
In Situ Measurements of Plankton Biorhythms Using Submersible Holographic Camera
 , , , and
, , , and
Abstract
:1. Introduction
2. Problem Statement
- -
- Non-invasiveness (in terms of affecting the ecosystem) of obtaining information, which necessarily excludes the use of net catching methods;
- -
- Long-term (multiday) monitoring of representative plankton samples for the creation of time series with regular time intervals (requirement of further spectral analysis);
- -
- Parallel registration of plankton vital signs and changes in hydrological characteristics of the medium in order to detect their synchronization (asynchronization) due to adverse changes.
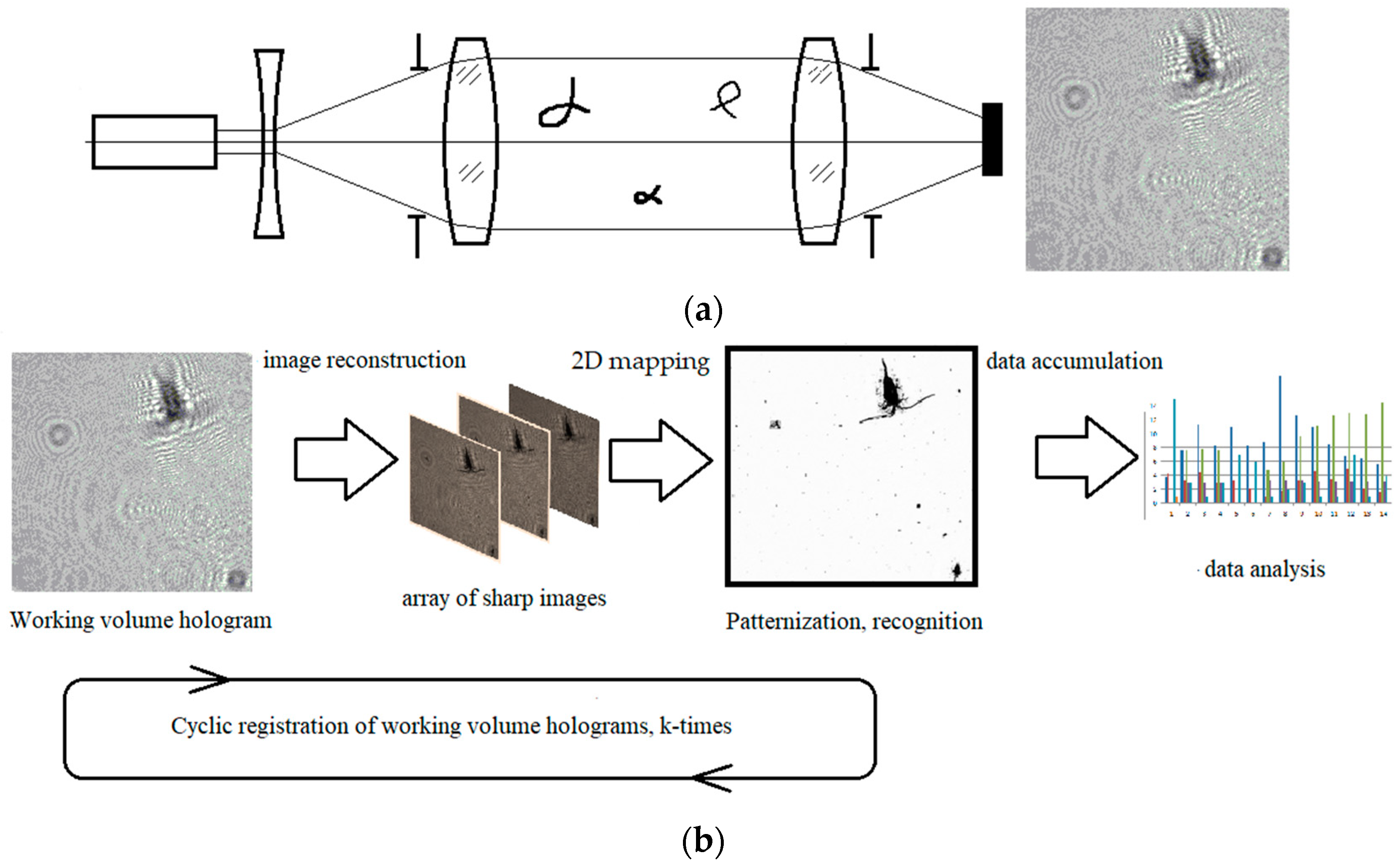
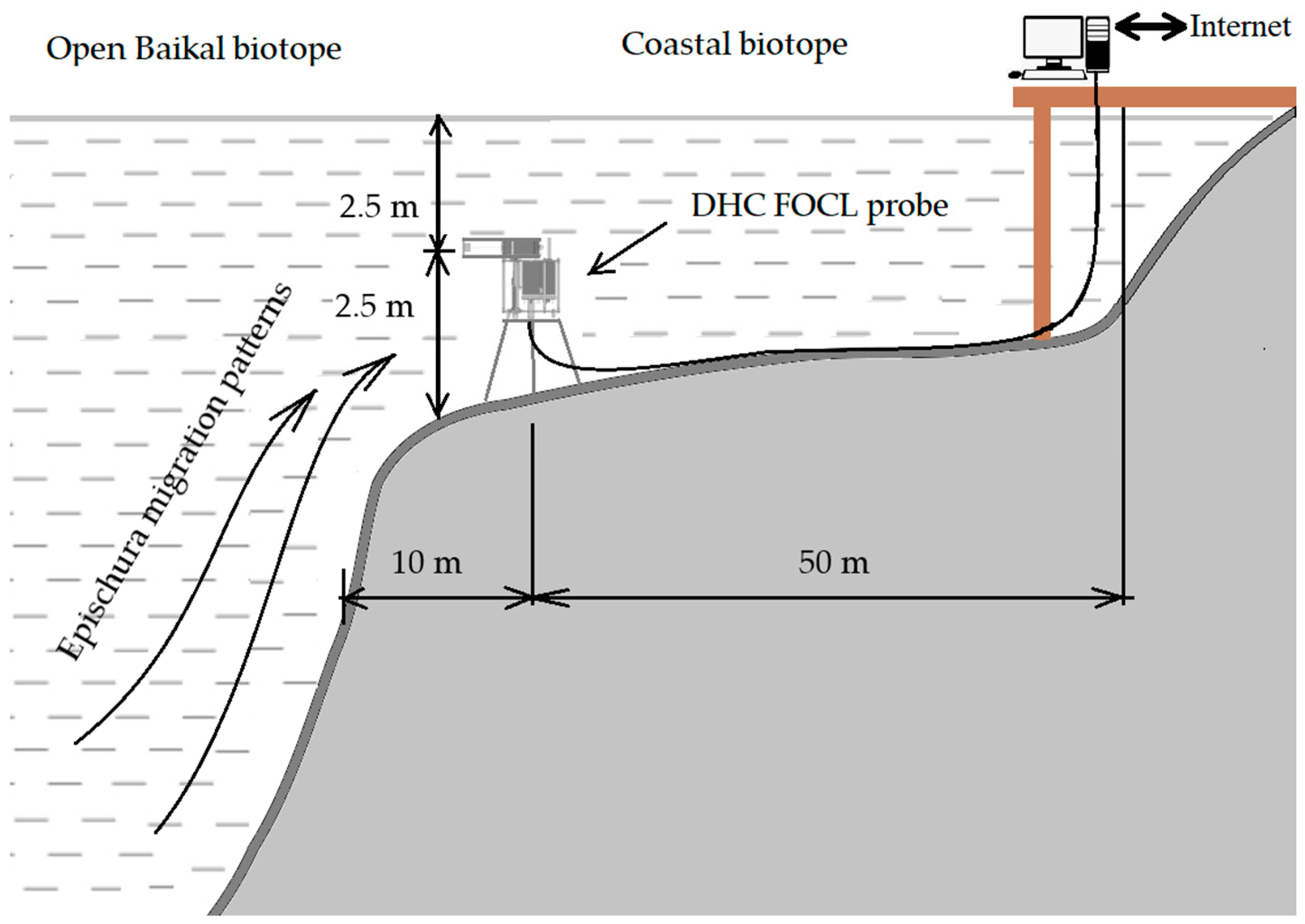
3. Equipment and Methods
- Plankton concentration;
- Distribution of plankton by main taxa;
- Average size and dispersion of sizes of individuals;
- Particle size distribution;
- Average size of individuals and size dispersion within main taxa;
- Particle size distribution within main taxa;
- Water turbidity;
- Suspension statistics (histogram by non-living particle size).
