
Inertial Measurement of Head Tilt in Rodents: Principles and Applications to Vestibular Research

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Hardware
2.3. Cranial Implants
2.4. Unilateral Vestibular Lesion Model
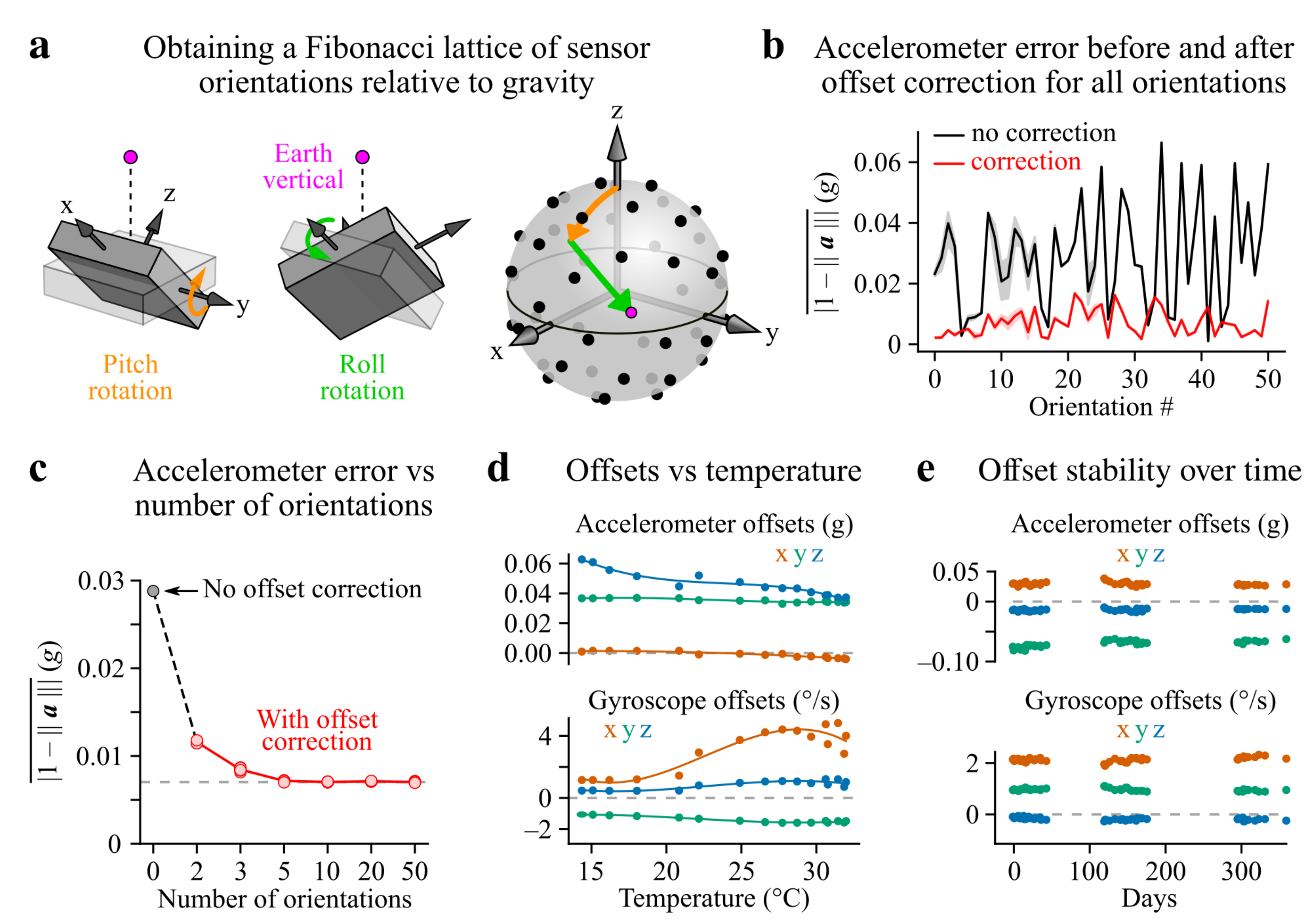
2.5. Sensor Offset Calibration
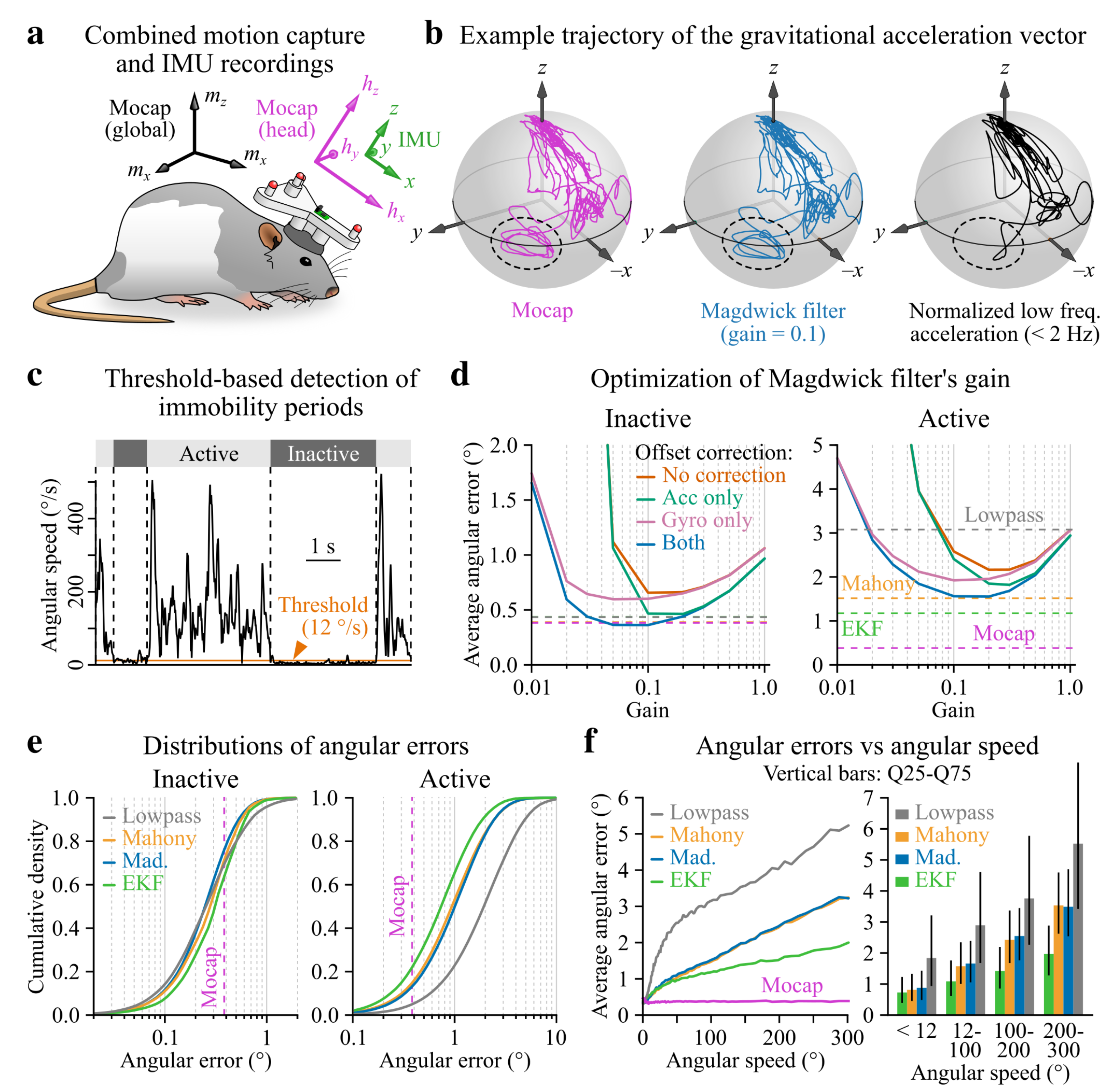
2.6. Benchmarking IMU Filter Algorithms against Optical Motion Capture Data
2.6.1. Experimental Setup
2.6.2. Initial Calibrations
2.6.3. Motion Capture Data Pre-Processing
2.6.4. Calculating Head Tilt Using Motion Capture Data
2.6.5. Calculating Head Tilt Using IMU Data
2.6.6. Computation of IMU-Based Head Tilt Estimation Errors
2.6.7. Computation of Optimal Filter Parameters
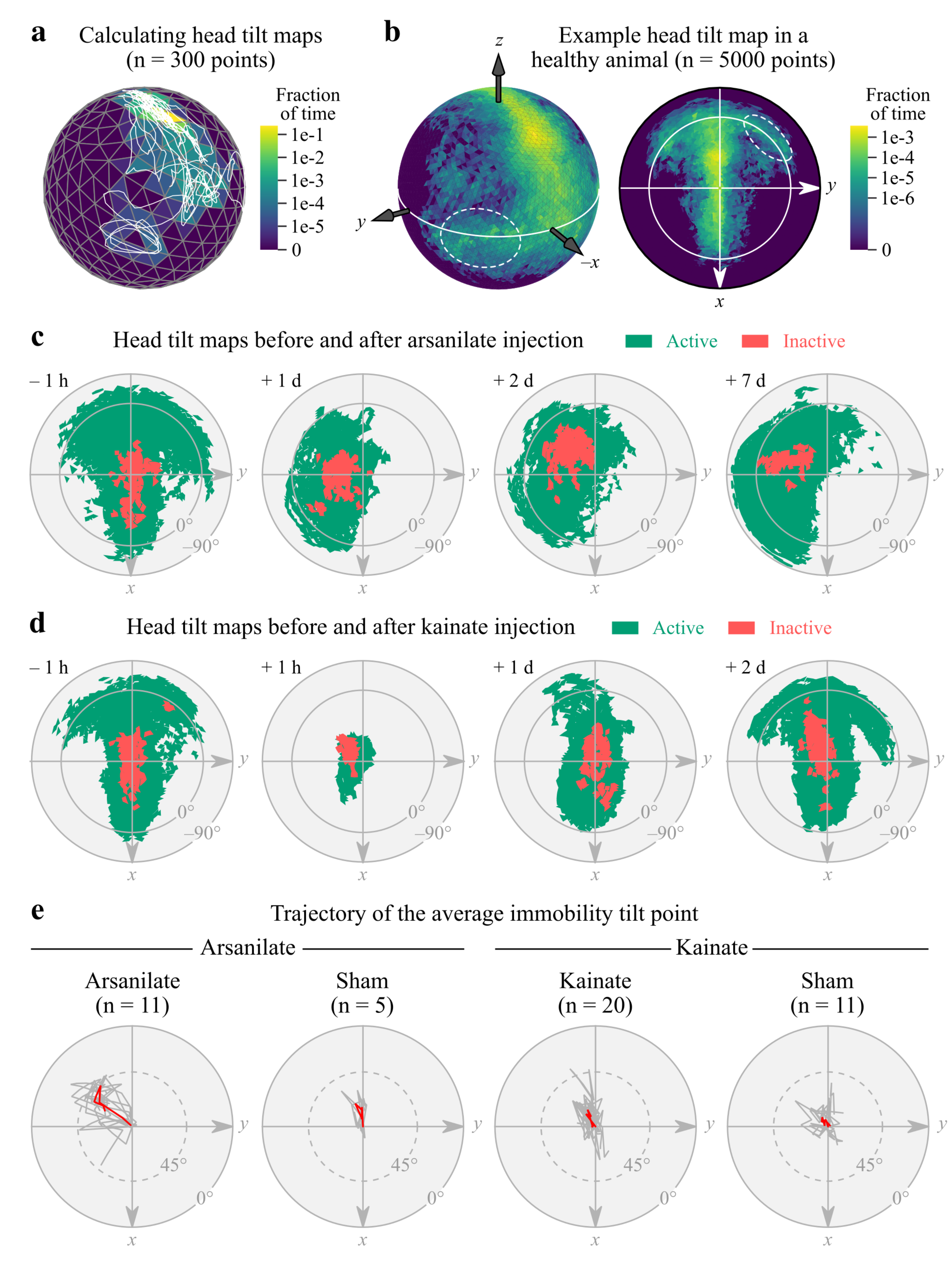
2.7. Calculation of Head Tilt Maps
2.8. Computing the Average Head Tilt Point
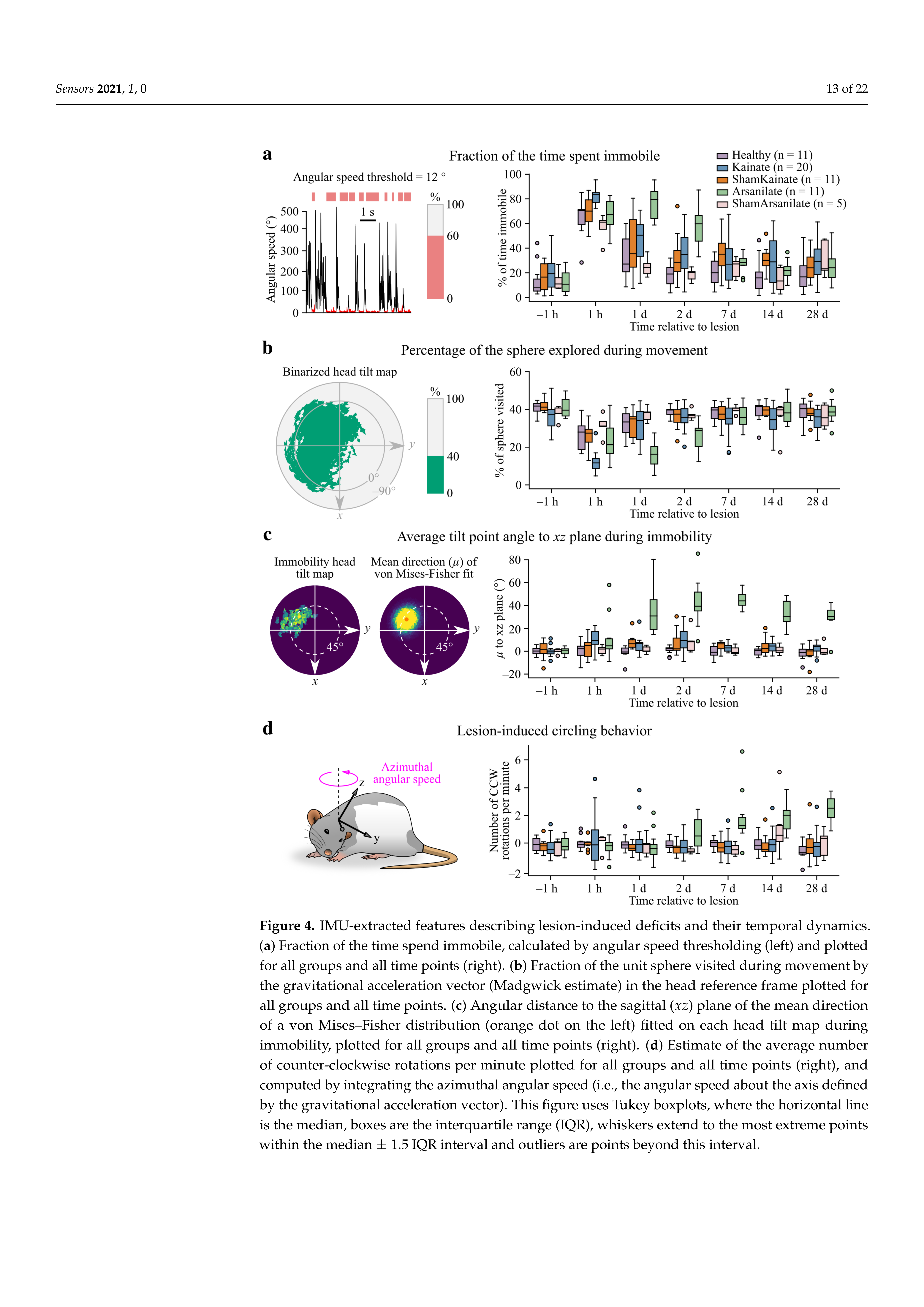
2.9. Detection of Periods of Immobility
2.10. Calculation of a Circling Index
3. Results
3.1. Offset Calibration
3.2. Accuracy of Head Tilt Estimation in Static vs. Dynamic Conditions
3.3. IMU-Based Measurements of Head Tilt in a Rat Model of Unilateral Vestibular Lesion
3.4. Quantitative Assessment of Lesion-Induced Deficits and Their Recovery Using IMU Data
4. Discussion
4.1. Accuracy of IMU-Based Head Tilt Estimation
4.2. Inertial vs. Optical Head Tilt Estimation
4.3. Application of IMUs to Rodent Vestibular Research
4.4. Perspectives: Quantitative Rodent Behavioral Scoring and 3D Orientation Tracking
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Obtaining a Spherical Fibonacci Lattice of Sensor Orientations Relative to Gravity
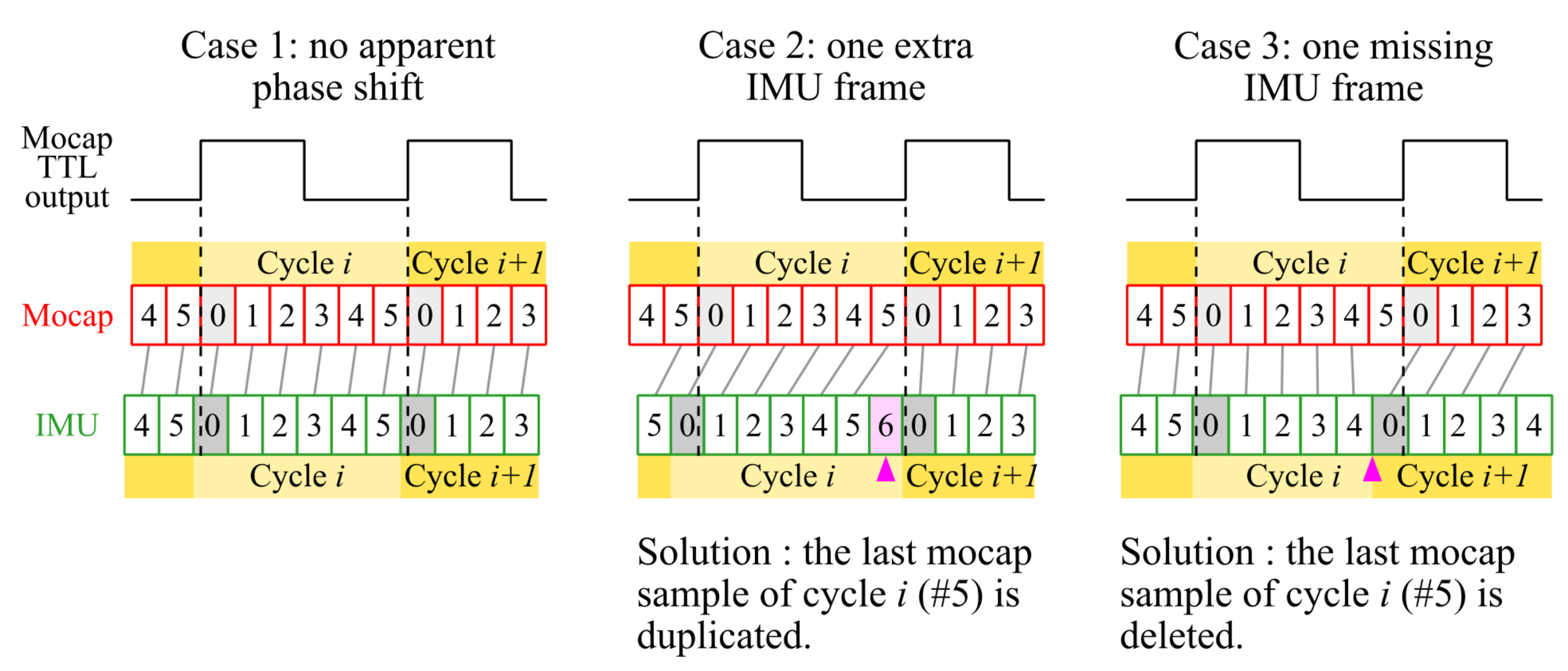
Appendix A.2. Correcting for Minor Phase Shifts between Motion Capture and IMU Acquisition Clocks

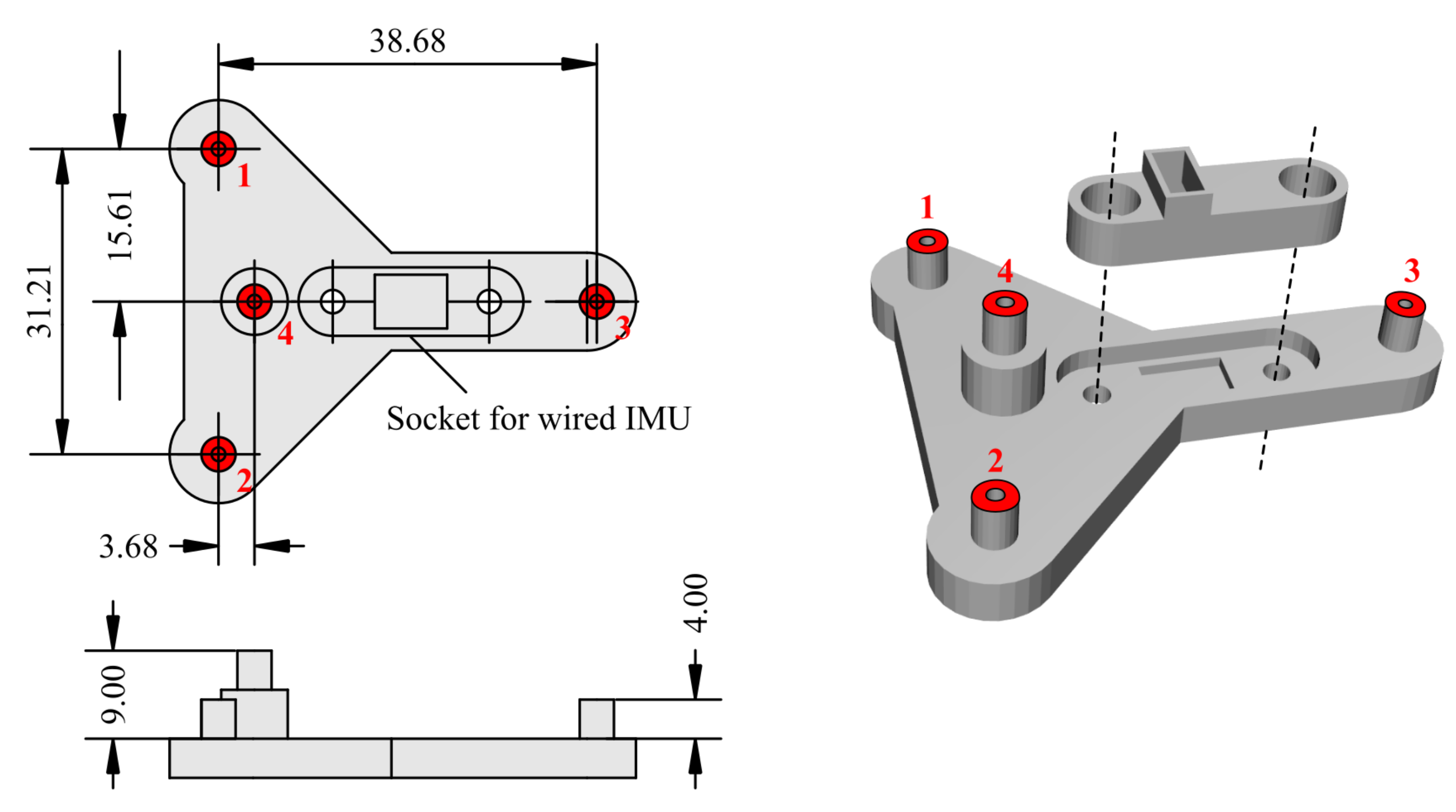
Appendix A.3. Design of the Headborne Support Used for Concurrent Motion Capture and IMU Recordings

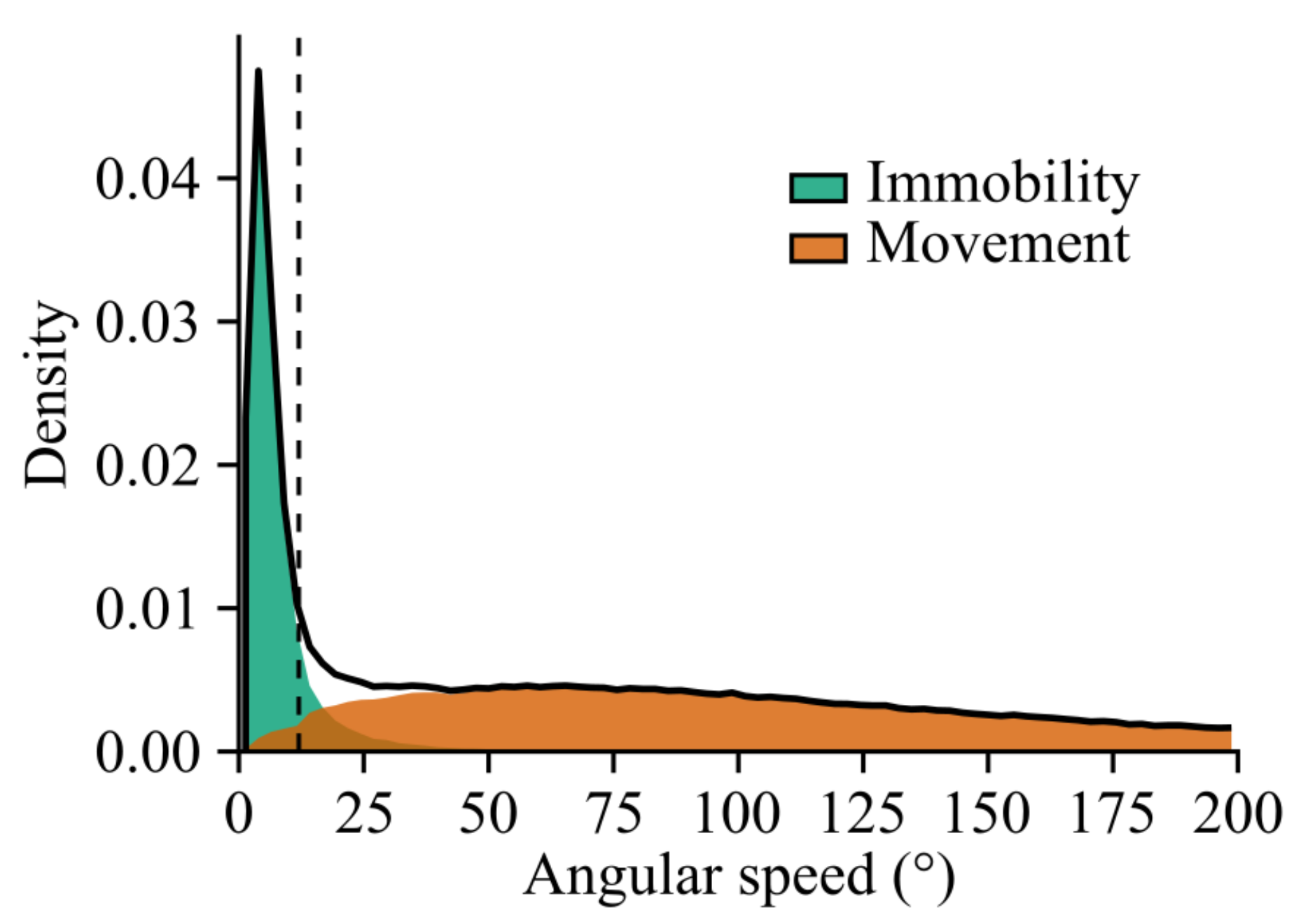
Appendix A.4. Immobility Detection by Angular Speed Thresholding

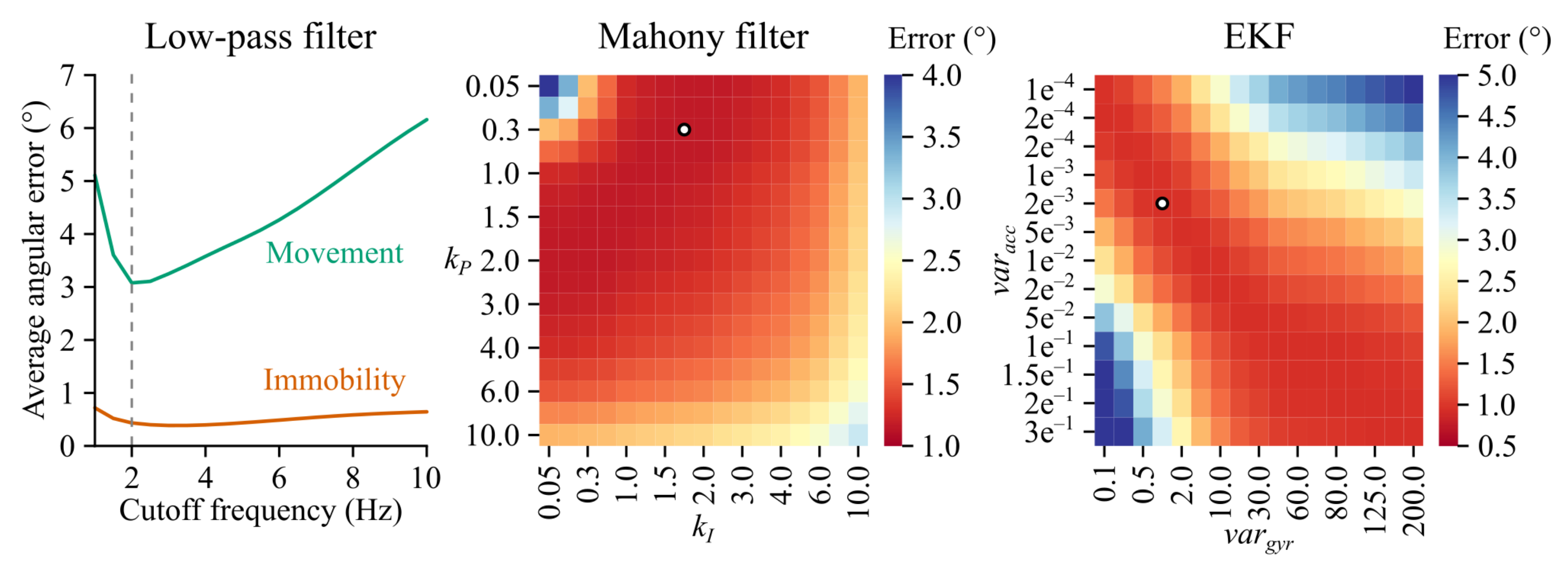
Appendix A.5. Filter Parameter Optimization

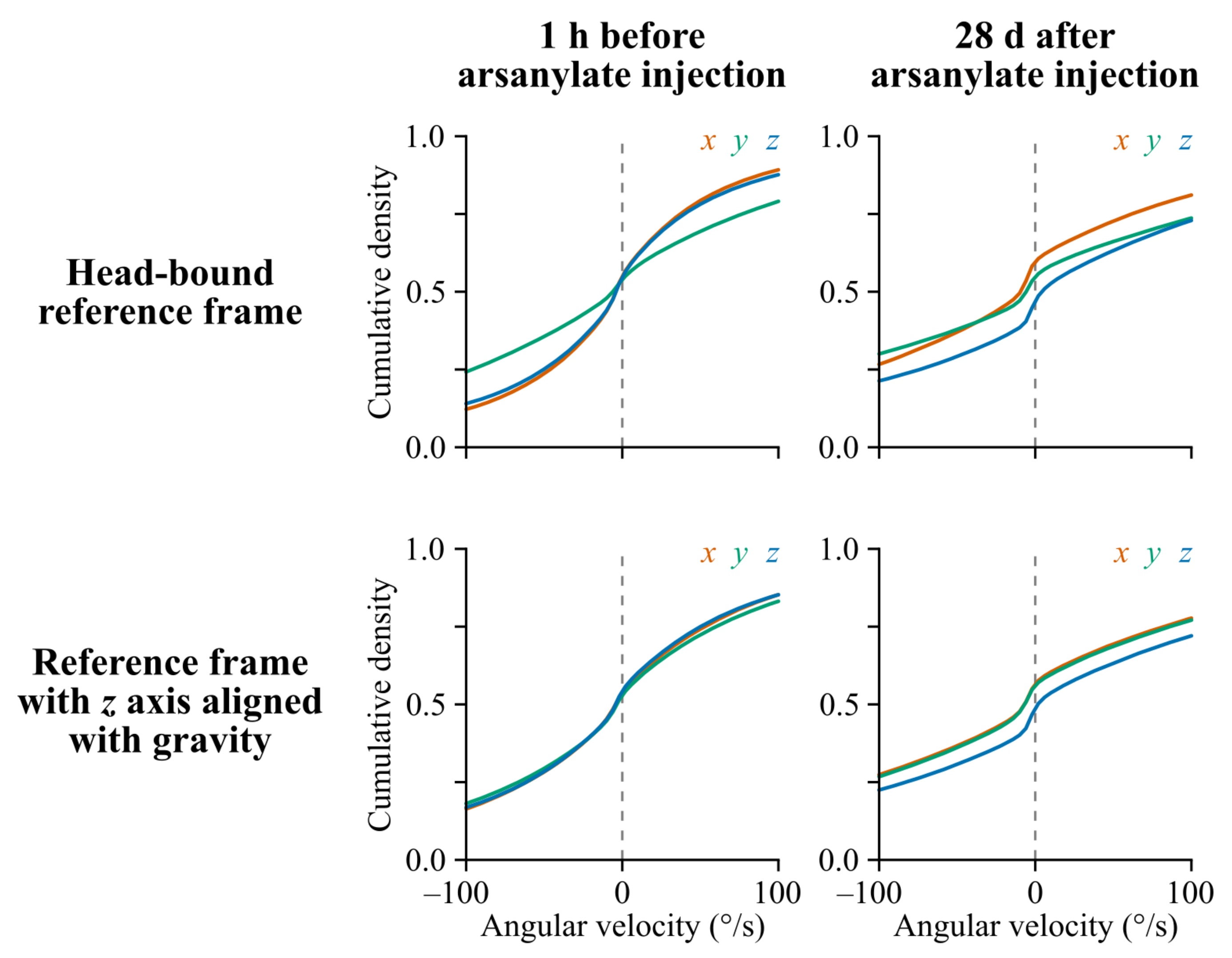
Appendix A.6. Distribution of Head-Centered vs. Gravity-Centered Angular Velocity Values before and after Arsanilate Injection into the Inner Ear

Appendix A.7. Significativity of Inter-Group and Inter-Time Point Differences in the Unilateral Vestibular Lesion Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 h | 1 d | 2 d | 7 d | 14 d | 28 d | |
|---|---|---|---|---|---|---|
