6, 065025. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Conclusion |
|---|---|
| MRI-guided NP drug delivery [39] | Fe3O4 NPs aggregate with stronger fields. |
| Cancer hyperthermia treatment [40,41] | Aggregation reduces heating. |
| Toxicity due to Ag NP aggregation [42] | Larger Ag NPs aggregate less. |
| Functionalized Au NPs [43] | Antibody detection with microscopy. |
| Bioassays with mag. spectroscopy [44] | In vitro detection of several biomarkers. |
| Parameter | Value | Parameter | Value |
|---|---|---|---|
| mNP effective core diameter | 15–25 nm | Temperature, T | 31 °C |
| mNP hydrodynamic dia. | 100 nm | Viscosity, η | 0.78–1.17 mPa.s |
| Applied oscillating field, H | 10 mT | Fe concentration | <5 µg/ml |
| Applied field frequencies, ω | 0.4–2 kHz | mNP magnetization | 49 Am2/kg Fe |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jyoti, D.; Gordon-Wylie, S.W.; Reeves, D.B.; Paulsen, K.D.; Weaver, J.B. Distinguishing Nanoparticle Aggregation from Viscosity Changes in MPS/MSB Detection of Biomarkers. Sensors 2022, 22, 6690. https://doi.org/10.3390/s22176690
Jyoti D, Gordon-Wylie SW, Reeves DB, Paulsen KD, Weaver JB. Distinguishing Nanoparticle Aggregation from Viscosity Changes in MPS/MSB Detection of Biomarkers. Sensors. 2022; 22(17):6690. https://doi.org/10.3390/s22176690
Chicago/Turabian StyleJyoti, Dhrubo, Scott W. Gordon-Wylie, Daniel B. Reeves, Keith D. Paulsen, and John B. Weaver. 2022. "Distinguishing Nanoparticle Aggregation from Viscosity Changes in MPS/MSB Detection of Biomarkers" Sensors 22, no. 17: 6690. https://doi.org/10.3390/s22176690