
Electroencephalogram-Electromyogram Functional Coupling and Delay Time Change Based on Motor Task Performance



Abstract
:1. Introduction
2. Materials
2.1. Participants
2.2. Experimental Setting
2.3. Experimental Design and Procedures
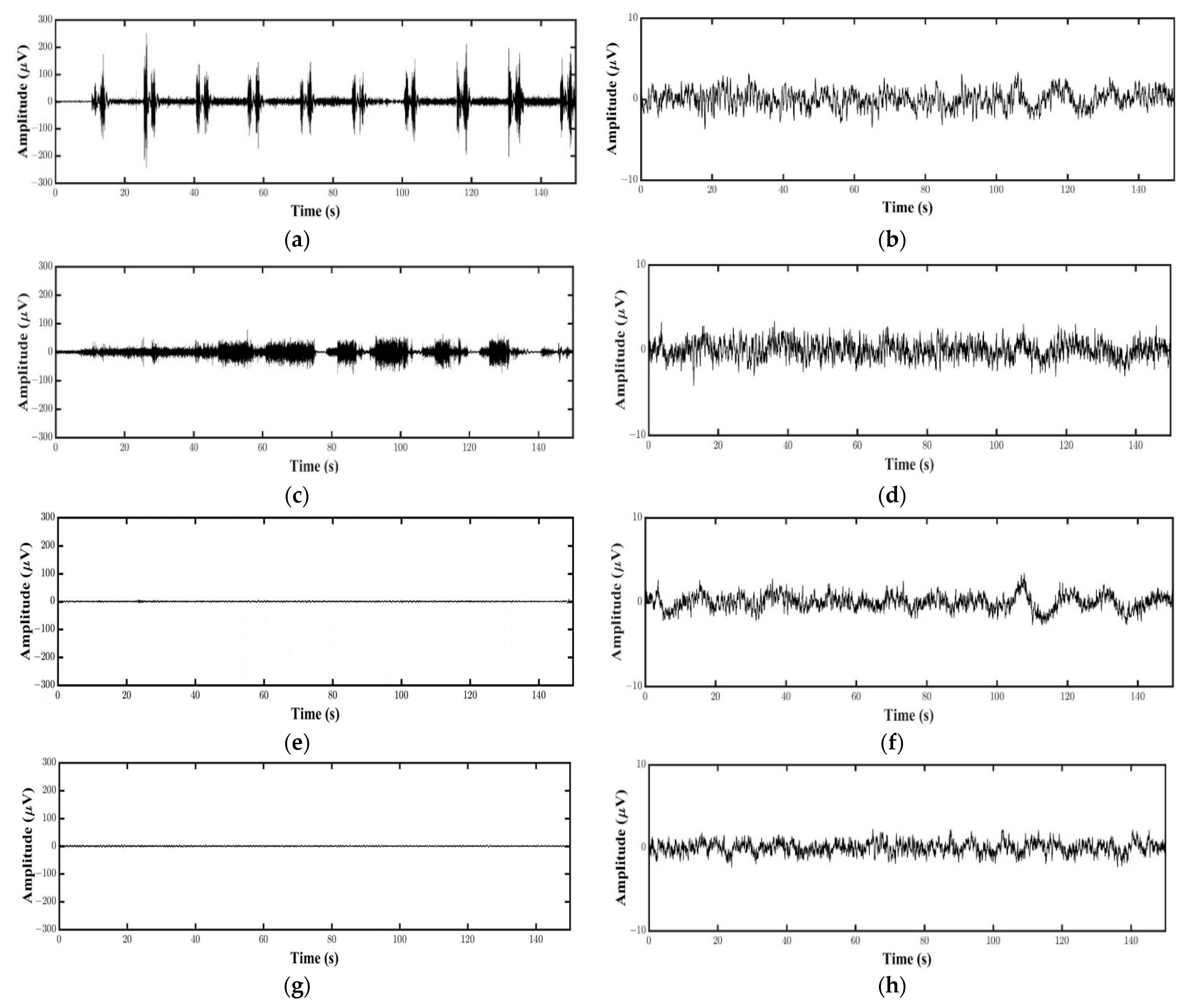
2.4. Data Analysis
3. Methods
3.1. Functional Coupling of Brain and Muscle Signals with Linear Correlation Analysis
3.2. Functional Coupling of Brain and Muscle Signals with Nonlinear Correlation Analysis
3.3. Delay Time Investigation with Nonlinear Mutual Information Flow
4. Results
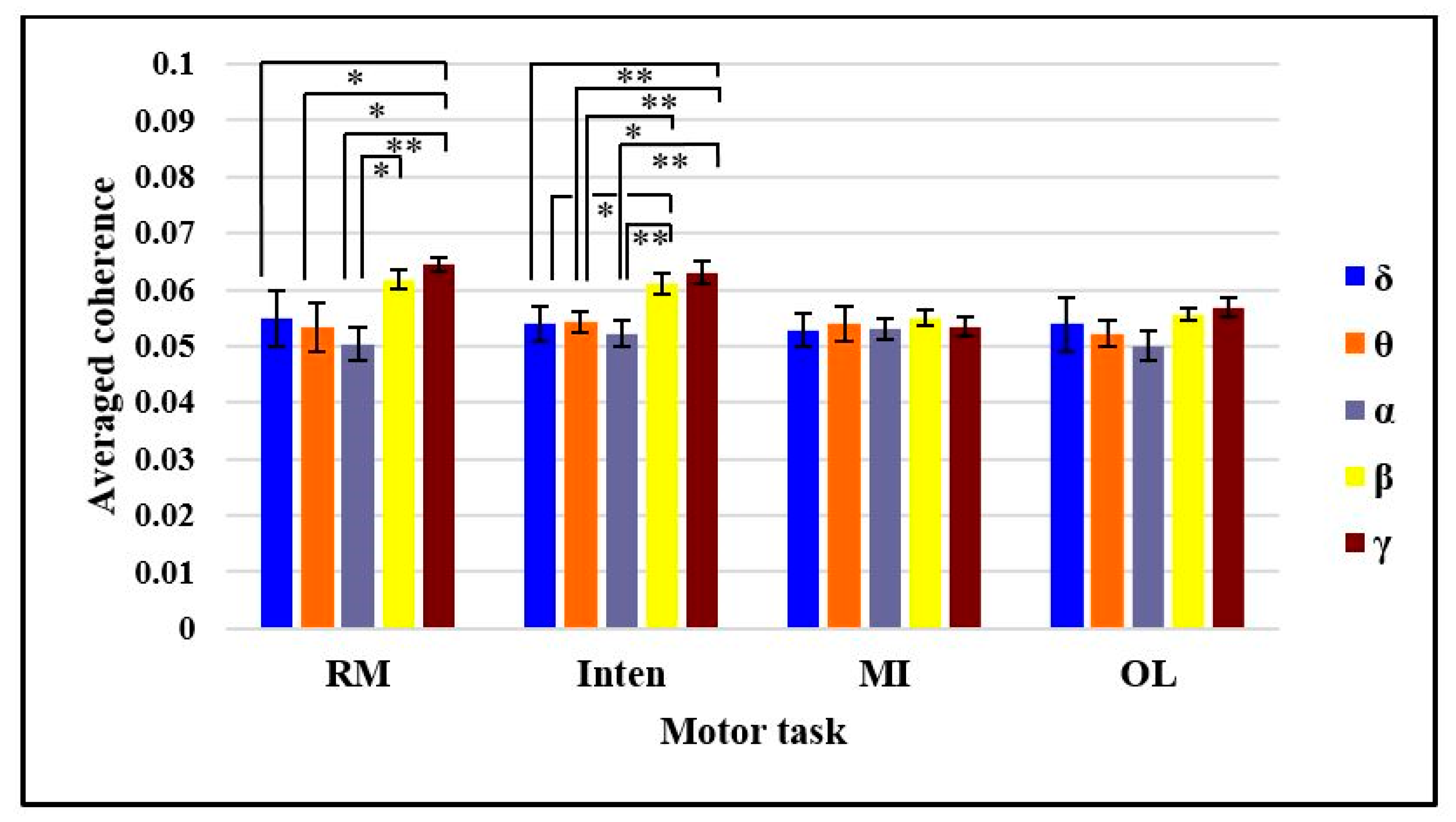
4.1. Comparison of EEG-EMG Coherence in Each Motor Task Based on Bands
4.2. Comparison of EEG-EMG Coherence in Beta Band and Gamma Bands Based on Motor Tasks
4.3. Comparison of EEG-EMG Mutual Information Based on Motor Tasks
4.4. Comparison EEG-EMG Averaged Mutual Information Across All Subjects
4.5. Calculation Delay Time between Brain Motor Cortex and Peripheral Muscle
4.6. Statistical Analysis of Delay Time across All Subjects
5. Discussion
5.1. EEG-EMG Functional Coupling Analysis Using Linear Coherence
5.2. EEG-EMG Functional Coupling Analysis Using Nonlinear Mutual Information
5.3. Functional Coupling Delay Time Change Based on Motor Tasks
5.4. Real-World Applications, Limitations and Future Works of Study
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ref. | Authors | Investigated Area | Method | Strengths and Weakness |
|---|---|---|---|---|
| [3] | Yasunari, H., et al., 2010 | EEG-EMG coherence during Isometric contraction and its imagery. | Power spectrum. EEG—Cz, FCz, C1, C2, CPz. Rectified EMG—right TA muscle. n = 13 | Coherence occurred in motor imagery conditions. Uses linear correlation analysis. Remains controversial issues of EMG during motor imagery. |
| [9] | James, M., et al., 2000 | Task-dependent modulation in coherence between motor cortex and hand muscles. | Amplitude and phase correlation method. MEG—over the left sensorimotor cortex. EMG—1DI, AbPB, FDS, EDC. n = 13 | Tests task-dependent modulation of coherence. Coherence was much lower level during isometric grip of the fixed levers compared to grasp under complaint conditions. Remains to investigate with different motor tasks. |