| Arsanilate vs. Healthy | 6.202 | <0.001 | <0.001 | 0.790 | 1.702 | 2.379 |
| Arsanilate vs. ShamArsanilate | 1.751 | 0.003 | 0.004 | 5.951 | 1.945 | 13.413 |
| ShamArsanilate vs. Healthy | 16.376 | 14.032 | 9.000 | 5.250 | 9.349 | 1.990 |
| Kainate vs. Healthy | 0.001 | 0.861 | 0.089 | 1.017 | 0.512 | 0.714 |
| Kainate vs. ShamKainate | 0.070 | 4.500 | 4.945 | 14.855 | 9.853 | 4.945 |
| ShamKainate vs. Healthy | 4.991 | 3.876 | 0.686 | 0.321 | 0.136 | 1.511 |
| 1 h | 1 d | 2 d | 7 d | 14 d | 28 d | |
|---|---|---|---|---|---|---|
| Healthy vs. Healthy baseline | 0.003 | 0.009 | 0.961 | 0.126 | 1.239 | 0.618 |
| Arsanilate vs. Arsanilate baseline | 0.001 | 0.001 | 0.003 | 0.006 | 0.041 | 0.029 |
| ShamArsanilate vs. ShamArsanilate baseline | 0.188 | 0.188 | 0.562 | 0.188 | 3.469 | 0.188 |
| Kainate vs. Kainate baseline | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| ShamKainate vs. ShamKainate baseline | 0.003 | 0.009 | 0.202 | 0.073 | 0.009 | 0.249 |
| 1 h | 1 d | 2 d | 7 d | 14 d | 28 d | |
|---|---|---|---|---|---|---|
| Arsanilate vs. Healthy | 3.350 | 0.002 | 0.001 | 4.603 | 5.394 | 6.684 |
| Arsanilate vs. ShamArsanilate | 1.166 | 0.003 | 0.016 | 2.407 | 5.317 | 14.032 |
| ShamArsanilate vs. Healthy | 16.376 | 16.592 | 2.407 | 13.204 | 5.944 | 5.951 |
| Kainate vs. Healthy | <0.001 | 9.000 | 1.617 | 3.512 | 0.426 | 0.449 |
| Kainate vs. ShamKainate | <0.001 | 9.000 | 7.819 | 4.985 | 0.125 | 1.617 |
| ShamKainate vs. Healthy | 7.594 | 8.060 | 2.641 | 6.684 | 8.060 | 6.684 |
| 1 h | 1 d | 2 d | 7 d | 14 d | 28 d | |
|---|---|---|---|---|---|---|
| Healthy vs. Healthy baseline | 0.006 | 0.009 | 0.442 | 0.442 | 0.835 | 1.239 |
| Arsanilate vs. Arsanilate baseline | 0.001 | 0.001 | 0.003 | 0.021 | 0.442 | 0.721 |
| ShamArsanilate vs. ShamArsanilate baseline | 0.188 | 2.438 | 2.438 | 5.438 | 4.688 | 2.438 |
| Kainate vs. Kainate baseline | <0.001 | 0.126 | 3.044 | 2.471 | 0.863 | 1.712 |
| ShamKainate vs. ShamKainate baseline | 0.003 | 0.009 | 0.021 | 0.056 | 0.442 | 0.202 |
| 1 h | 1 d | 2 d | 7 d | 14 d | 28 d | |
|---|---|---|---|---|---|---|
| Arsanilate vs. Healthy | 0.715 | <0.001 | <0.001 | <0.001 | <0.001 | 0.002 |
| Arsanilate vs. ShamArsanilate | 1.440 | 0.003 | 0.029 | 0.004 | 0.001 | 0.016 |
| ShamArsanilate vs. Healthy | 7.442 | 5.250 | 1.990 | 7.442 | 4.724 | 5.250 |
| Kainate vs. Healthy | 0.022 | 0.192 | 0.100 | 0.870 | 0.112 | 0.369 |
| Kainate vs. ShamKainate | 0.220 | 15.443 | 4.005 | 14.296 | 5.207 | 0.596 |
| ShamKainate vs. Healthy | 3.876 | 0.015 | 7.135 | 0.512 | 1.909 | 5.394 |