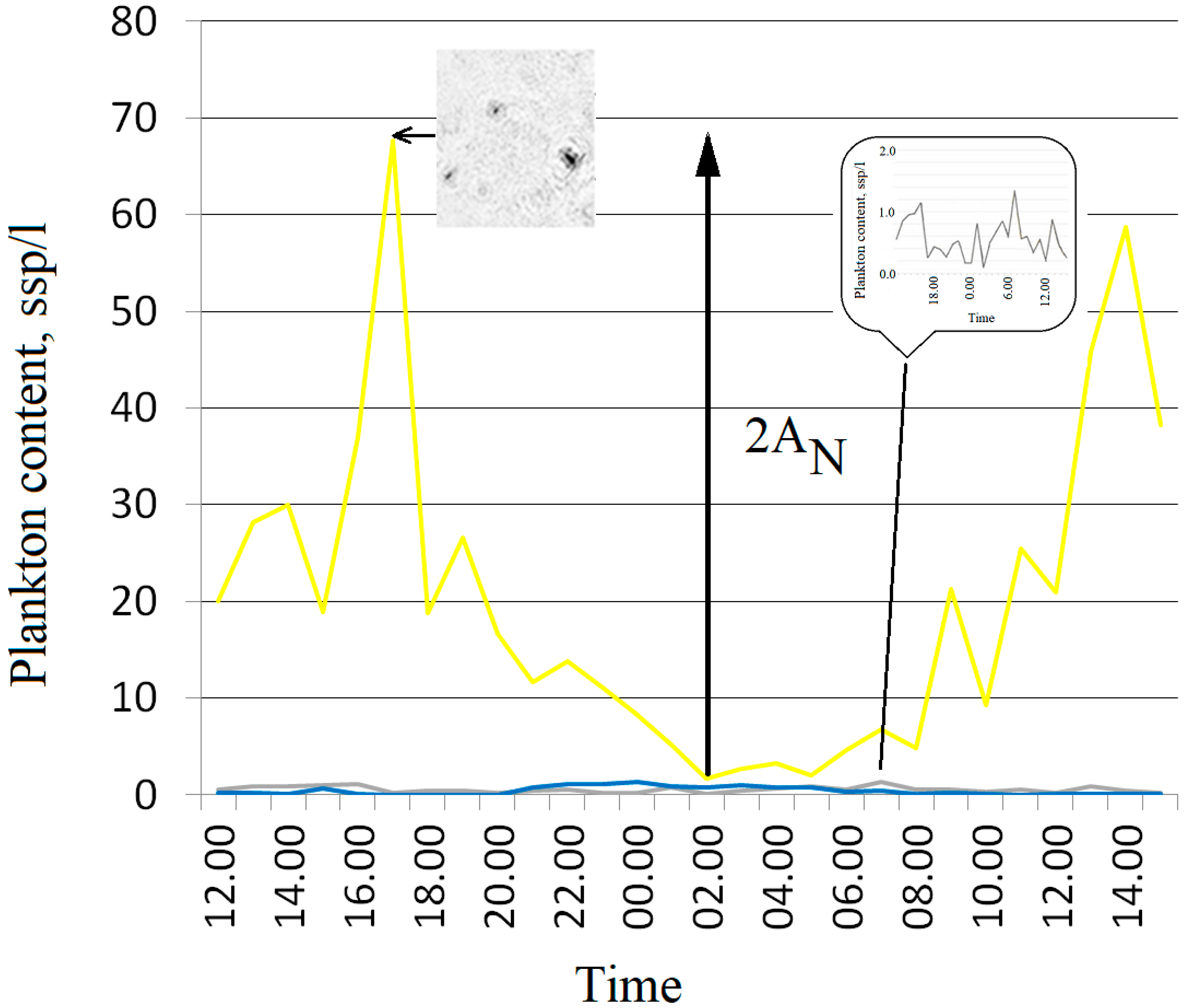
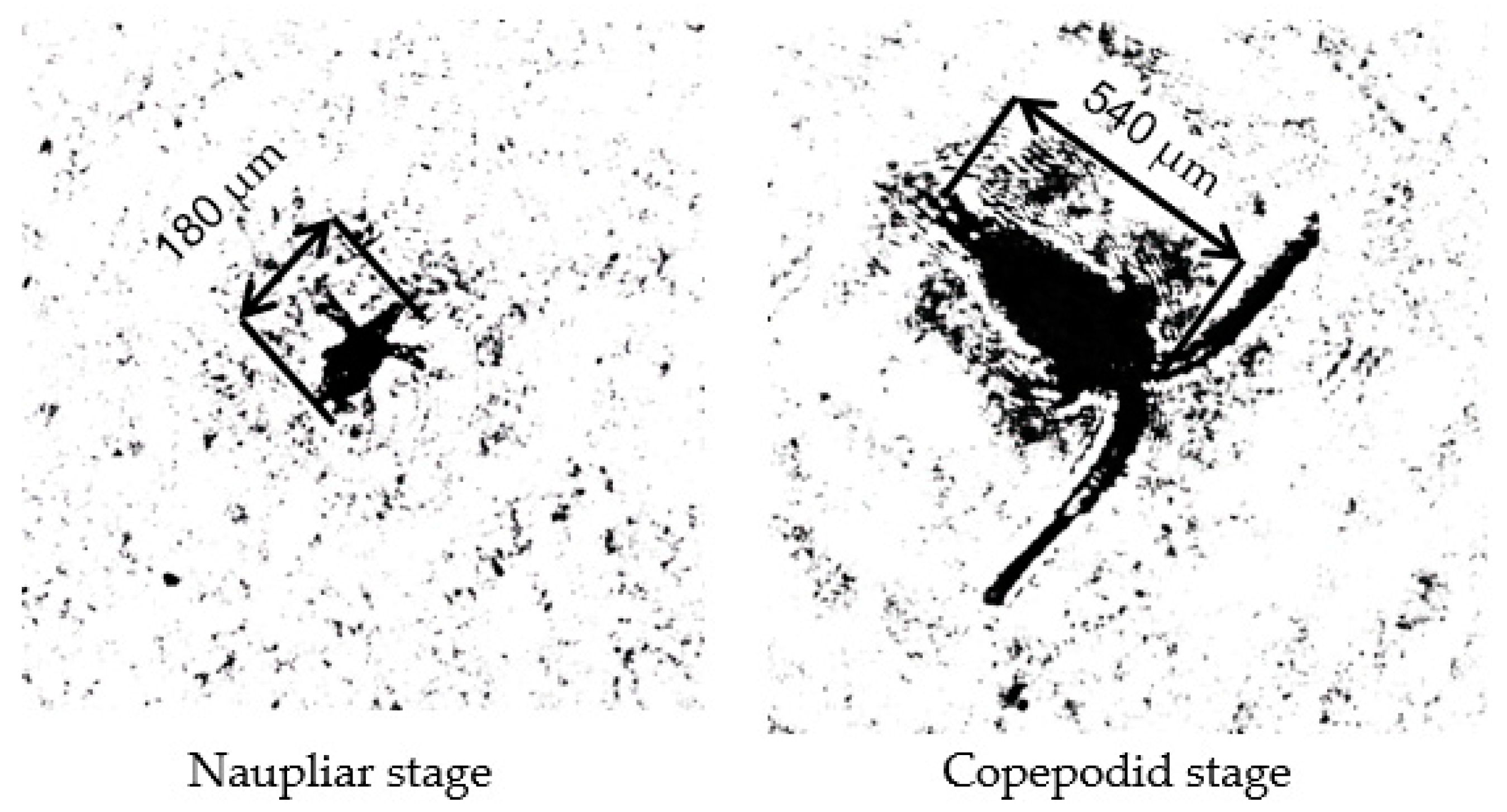
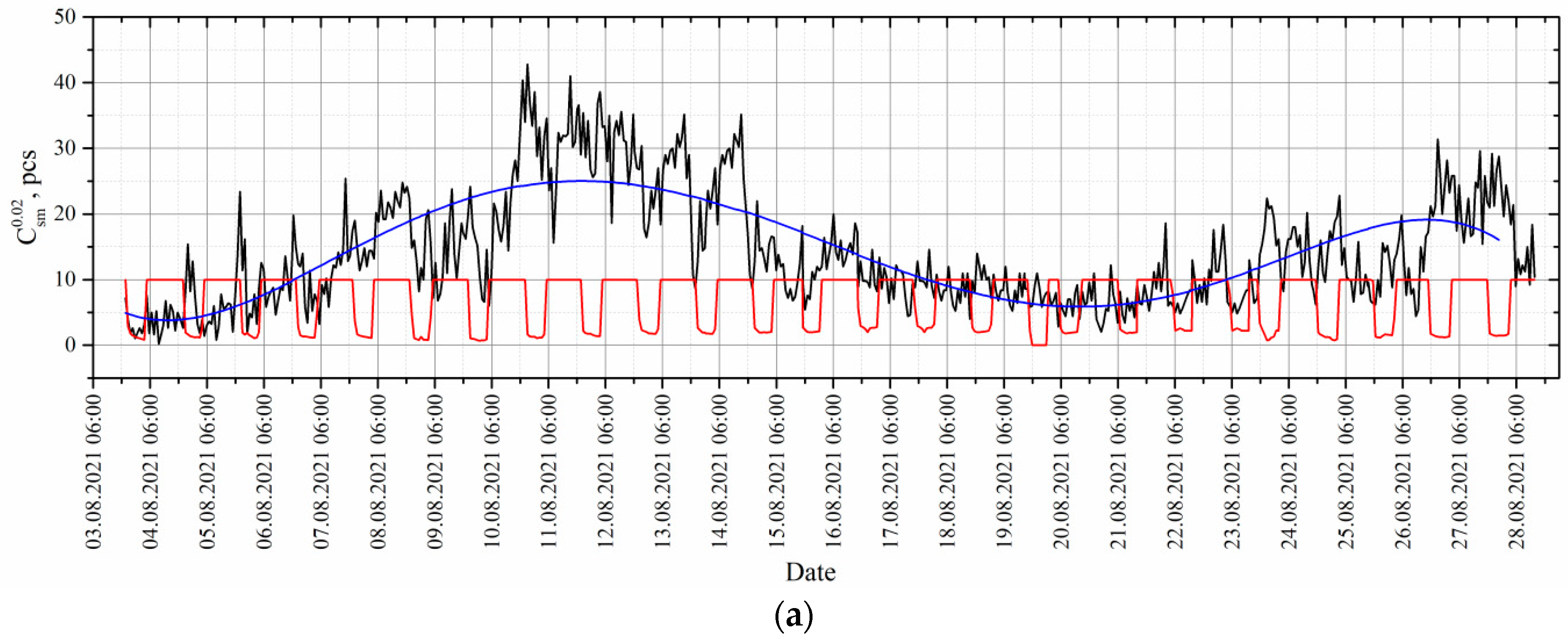
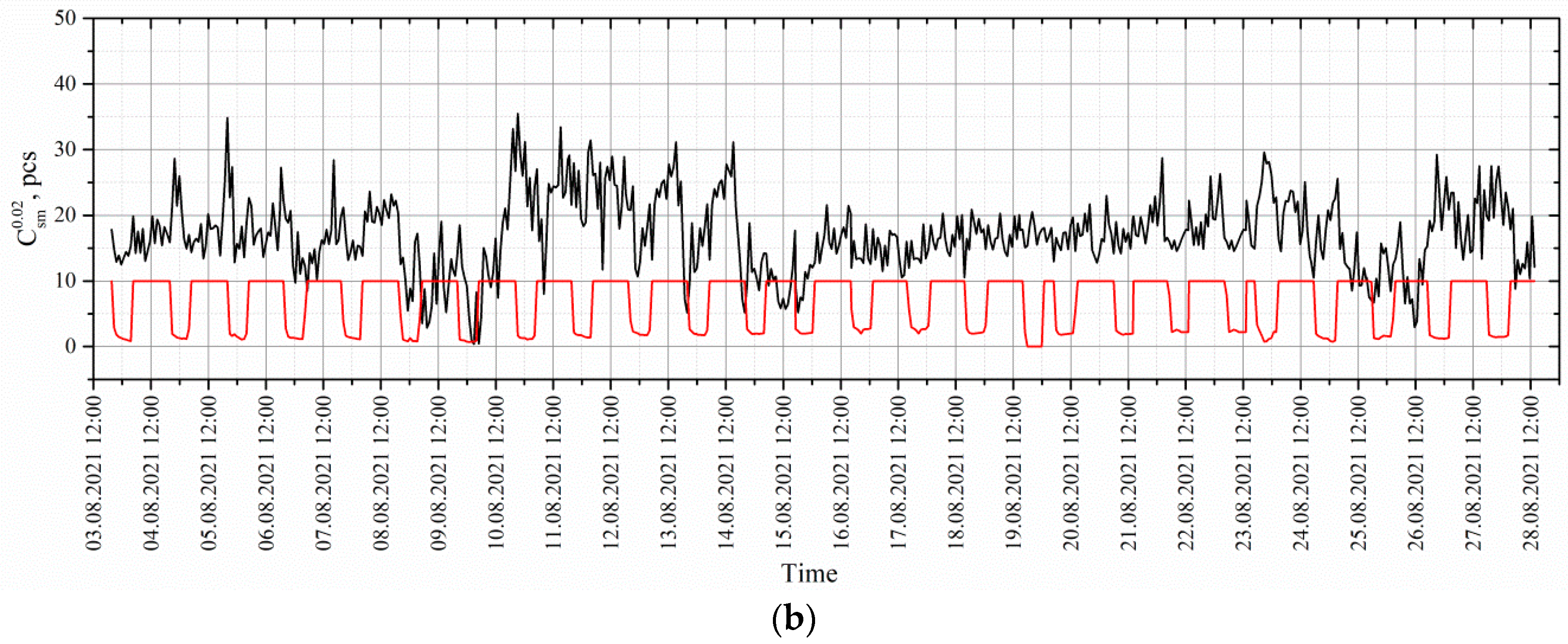
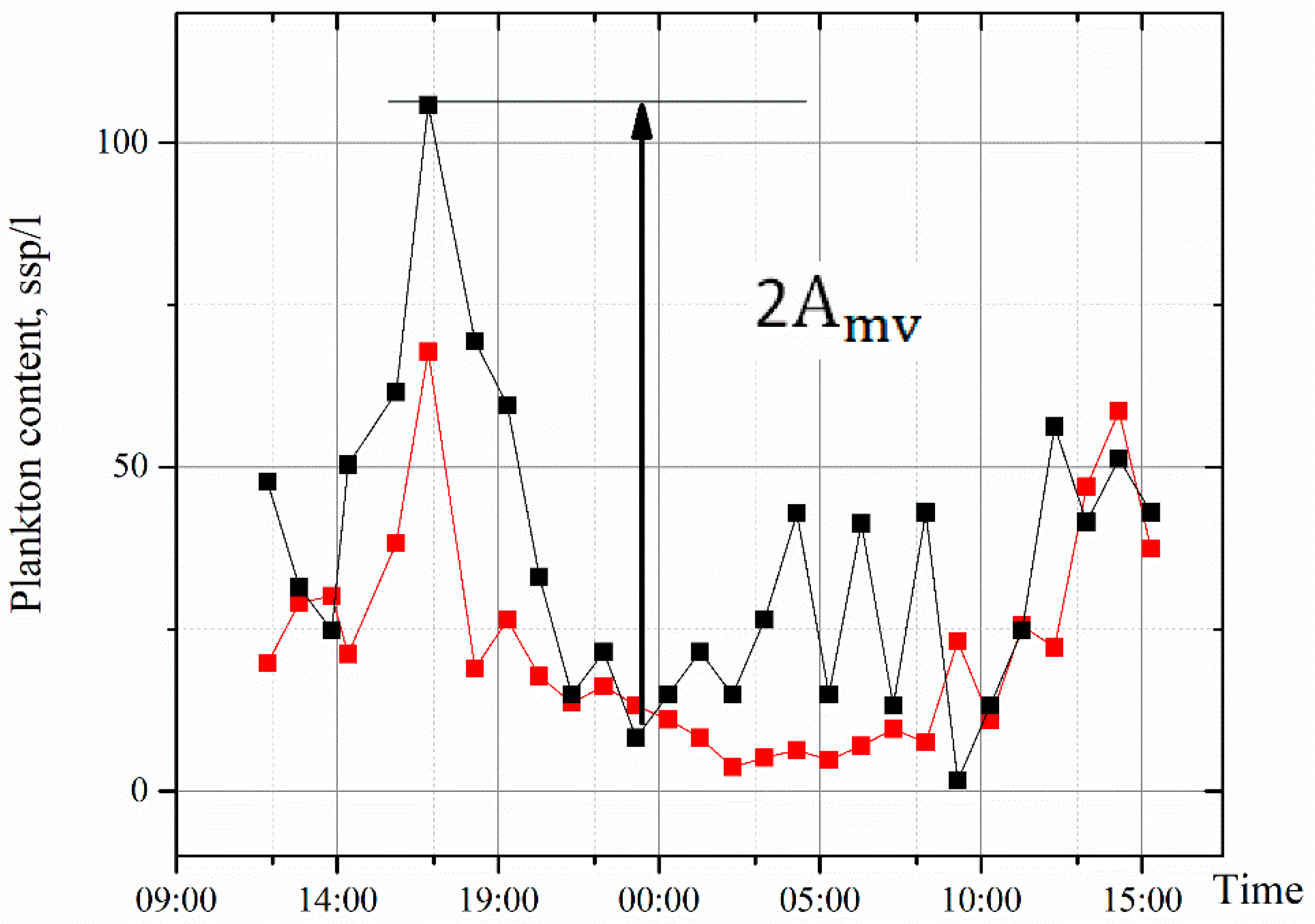
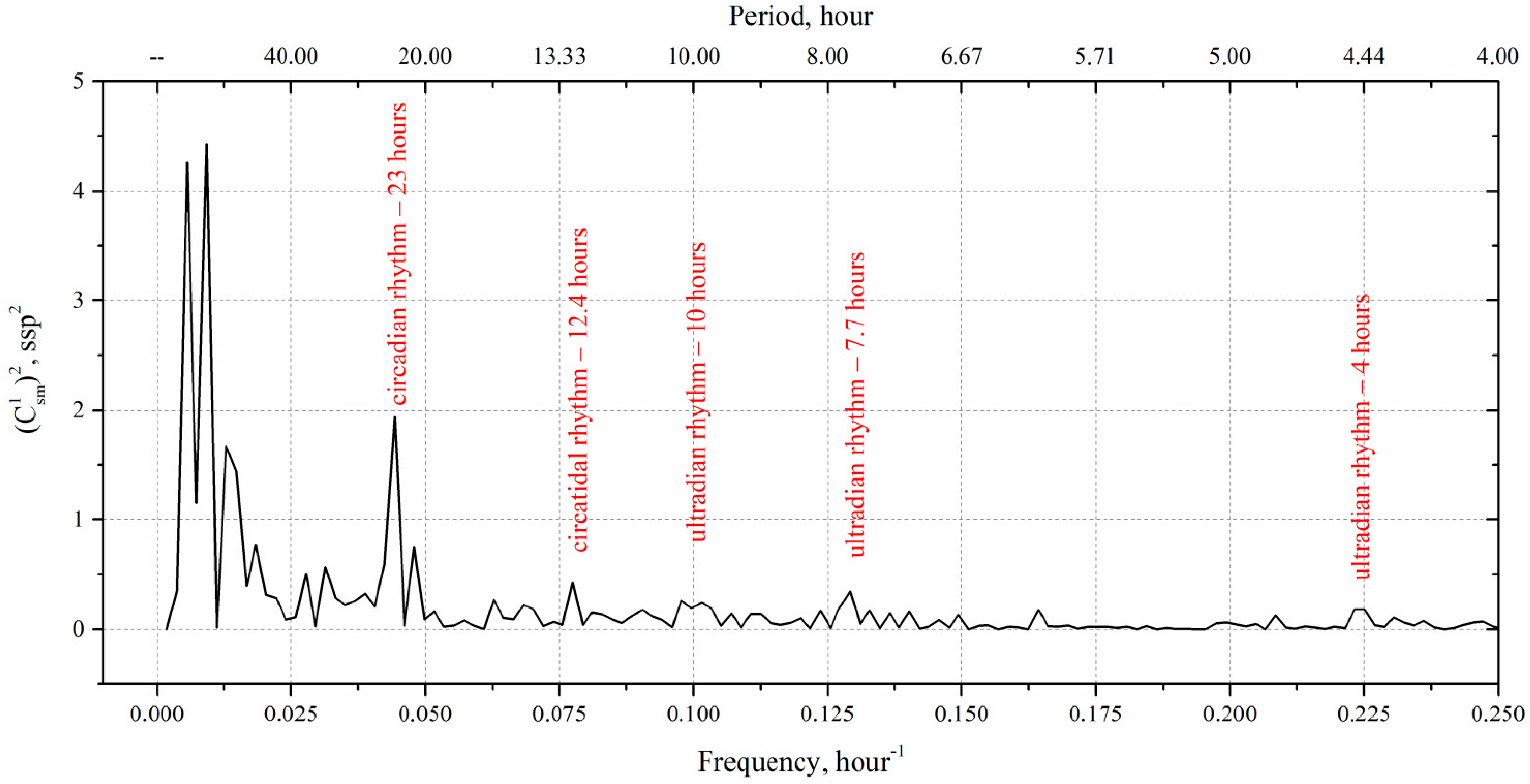
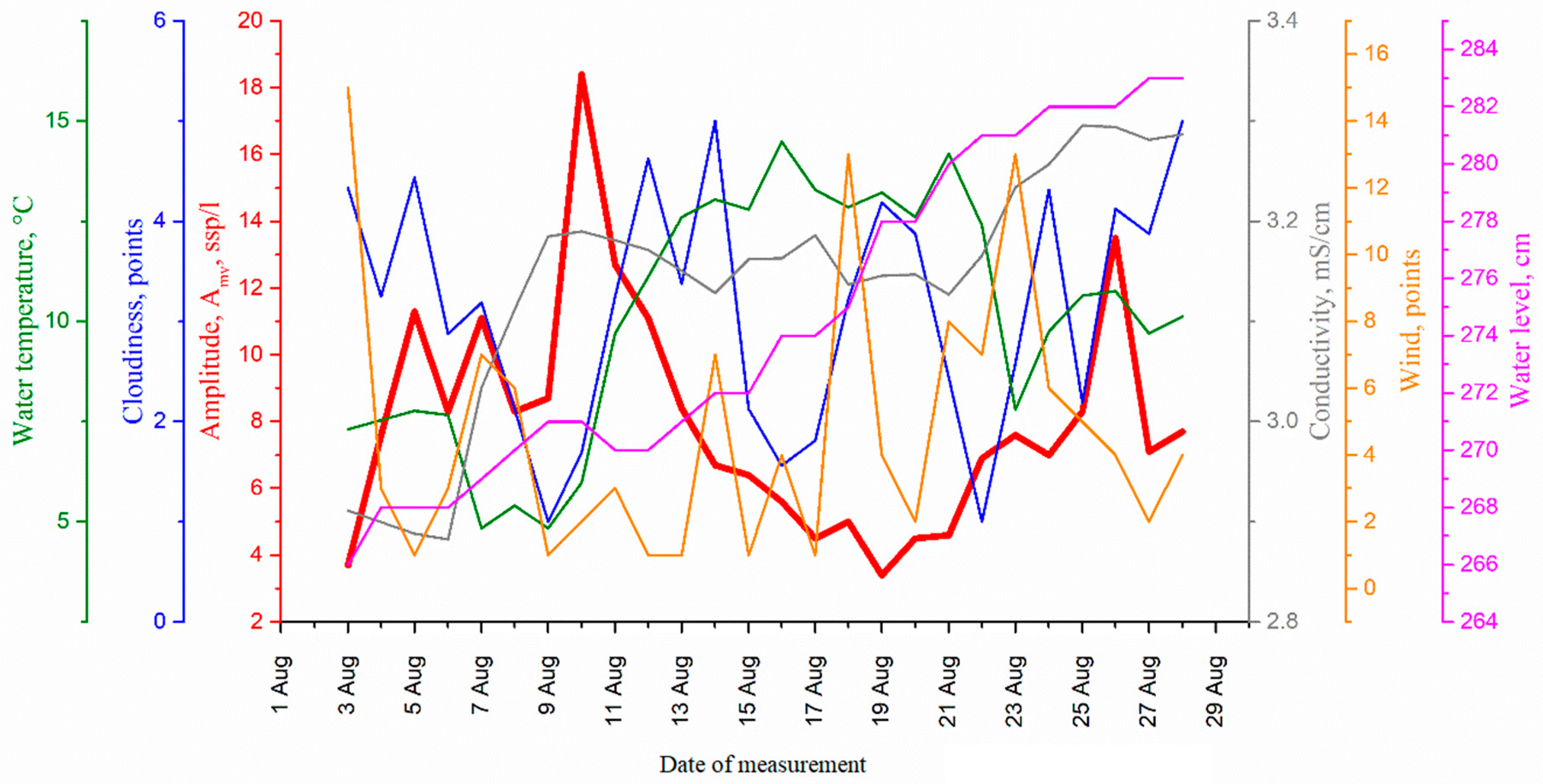
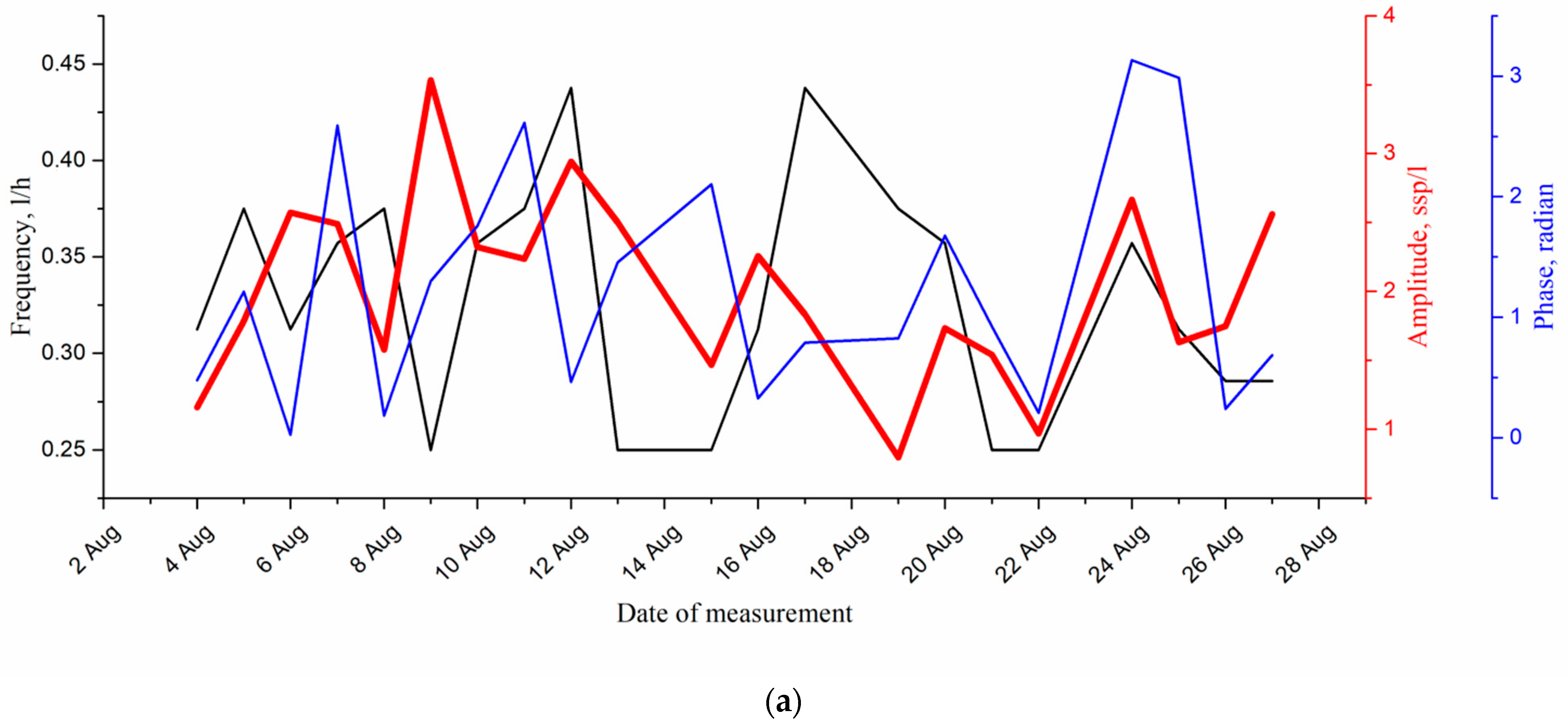
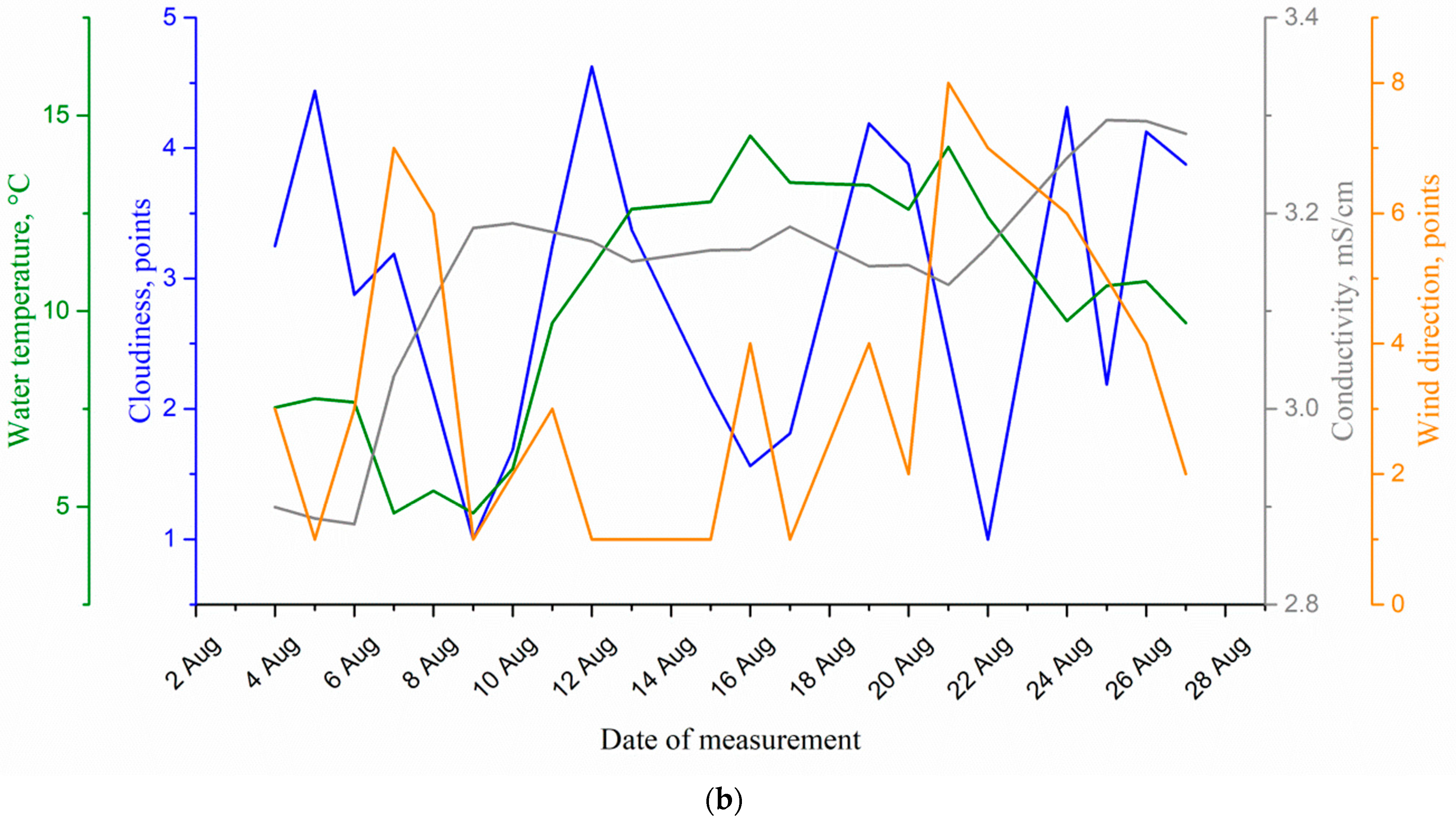
4. Results and Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Sunagawa, S.; Coelho, L.P.; Chaffron, S.; Kultima, J.R.; Labadie, K.; Salazar, G.; Djahanschiri, B.; Zeller, G.; Mende, D.R.; Alberti, A.; et