| [10] | Shinji, O., et al., 2000 | ECoG-EMG coherence during isometric contraction in hand muscle. | Auto spectra and frequency domain analysis method. Rectified EMG. ECoG—mesial and lateral surfaces of frontoparietal cortices. EMG—ECR muscle. n = 8 (patients with epilepsy) | Coherence occurred only in the 15 ± 3 Hz beta bands. Time lags were calculated with a cross-correlogram method. Time lags range from 10 ms to 22 ms. Lack of directionality inference and nonlinear correlation. Remains to find out the coherence in other bands. |
| [22] | Wolfgang, O., et al., 2006 | Gamma range Cortico-muscular coherence during dynamic performance in visuo motor tasks. | Cortico motor spectral power method. EEG—52 electrodes. Rectified EMG—flexor digitorum superficialis muscle. n = 8 | Beta band coherences occur during static force. Gamma band coherence occurs during dynamic force. Uses only linear correlation method. No include delay time estimation. Remains task-dependent CMC investigation. |
| [34] | Seung-Hyun, P., et al., 2010 | Linear and nonlinear information flow with time delay mutual information. | Used surrogate data sets and experimental data sets. Investigated CM interaction during a right wrist extension tasks. EEG—29 electrodes. EMG—Extensor digitorum communis. n = 7 | Well-distinguished linear and nonlinear information flow. Requires relatively long stationary time series data for the analysis. Needs to improve directionality inferences with stationarity. |
| [41] | Andreas, W., et al., 2012 | Time delay mutual information of the phase as a measure of functional connectivity. | Phase lag index and weighted phase lag index methods. Making numerical implementation. Synthetic data sets by a mutual amplitude coupled network of Rossler oscillatior. | Limitations and assumptions existed as synthetic data sets were applied. De-correlation step does not respect a background synchronicity. Uses a data-driven approach. |
References
- Valery, L.F.; Bo, N.; George, A.M. Global burden of stroke. Circ. Res. 2017, 120, 439–448. [Google Scholar] [CrossRef]
- Richard, K.; Johanna, S.; Susanne, V.; Sabine, L.; Micheal, S.; Sickert, A.; Lamprecht, J.; Perdikis, S.; Corbet, T.; Millán, J.D.R.; et al. Cortico-muscular coherence is reduced actuely post-stroke and increases bilaterally during motor recovery: A pilot study. J. Front. Neurol. 2019, 10, 1–11. [Google Scholar]
- Yasunari, H.; Junichi, U.; Akio, K.; Meigen, L.; Yutaka, T. Correlation between EEG-EMG coherence during isometric contraction and its imaginary execution. Acta Neurol. Exp. 2010, 70, 76–85. [Google Scholar]
- Yuhang, X.; Verity, M.M.; Zoran, C.; Kerry, R.M. Corticomuscular coherence with time lag with application to delay estimation. IEEE Trans. Biomed. Eng. 2016, 64, 588–600. [Google Scholar] [CrossRef] [Green Version]