| 1 h | 1 d | 2 d | 7 d | 14 d | 28 d | |
|---|---|---|---|---|---|---|
| Healthy vs. Healthy baseline | 3.507 | 2.695 | 1.913 | 3.305 | 3.899 | 5.039 |
| Arsanilate vs. Arsanilate baseline | 0.081 | 0.001 | 0.003 | 0.003 | 0.003 | 0.003 |
| ShamArsanilate vs. ShamArsanilate baseline | 1.312 | 1.875 | 0.375 | 1.875 | 0.656 | 2.438 |
| Kainate vs. Kainate baseline | <0.001 | 0.024 | 0.005 | 0.018 | 0.042 | 0.109 |
| ShamKainate vs. ShamKainate baseline | 1.239 | 0.056 | 0.961 | 0.618 | 2.294 | 4.441 |
| 1 h | 1 d | 2 d | 7 d | 14 d | 28 d | |
|---|---|---|---|---|---|---|
| Arsanilate vs. Healthy | 14.650 | 15.706 | 1.511 | 0.012 | 0.003 | <0.001 |
| Arsanilate vs. ShamArsanilate | 17.066 | 12.185 | 0.247 | 0.029 | 1.341 | 0.008 |
| ShamArsanilate vs. Healthy | 3.396 | 13.413 | 17.753 | 17.382 | 1.945 | 4.587 |
| Kainate vs. Healthy | 10.904 | 11.776 | 14.627 | 16.069 | 10.135 | 3.373 |
| Kainate vs. ShamKainate | 8.852 | 3.643 | 5.714 | 8.863 | 10.135 | 11.469 |
| ShamKainate vs. Healthy | 5.394 | 16.298 | 16.663 | 15.865 | 12.188 | 3.221 |
| 1 h | 1 d | 2 d | 7 d | 14 d | 28 d | |
|---|---|---|---|---|---|---|
| Healthy vs. Healthy baseline | 1.731 | 2.101 | 3.305 | 2.493 | 3.507 | 5.903 |
| Arsanilate vs. Arsanilate baseline | 5.096 | 3.627 | 0.161 | 0.015 | 0.006 | 0.003 |
| ShamArsanilate vs. ShamArsanilate baseline | 0.562 | 2.438 | 5.438 | 5.062 | 0.281 | 0.938 |
| Kainate vs. Kainate baseline | 0.990 | 0.537 | 1.425 | 2.324 | 0.961 | 2.782 |
| ShamKainate vs. ShamKainate baseline | 0.961 | 4.441 | 4.087 | 3.305 | 3.507 | 3.899 |
References
- Porciuncula, F.; Roto, A.V.; Kumar, D.; Davis, I.; Roy, S.; Walsh, C.J.; Awad, L.N. Wearable Movement Sensors for Rehabilitation: A Focused Review of Technological and Clinical Advances. Innov. Influ. Phys. Med. Rehabil. 2018, 10, S220–S232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobkin, B.H.; Martinez, C. Wearable Sensors to Monitor, Enable Feedback, and Measure Outcomes of Activity and Practice. Curr. Neurol. Neurosci. Rep. 2018, 18, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasanth, H.; Caban, M.; Keller, U.; Courtine, G.; Ijspeert, A.; Vallery, H.; von Zitzewitz, J. Wearable sensor-based real-time gait detection: A systematic review. Sensors 2021, 21, 2727. [Google Scholar] [CrossRef] [PubMed]
- Taborri, J.; Keogh, J.; Kos, A.; Santuz, A.; Umek, A.; Urbanczyk, C.; van der Kruk, E.; Rossi, S. Sport biomechanics applications using inertial, force, and EMG sensors: A literature overview. Appl. Bionics Biomech. 2020, 2020, 2041549. [Google Scholar] [CrossRef]