al. Ocean plankton. Structure and function of the global ocean microbiome. Science 2015, 348, 1261359. [Google Scholar] [CrossRef] [PubMed]
- Bairey, E.; Kelsic, E.D.; Kishony, R. High-order species interactions shape ecosystem diversity. Nat. Commun. 2016, 7, 12285. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, D.; Fiegna, F.; Behrend, V.; Bundy, J.G.; Phillimore, A.B.; Bell, T.; Barraclough, T.G. Species interactions alter evolutionary responses to a novel environment. PLoS Biol. 2012, 10, e1001330. [Google Scholar] [CrossRef] [PubMed]
- Beaugrand, G.; Kirby, R.R. How Do Marine Pelagic Species Respond to Climate Change? Theories and Observations. Annu. Rev. Mar. Sci. 2018, 10, 169–197. [Google Scholar] [CrossRef]
- Beaugrand, G.; Conversi, A.; Atkinson, A.; Cloern, J.; Chiba, S.; Fonda-Umani, S.; Kirby, R.R.; Greene, C.H.; Goberville, E.; Otto, S.A.; et al. Prediction of unprecedented biological shifts in the global ocean. Nat. Clim. Chang. 2019, 9, 237–243. [Google Scholar] [CrossRef]
- Barton, A.D.; Irwin, A.J.; Finkel, Z.V.; Stock, C.A. Anthropogenic climate change drives shift