- Conway, B.A.; Halliday, D.M.; Farmer, S.F.; Shahani, U.; Mass, P.; Weir, A.I.; Rosenberg, J.R. Synchronization between motor cortex and spinal motoneuronal pool during the performance of a maintained motor task in man. J. Physiol. 1995, 489, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, J.R.; Amjad, A.M.; Breeze, P.; Brillinger, D.R.; Halliday, D.M. The Fourier approach to the identification of functional coupling between neuronal spike trains. Prog. Biophys. Mol. Biol. 1989, 53, 1–31. [Google Scholar] [CrossRef]
- Tatsuya, M.; Keiichiro, T.; Benjamin, K.; Mark, H. Coherence between cortical and muscular activities after subcortical stroke. Ameri. Heart Asso. 2001, 32, 2597–2601. [Google Scholar] [CrossRef]
- Yang, Z.; Yu, P.; Guanghua, X.; Long, L.; Jue, W. Using corticomuscular coherence to reflect function recovery of paretic upper limb after stroke: A case study. Front. Neurol. 2018, 8, 728. [Google Scholar] [CrossRef]
- James, M.K.; Stuart, N.B.; Stephan, S.; Riitta, H.; Roger, N.L. Human cortical muscle coherence is directly related to specific motor parameters. J. Neurosci. 2000, 20, 8838–8845. [Google Scholar] [CrossRef]
- Shinji, O.; Takashi, N.; Akio, I.; Takeharu, K.; Riki, M.; Waro, T.; Nobuo, H.; Koichi, B.; Tadahiro, M.; Stephan, S.; et al. Electrocorticogram-Electromyogram coherence during isometric contraction of hand muscle in human. Clin. Neurophysiol. 2000, 111, 2014–2024. [Google Scholar]
- Carlowitz-Ghori, K.V.; Bayraktaroglu, Z.; Waterstraat, G.; Curio, G.; Nikulin, V.V. Voluntary control of corticomuscular coherence through neurofeedback: A proof of principle study in health subjects. Neuroscience 2015, 290, 243–254. [Google Scholar] [CrossRef]
- Anirban, C.; Haider, R.; Yogesh, K.M.; Ashish, D.; Girijesh, P. An EEG-EEM correlation based brain computer interface for hand orthosis supported neuro-rehabilitation. J. Neurosci. 2019, 312, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Xugang, X.; Cunbin, M.; Changmin, Y.; Seyed, M.M.; Xian, H.; Yun, B.Z.; Zhizeng, L. Enhanced EEG-EMG coherence analysis based on hand movements. Biomed. Sig. Pro. Cont. 2020, 56, 1–10. [Google Scholar]
- Xugang, X.; Ziyang, S.; Xian, H.; Changmin, Y.; Yun-Bo, Z.; Seyed, M.M.; Zhizeng, L.; Zhong, L. Construction and analysis of cortical-muscular functional network based on EEG-EMG coherence using wavelet coherence. Neurocomputing 2021, 438, 248–258. [Google Scholar]