- Whitford, M.; Klimley, A.P. An overview of behavioral, physiological, and environmental sensors used in animal biotelemetry and biologging studies. Anim. Biotelem. 2019, 7, 26. [Google Scholar] [CrossRef]
- Leoni, J.; Tanelli, M.; Strada, S.C.; Berger-Wolf, T. Ethogram-based automatic wild animal monitoring through inertial sensors and GPS data. Ecol. Inform. 2020, 59, 101112. [Google Scholar] [CrossRef]
- Venkatraman, S.; Jin, X.; Costa, R.M.; Carmena, J.M. Investigating neural correlates of behavior in freely behaving rodents using inertial sensors. J. Neurophysiol. 2010, 104, 569–575. [Google Scholar] [CrossRef] [Green Version]
- Sauerbrei, B.A.; Lubenov, E.V.; Siapas, A.G. Structured Variability in Purkinje Cell Activity during Locomotion. Neuron 2015, 87, 840–852. [Google Scholar] [CrossRef] [Green Version]
- Pasquet, M.O.; Tihy, M.; Gourgeon, A.; Pompili, M.N.; Godsil, B.P.; Léna, C.; Dugué, G.P. Wireless inertial measurement of head kinematics in freely-moving rats. Sci. Rep. 2016, 6, 35689. [Google Scholar] [CrossRef]
- Wilson, J.J.; Alexandre, N.; Trentin, C.; Tripodi, M. Three-Dimensional Representation of Motor Space in the Mouse Superior Colliculus. Curr. Biol. 2018, 28, 1744–1755.e12. [Google Scholar] [CrossRef] [Green Version]
- Meyer, A.F.; Poort, J.; O’Keefe, J.; Sahani, M.; Linden, J.F. A Head-Mounted Camera System Integrates Detailed Behavioral Monitoring with Multichannel Electrophysiology in Freely Moving Mice. Neuron 2018, 100, 46–60.e7. [Google Scholar] [CrossRef] [Green Version]
- Meyer, A.F.; O’Keefe, J.; Poort, J. Two Distinct Types of Eye-Head Coupling in Freely Moving Mice. Curr. Biol. 2020, 30, 2116–2130.e6. [Google Scholar] [CrossRef]
- Guitchounts, G.; Masís, J.; Wolff, S.B.; Cox, D. Encoding of 3D Head Orienting Movements in the Primary Visual Cortex. Neuron 2020, 108, 512–525.e4. [Google Scholar] [CrossRef]
- Bouvier, G.; Senzai, Y.; Scanziani, M. Head Movements Control the Activity of Primary Visual Cortex in a Luminance-Dependent Manner. Neuron 2020, 108, 500–511.e5. [Google Scholar] [CrossRef]
- Michaiel, A.M.; Abe, E.T.; Niell, C.M. Dynamics of gaze control during prey capture in freely moving mice. eLife 2020, 9, e57458. [Google Scholar] [CrossRef]
- Mallory, C.S.; Hardcastle, K.; Campbell, M.G.; Attinger, A.; Low, I.I.; Raymond, J.L.; Giocomo, L.M. Mouse entorhinal cortex encodes a diverse repertoire of self-motion signals. Nat. Commun. 2021, 12, 671. [Google Scholar] [CrossRef]