and shuffle in North Atlantic phytoplankton communities. Proc. Natl. Acad. Sci. USA 2016, 113, 2964–2969. [Google Scholar] [CrossRef]
- Hutchins, D.A.; Fu, F. Microorganisms and ocean global change. Nat. Microbiol. 2017, 2, 17058. [Google Scholar] [CrossRef]
- Pittendrigh, C.S. Circadian Rhythms and the Circadian Organization of Living Systems. Cold Spring Harb. Symp. Quant. Biol. 1960, 25, 159–184. [Google Scholar] [CrossRef]
- Soviadan, Y.D.; Benedetti, F.; Brandão, M.C.; Ayata, S.-D.; Irisson, J.-O.; Jamet, J.L.; Kiko, R.; Lombard, F.; Gnandi, K.; Stemmann, L. Patterns of Mesozooplankton Community Composition and Vertical Fluxes in the Global Ocean. Ecology 2021, 200, 102717. [Google Scholar] [CrossRef]
- Aschoff, J. A Survey on Biological Rhythms. In Biological Rhythms; Aschoff, J., Ed.; Springer: Boston, MA, USA, 1981; pp. 3–10. ISBN 978-1-4615-6554-3. [Google Scholar]
- How Road Salt and Other Environmental Pollution Disrupts the Circadian Rhythm. Available online: https://www.chronobiology.com/how-road-salt-and-other-environmental-pollution-disrupts-the-circadian-rhythm/ (accessed on 1 June 2022).