- Joseph, T.G.; Daniel, P.F. Beta-and gamma-range human lower limb corticomuscular coherence. Front. Hum. Neurosci. 2012, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Pascal, G.; Peter, B. Corticomuscular and intermuscular frequency analysis. Physiological principles and applications in disorders of the motor system. Suppl. Clin. Neurophysiol. 2004, 57, 570–576. [Google Scholar]
- Fabien, D.M.; Marieke, L.; Sylvain, C.; David, A. Effect of training status on beta-range corticomuscular coherence in agonist vs. antagonist muscles during isometric knee contractions. Exp. Brain Res. 2017, 235, 3023–3031. [Google Scholar] [CrossRef]
- Saeid, M.; Michael, B.; Tjeerd, W.B. The reorganization of corticomuscular coherence during a transition between sensorimotor states. NeuroImage 2014, 100, 692–702. [Google Scholar] [CrossRef]
- Hellwig, B.; Häussler, S.; Lauk, M.; Guschlbauer, B.; Köster, B.; KristevaFeige, R.; Timmer, J.; Lucking, C.H. Tremor-correlated cortical activity detected by electroencephalography. Clin. Neurophy. 2000, 111, 806–809. [Google Scholar] [CrossRef]
- Lars, T.; Joachim, G.; Martin, D.; Jens, V.; Hans, J.F.; Alfons, S. The cerebral oscillatory network of parkinsonian resting tremor. Brain 2003, 126, 199–212. [Google Scholar] [CrossRef]
- Jan-Mathiji, S.; Robert, O.; Pascal, F. Neuronal coherence as a mechanism of effective corticospinal interaction. Science 2005, 308, 111–113. [Google Scholar] [CrossRef] [Green Version]
- Wolfgang, O.; Luis, P.; Marie-Claude, H.R.; Rumyana, K. Gamma-range corticomuscular coherence during dynamic force output. NeuroImage 2007, 34, 1191–1198. [Google Scholar] [CrossRef]
- Tatsuya, M.; Jurg, S.; Andrew, E.S.; Christian, G.; Mark, H. Electroencephalographic measurement of motor cortex control of muscle activity in humans. Clin. Neurophysio. 2000, 111, 326–337. [Google Scholar]
- Rumyana, K.F.; Christoph, F.; Jens, T.; Carl, H.L. Effects of attention and precision of exerted force on beta range EEG-EMG synchronization during a maintained motor contraction task. Clin. Neurophy. 2002, 119, 124–131. [Google Scholar]
- Tianchen, Z.; Anshuang, F.; Rui, X.; Shuang, Q.; Hongzhi, Q.; Peng, Z.; Lixin, Z.; Baikun, W.; Dong, M. Cortico-muscular coherence analysis under voluntary, simulated, and imagery neuromuscular activities. Bio. Tech. 2013, 58. [Google Scholar] [CrossRef]