- Mathis, A.; Mamidanna, P.; Cury, K.M.; Abe, T.; Murthy, V.N.; Mathis, M.W.; Bethge, M. DeepLabCut: Markerless pose estimation of user-defined body parts with deep learning. Nat. Neurosci. 2018, 21, 1281–1289. [Google Scholar] [CrossRef]
- Karashchuk, P.; Rupp, K.L.; Dickinson, E.S.; Sanders, E.; Azim, E.; Brunton, B.W.; Tuthill, J.C. Anipose: A toolkit for robust markerless 3D poste estimation. bioRxiv 2020. [Google Scholar] [CrossRef]
- Dunn, T.W.; Marshall, J.D.; Severson, K.S.; Aldarondo, D.E.; Hildebrand, D.G.; Chettih, S.N.; Wang, W.L.; Gellis, A.J.; Carlson, D.E.; Aronov, D.; et al. Geometric deep learning enables 3D kinematic profiling across species and environments. Nat. Methods 2021, 18, 564–573. [Google Scholar] [CrossRef]
- Schroeder, J.B.; Ritt, J.T. Selection of head and whisker coordination strategies during goal-oriented active touch. J. Neurophysiol. 2016, 115, 1797–1809. [Google Scholar] [CrossRef] [Green Version]
- Kurnikova, A.; Moore, J.D.; Liao, S.M.; Deschênes, M.; Kleinfeld, D. Coordination of Orofacial Motor Actions into Exploratory Behavior by Rat. Curr. Biol. 2017, 27, 688–696. [Google Scholar] [CrossRef] [Green Version]
- Findley, T.M.; Wyrick, D.G.; Cramer, J.L.; Brown, M.A.; Holcomb, B.; Attey, R.; Yeh, D.; Monasevitch, E.; Nouboussi, N.; Cullen, I.; et al. Sniff-synchronized, gradient-guided olfactory search by freely moving mice. eLife 2021, 10, e58523. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.J.; Greenberg, D.S.; Sawinski, J.; Rulla, S.; Notaro, G.; Kerr, J.N. Rats maintain an overhead binocular field at the expense of constant fusion. Nature 2013, 498, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Klaus, A.; Martins, G.J.; Paixao, V.B.; Zhou, P.; Paninski, L.; Costa, R.M. The Spatiotemporal Organization of the Striatum Encodes Action Space. Neuron 2017, 95, 1171–1180.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullen, K.E.; Sadeghi, S.G.; Beraneck, M.; Minor, L.B. Neural substrates underlying vestibular compensation: Contribution of peripheral versus central processing. J. Vestib. Res. 2009, 19, 171–182. [Google Scholar] [CrossRef]
- Dyhrfjeld-Johnsen, J.; Gaboyard-Niay, S.; Broussy, A.; Saleur, A.; Brugeaud, A.; Chabbert, C. Ondansetron reduces lasting vestibular deficits in a model of severe peripheral excitotoxic injury. J. Vestib. Res. Equilib. Orientat. 2013, 23, 177–186. [Google Scholar] [CrossRef]