- Walcutt, N.L.; Knörlein, B.; Sgouros, T.; Cetinić, I.; Omand, M.M. Virtual Reality and Oceanography: Overview, Applications, and Perspective. Front. Mar. Sci. 2019, 6, 664. [Google Scholar] [CrossRef]
- Briseño-Avena, C.; Roberts, P.L.; Franks, P.J.; Jaffe, J.S. ZOOPS-O2: A broadband echosounder with coordinated stereo optical imaging for observing plankton in situ. Methods Oceanogr. 2015, 12, 36–54. [Google Scholar] [CrossRef]
- Cowen, R.K.; Guigand, C.M. In situ ichthyoplankton imaging system (ISIIS): System design and preliminary results. Limnol. Oceanogr. Methods 2008, 6, 126–132. [Google Scholar]
- Olson, R.J.; Sosik, H.M. A submersible imaging-in-flow instrument to analyze nano-and microplankton: Imaging FlowCytobot. Limnol. Oceanogr. Methods 2007, 5, 195–203. [Google Scholar]
- Talapatra, S.; Sullivan, J.; Katz, J.; Twardowski, M.; Czerski, H.; Donaghay, P.; Hong, J.; Rines, J.; McFarland, M.; Nayak, A.R.; et al. Application of in-situ digital holography in the study of particles, organisms and bubbles within their natural environment. Proc. SPIE 2012, 83720, 41–57. [Google Scholar]
- Lertvilai, P. The In situ Plankton Assemblage eXplorer (IPAX): An inexpensive underwater imaging system for zooplankton study. Methods Ecol. Evol. 2020, 11, 1042–1048. [Google Scholar] [CrossRef]
- Picheral, M.; Catalano, C.; Brousseau, D.; Claustre, H.; Coppola, L.; Leymarie, E.; Coindat, J.; Dias, F.; Fevre, S.; Guidi, L. The Underwater Vision Profiler 6: An Imaging Sensor of Particle Size Spectra and Plankton, for Autonomous and Cabled Platforms. Limnol. Oceanogr. Methods 2022, 20, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hastings, M.H.; Green, E.W.; Tauber, E.; Sladek, M.; Webster, S.G.; Kyriacou, C.P.; Wilcockson, D.C. Dissociation of Circadian and Circatidal Timekeeping in the Marine Crustacean Eurydice Pulchra. Curr. Biol. 2013, 23, 1863–1873. [Google Scholar] [CrossRef]
- Ohman, D.M.; Frost, B.W.; Cohen, E.H. Reverse diel vertical migration—An escape from invertebrate predators. Science 1983, 220, 1404–1407. [Google Scholar] [CrossRef]
- Cohen, J.H.; Forward, R.B. Spectral Sensitivity of Vertically Migrating Marine Copepods. Biol. Bull. 2002, 203, 307–314. [Google Scholar] [CrossRef]
- Häfker, N.S.; Meyer, B.; Last, K.S.; Pond, D.W.; Hüppe, L.; Teschke, M. Circadian Clock Involvement in Zooplankton Diel Vertical Migration. Curr. Biol. 2017, 27, 2194–2201.e3. [Google Scholar] [CrossRef] [PubMed]
- Hurley, J.M. Can Road Salt and Other Pollutants Disrupt Our Circadian Rhythms? Available online: http://theconversation.com/can-road-salt-and-other-pollutants-disrupt-our-circadian-rhythms-89211 (accessed on 25 February 2022).
- Dyomin, V.; Davydova, A.; Morgalev, S.; Kirillov, N.; Olshukov, A.; Polovtsev, I.; Davydov, S. Monitoring of plankton spatial and temporal characteristics with the use of a submersible digital holographic camera. Front. Mar. Sci. 2020, 7, 653. [Google Scholar] [CrossRef]
- Sun, H.; Benzie, P.W.; Burns, N.; Hendry, D.C.; Player, M.A.; Watson, J. Underwater digital holography for studies of marine plankton. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2008, 366, 1789–1806. [Google Scholar] [CrossRef]
- Watson, J. Submersible digital holographic cameras and their application to marine science. Opt. Eng. 2011, 50, 091313. [Google Scholar] [CrossRef]
- Bochdansky, A.B.; Jericho, M.H.; Herndl, G.J. Development and deployment of a point-source digital inline holographic microscope for the study of plankton and particles to a depth of 6000 m. Limnol. Oceanogr. Methods 2013, 11, 28–40. [Google Scholar] [CrossRef]
- Rotermund, L.M.; Samson, J.; Kreuzer, J. A Submersible Holographic Microscope for 4-D In-Situ Studies of Micro-Organisms in the Ocean with Intensity and Quantitative Phase Imaging. J. Mar. Sci. Res. Dev. 2015, 06, 1–5. [Google Scholar] [CrossRef]
- Guo, B.; Yu, J.; Yuan, G.; Liu, H.; Xu, W.; Hou, R.; Guo, G. Miniaturized digital inline holographic camera for in-situ plankton detection. Proc. SPIE 2018, 10821, 92–98. [Google Scholar]
- Liu, Z.; Takahashi, T.; Lindsay, D.; Thevar, T.; Sangekar, M.; Watanabe, H.K.; Burns, N.; Watson, J.; Thornton, B. Digital in-line holography for large-volume analysis of vertical motion of microscale marine plankton and other particles. IEEE J. Ocean. Eng. 2021, 46, 1248–1260. [Google Scholar] [CrossRef]
- Dyomin, V.; Gribenyukov, A.; Davydova, A.; Zinoviev, M.; Olshukov, A.; Podzyvalov, S.; Polovtsev, I.; Yudin, N. Holography of particles for diagnostics tasks [Invited]. Appl. Opt. 2019, 58, G300–G310. [Google Scholar] [CrossRef]
- Dyomin, V.; Davydova, A.; Olshukov, A.; Polovtsev, I. Hardware means for monitoring research of plankton in the habitat: Problems, state of the art, and prospects. OCEANS 2019-Marseille 2019, 17, 1–5. [Google Scholar]
- Dyomin, V.; Davydova, A.; Polovtsev, I.; Olshukov, A.; Kirillov, N.; Davydov, S. Underwater Holographic Sensor for Plankton Studies In Situ including Accompanying Measurements. Sensors 2021, 21, 4863. [Google Scholar] [CrossRef] [PubMed]
- Robert, K.O.; Loren, E. Basic techniques. In Applied Time Series Analysis; John Wiley & Sons Inc.: Hoboken, NJ, USA, 1978; Volume 1. [Google Scholar]
- Halberg, F. Temporal coordination of physiologic function. Cold Spring Harb. Symp. Quant. Biol. 1960, 25, 289–310. [Google Scholar] [CrossRef] [PubMed]
- Mirick, D.K.; Davis, S. Melatonin as a biomarker of circadian dysregulation. Cancer Epidemiol. Biomark. Prev. 2008, 17, 3306–3313. [Google Scholar] [CrossRef] [PubMed]
- Njuis, D.; Sulzman, F.; Hastings, J.W. Membrane model for the-circadian clock. Nature 1974, 248, 116–120. [Google Scholar] [CrossRef]
- Sweeney, B.; Borgese, M.B. A circadian rhythm in cell division in a prokaryote the cyanobacterium Synechococcus WH7803. J. Phycol. 1989, 25, 183–186. [Google Scholar] [CrossRef]
- Holzberg, D.; Albrecht, U. The circadian clock: A manager of biochemical processes within the organism. J. Neuroendocrinol. 2003, 15, 339–343. [Google Scholar] [CrossRef]
- Sweeney, B.M. A physiological model for circadian rhythms derived from the Acetabularia rhythm paradoxes. Int. J. Chronobiol. 1974, 2, 25–33. [Google Scholar]
- Weather in the World. Available online: https://rp5.ru/Weather_in_the_world (accessed on 1 June 2022).