- Pan, L.; Qi, R.; Wang, J.; Zhou, W.; Liu, J.; Cai, Y. Evidence for a role of orexin/hypocretin system in vestibular lesion-induced locomotor abnormalities in rats. Front. Neurosci. 2016, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Petremann, M.; Gueguen, C.; Delgado Betancourt, V.; Wersinger, E.; Dyhrfjeld-Johnsen, J. Effect of the novel histamine H4 receptor antagonist SENS-111 on spontaneous nystagmus in a rat model of acute unilateral vestibular loss. Br. J. Pharmacol. 2020, 177, 623–633. [Google Scholar] [CrossRef]
- Gaboyard-Niay, S.; Travo, C.; Saleur, A.; Broussy, A.; Brugeaud, A.; Chabbert, C. Correlation between afferent rearrangements and behavioral deficits after local excitotoxic insult in the mammalian vestibule: A rat model of vertigo symptoms. Dis. Model. Mech. 2016, 9, 1181–1192. [Google Scholar] [CrossRef] [Green Version]
- Cassel, R.; Bordiga, P.; Carcaud, J.; Simon, F.; Beraneck, M.; Le Gall, A.; Benoit, A.; Bouet, V.; Philoxene, B.; Besnard, S.; et al. Morphological and functional correlates of vestibular synaptic deafferentation and repair in a mouse model of acute-onset vertigo. Dis. Model. Mech. 2019, 12, 39115. [Google Scholar] [CrossRef] [Green Version]
- Vignaux, G.; Chabbert, C.; Gaboyard-Niay, S.; Travo, C.; Machado, M.L.; Denise, P.; Comoz, F.; Hitier, M.; Landemore, G.; Philoxène, B.; et al. Evaluation of the chemical model of vestibular lesions induced by arsanilate in rats. Toxicol. Appl. Pharmacol. 2012, 258, 61–71. [Google Scholar] [CrossRef]
- Montardy, Q.; Wei, M.; Liu, X.; Yi, T.; Zhou, Z.; Lai, J.; Zhao, B.; Besnard, S.; Tighilet, B.; Chabbert, C.; et al. Selective optogenetic stimulation of glutamatergic, but not GABAergic, vestibular nuclei neurons induces immediate and reversible postural imbalance in mice. Prog. Neurobiol. 2021, 204, 102085. [Google Scholar] [CrossRef]
- Tighilet, B.; Péricat, D.; Frelat, A.; Cazals, Y.; Rastoldo, G.; Boyer, F.; Dumas, O.; Chabbert, C. Adjustment of the dynamic weight distribution as a sensitive parameter for diagnosis of postural alteration in a rodent model of vestibular deficit. PLoS ONE 2017, 12, e0187472. [Google Scholar] [CrossRef] [Green Version]
- Rastoldo, G.; Marouane, E.; El Mahmoudi, N.; Péricat, D.; Bourdet, A.; Timon-David, E.; Dumas, O.; Chabbert, C.; Tighilet, B. Quantitative Evaluation of a New Posturo-Locomotor Phenotype in a Rodent Model of Acute Unilateral Vestibulopathy. Front. Neurol. 2020, 11, 505. [Google Scholar] [CrossRef]
- Marouane, E.; Rastoldo, G.; El Mahmoudi, N.; Péricat, D.; Chabbert, C.; Artzner, V.; Tighilet, B. Identification of New Biomarkers of Posturo-Locomotor Instability in a Rodent Model of Vestibular Pathology. Front. Neurol. 2020, 11, 470. [Google Scholar] [CrossRef]
- Houpt, T.A.; Cassell, J.; Carella, L.; Neth, B.; Smith, J.C. Head tilt in rats during exposure to a high magnetic field. Physiol. Behav. 2012, 105, 388–393. [Google Scholar] [CrossRef] [Green Version]
- Inayat, S.; Singh, S.; Ghasroddashti, A.; Qandeel; Egodage, P.; Whishaw, I.Q.; Mohajerani, M.H. A matlab-based toolbox for characterizing behavior of rodents engaged in string-pulling. eLife 2020, 9, e54540. [Google Scholar] [CrossRef]
- Ebbesen, C.; Froemke, R. Automatic tracking of mouse social posture dynamics by 3D videography, deep learning and GPU-accelerated robust optimization. bioRxiv 2020. [Google Scholar] [CrossRef]
- Marshall, J.D.; Aldarondo, D.E.; Dunn, T.W.; Wang, W.L.; Berman, G.J.; Ölveczky, B.P. Continuous Whole-Body 3D Kinematic Recordings across the Rodent Behavioral Repertoire. Neuron 2021, 109, 420–437.e8. [Google Scholar] [CrossRef]
- Dugué, G.P.; Tihy, M.; Gourévitch, B.; Léna, C. Cerebellar re-encoding of self-generated head movements. eLife 2017, 6, e26179. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Hartsock, J.J.; Dai, C.; Salt, A.N. Permeation enhancers for intratympanically-applied drugs studied using fluorescent dexamethasone as a marker. Otol. Neurotol. 2018, 39, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0: Fundamental algorithms for scientific computing in Python. Nat. Methods 2020, 17, 261–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahony, R.; Hamel, T.; Pflimlin, J.M. Nonlinear complementary filters on the special orthogonal group. IEEE Trans. Autom. Control. 2008, 53, 1203–1218. [Google Scholar] [CrossRef] [Green Version]
- Madgwick, S.O.H.; Harrison, A.J.L.; Vaidyanathan, R. Estimation of IMU and MARG orientation using a gradient descent algorithm. In Proceedings of the 2011 IEEE International Conference on Rehabilitation Robotics, Zurich, Switzerland, 29 June–1 July 2011. [Google Scholar] [CrossRef]
- Lam, S.K.; Pitrou, A.; Seibert, S. Numba: A LLVM-based Python JIT compiler. In Proceedings of the Second Workshop on the LLVM Compiler Infrastructure in HPC (LLVM ’15), Austin, TX, USA, 15 November 2015. [Google Scholar] [CrossRef]
- Kabsch, W. A solution for the best rotation to relate two sets of vectors. Acta Crystallogr. 1976, A32, 922–923. [Google Scholar] [CrossRef]
- Hillen, T.; Painter, K.J.; Swan, A.C.; Murtha, A.D. Moments of von Mises and Fisher distributions and applications. Math. Biosci. Eng. 2017, 14, 673–694. [Google Scholar] [CrossRef] [Green Version]