- Svodka Izmerenii po Gidropostu pos. Uzur, o. Ol’khon na r. Ozero Baikal Segodnia. Available online: https://allrivers.info/gauge/baykal-uzur/waterlevel (accessed on 1 June 2022). (In Russian).
- Chaffron, S.; Delage, E.; Budinich, M.; Vintache, D.; Henry, N.; Nef, C.; Ardyna, M.; Zayed, A.A.; Junger, P.C.; Galand, P.E.; et al. Environmental vulnerability of the global ocean plankton community interactome. BioRxiv 2020. [Google Scholar] [CrossRef]
- Baikal: Weather Statistics and Wind History. Available online: https://windy.app/forecast2/spot/1998551/Baikal/map (accessed on 1 June 2022).
- Kirillov, N.; Dyomin, V.; Davydova, A.; Polovtsev, I.; Morgalev, S.; Naumova, E.; Olshukov, A. Data of Figure 5 and Figure 7. Figshare Dataset 2022. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value |
|---|---|
| Power characteristics: —Power voltage, V —Power consumption, W | 12 20 |
| Maximum volume studied (working volume) per one exposure, l | 0.5 |
| Provided averaging volume, l | 5 |
| Maximum working volume length, mm | 338.4 |
| Submersion depth, m, not more than | 500 |
| Size of measured particles, mm | 0.1–28 |
| Submersion speed during vertical probing, m/s | 0.1–1.0 |
| Discreteness of counts formed in real time at the submersion speed of 0.3 m/s, m | 6 |
| Ethernet transmission rate, GB/s | 1 |
| Overall dimensions (length × diameter), mm | 320.5 × 142 |
| Weight, kg, not more than | 9 |
| Medium Parameter | Correlation Coefficient for Circadian Rhythm Amplitude with Medium Parameters | Optimal Time Shift of the Signal Reflecting the Change in the Medium Parameter, Day | Cross-Correlation Function Maximum at Optimal Time Shift |
|---|---|---|---|
| Cloudiness | 0.0043 | −11 No signs of causal relationship | −0.383344 |
| Water temperature | −0.62 | 2 | −0.74 |
| Water conductivity | 0.036 | 5 | −0.53 |
| Wind direction | −0.29 | 8 | 0.41 |
| Water level | −0.29 | −5 No signs of causal relationship | −0.42 |
| Date | Diurnal Period of Vertical Migrations, Hour | Frequency of Diurnal Harmonics of Migrations, Hour −1 | Amplitude of Diurnal Harmonics, spp |
|---|---|---|---|
| 4 August 2021 | 8.0 | 0.125 | 1.34 |
| 3.2 | 0.31 | 1.16 | |
| 2.0 | 0.5 | 1.94 | |
| 5 August 2021 | 4.0 | 0.25 | 2.35 |
| 2.67 | 0.375 | 1.79 | |
| 2.0 | 0.5 | 1.56 | |
| 6 August 2021 | 5.3 | 0.188 | 1.69 |
| 3.2 | 0.31 | 2.57 | |
| 2.29 | 0.44 | 1.18 | |
| 7 August 2021 | 4.67 | 0.21 | 2.65 |
| 2.8 | 0.36 | 2.49 | |
| 2.0 | 0.5 | 1.9 | |
| 8 August 2021 | 5.3 | 0.19 | 1.43 |
| 2.67 | 0.38 | 1.58 | |
| 2.0 | 0.5 | 1.2 | |
| 9 August 2021 | 8.0 | 0.125 | 2.92 |
| 4.0 | 0.25 | 3.53 | |
| 2.67 | 0.38 | 1.27 |
| Medium Parameter | Time Shift of the Signal Reflecting the Change in the Medium Parameter, Day | Cross-Correlation Function for Medium Parameter Shift |
|---|---|---|
| Cloudiness | 0 | −0.05 ± 0.2 |
| 1 | −0.30 ± 0.2 | |
| Water temperature | 0 | −0.58 ± 0.2 |
| 2 | −0.74 ± 0.21 | |
| Water conductivity | 0 | −0.02 ± 0.2 |
| 5 | −0.53 ± 0.22 | |
| Wind direction | 0 | −0.34 ± 0.2 |
| 7 | 0.40 ± 0.23 | |
| Water level | 0 | −0.21 ± 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dyomin, V.; Davydova, A.; Kirillov, N.; Morgalev, S.; Naumova, E.; Olshukov, A.; Polovtsev, I. In Situ Measurements of Plankton Biorhythms Using Submersible Holographic Camera. Sensors 2022, 22, 6674. https://doi.org/10.3390/s22176674
Dyomin V, Davydova A, Kirillov N, Morgalev S, Naumova E, Olshukov A, Polovtsev I. In Situ Measurements of Plankton Biorhythms Using Submersible Holographic Camera. Sensors. 2022; 22(17):6674. https://doi.org/10.3390/s22176674
Chicago/Turabian StyleDyomin, Victor, Alexandra Davydova, Nikolay Kirillov, Sergey Morgalev, Elena Naumova, Alexey Olshukov, and Igor Polovtsev. 2022. "In Situ Measurements of Plankton Biorhythms Using Submersible Holographic Camera" Sensors 22, no. 17: 6674. https://doi.org/10.3390/s22176674