- Hunt, M.A.; Miller, S.W.; Nielson, H.C.; Horn, K.M. Intratympanic Injection of Sodium Arsanilate (Atoxyl) Solution Results in Postural Changes Consistent With Changes Described for Labyrinthectomized Rats. Behav. Neurosci. 1987, 101, 427–428. [Google Scholar] [CrossRef]
- Soler-Martín, C.; Dí-Padrisa, N.; Boadas-Vaello, P.; Llorens, J. Behavioral disturbances and hair cell loss in the inner ear following nitrile exposure in mice, guinea pigs, and frogs. Toxicol. Sci. 2007, 96, 123–132. [Google Scholar] [CrossRef]
- Ossenkopp, K.P.; Prkacin, A.; Hargreaves, E.L. Sodium arsanilate-induced vestibular dysfunction in rats: Effects on open-field behavior and spontaneous activity in the automated digiscan monitoring system. Pharmacol. Biochem. Behav. 1990, 36, 875–881. [Google Scholar] [CrossRef]
- Bauer, P.; Lienhart, W.; Jost, S. Accuracy investigation of the pose determination of a vr system. Sensors 2021, 21, 1622. [Google Scholar] [CrossRef]
- Goldberg, J.M.; Cullen, K.E. Vestibular control of the head: Possible functions of the vestibulocollic reflex. Exp. Brain Res. 2011, 210, 331–345. [Google Scholar] [CrossRef] [Green Version]




| Angular Error during Immobility () | Angular Error during Movement () | |||||||
|---|---|---|---|---|---|---|---|---|
| Lowpass | Madgwick | Mahony | EKF | Lowpass | Madgwick | Mahony | EKF | |
| Mean | 0.43 | 0.36 | 0.39 | 0.44 | 3.08 | 1.56 | 1.52 | 1.17 |
| Std | 0.41 | 0.25 | 0.25 | 0.27 | 2.58 | 1.23 | 1.26 | 0.99 |
| Median | 0.32 | 0.31 | 0.35 | 0.39 | 2.44 | 1.27 | 1.19 | 0.93 |
| Q25 | 0.18 | 0.19 | 0.20 | 0.24 | 1.35 | 0.73 | 0.67 | 0.53 |
| Q75 | 0.55 | 0.47 | 0.52 | 0.58 | 4.12 | 2.08 | 2.00 | 1.55 |
| Q95 | 1.16 | 0.81 | 0.83 | 0.91 | 7.67 | 3.83 | 3.87 | 2.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fayat, R.; Delgado Betancourt, V.; Goyallon, T.; Petremann, M.; Liaudet, P.; Descossy, V.; Reveret, L.; Dugué, G.P. Inertial Measurement of Head Tilt in Rodents: Principles and Applications to Vestibular Research. Sensors 2021, 21, 6318. https://doi.org/10.3390/s21186318
Fayat R, Delgado Betancourt V, Goyallon T, Petremann M, Liaudet P, Descossy V, Reveret L, Dugué GP. Inertial Measurement of Head Tilt in Rodents: Principles and Applications to Vestibular Research. Sensors. 2021; 21(18):6318. https://doi.org/10.3390/s21186318
Chicago/Turabian StyleFayat, Romain, Viviana Delgado Betancourt, Thibault Goyallon, Mathieu Petremann, Pauline Liaudet, Vincent Descossy, Lionel Reveret, and Guillaume P. Dugué. 2021. "Inertial Measurement of Head Tilt in Rodents: Principles and Applications to Vestibular Research" Sensors 21, no. 18: 6318. https://doi.org/10.3390/s21186318