- Rui, X.; Feng, H.; Shuang, Q.; Chen, L.; Shuang, L.; Xin, Z.; Hongzhi, Q.; Lixin, Z.; Baikun, W.; Dong, M. Corticomuscular coherence based on wavelet transform during imagination, execution and stimulation tasks. J. Med. Imag. Health Inform. 2015, 5, 335–341. [Google Scholar] [CrossRef]
- Byeongnam, K.; Laehyun, K.; Yun-Hee, K.; Sun, K.Y. Cross-association analysis of EEG and EMG signals according to movement intention state. Sci. Dire. 2017, 44, 1–9. [Google Scholar] [CrossRef]
- Verity, M.M.; Zoran, C.; Kerry, R.M. Rectification of the EMG is an unnecessary and inappropriate step in the calculation of cortico-muscular coherence. J. Neurosci. Meth. 2012, 205, 190–201. [Google Scholar] [CrossRef]
- Liu, J.; Sheng, Y.; Liu, H. Corticomuscular coherence and its applications: A review. Front. Neurol. 2019, 13, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Myers, L.J.; Lowery, M.; O’Malley, M.; Vaughan, C.L.; Heneghan, C.; St Clair Gibson, A.; Harley, Y.X.R.; Sreenivasan, R. Rectification and non-linear preprocessing of EMG signals for cortico-muscular analysis. J. Neurol. 2003, 124, 157–165. [Google Scholar]
- Jaeseung, J.; John, C.G.; Bradley, S.P. Mutual information analysis of the EEG in patients with Alzheimer’s disease. Clin. Neurophysiol. 2001, 112, 827–835. [Google Scholar] [CrossRef]
- Thomas, M.C.; Joy, A.T. Elements of Information Theory, Solutions to Problems, 2nd ed.; John and Wiley and Sons: New York, NY, USA, 1991; pp. 1–397. [Google Scholar]
- Sun, H.N.; Seung-Hyun, J.; Soo, Y.K.; Byung-Joo, H. EEG in schizophrenic patients: Mutual information analysis. Clini. Neurophy. 2002, 113, 1954–1960. [Google Scholar]
- Seung-Hyun, J.; Peter, L.; Mark, H. Linear and nonlinear information flow based on time-delayed mutual information method and its application to corticomuscular interaction. Clini. Neurophy. 2010, 121, 392–401. [Google Scholar] [CrossRef] [Green Version]
- Stuart, N.B. Oscillatory interactions between sensorimotor cortex and the periphery. Sci. Dire. 2007, 17, 649–655. [Google Scholar]
- Mike, X.C. Chapter 29, Mutual Information. Analyzing Neural Time Series Data: Theory and Practice; MIT Press: Cambridge, MA, USA; London, UK, 2014; pp. 389–407. [Google Scholar]
- Nichol, J.M.; Seaver, M.; Trickey, S.T. A method for detecting damage-induced nonlinearities in structures using information theory. J. Sound. Vib. 2007, 297, 1–16. [Google Scholar] [CrossRef]
- Govindan, R.B.; Rathjen, J.; Kopper, F.; Claussen, J.C.; Deuschl, G. Estimation of time delay by coherence analysis. Phys. A Stat. Mech. Appl. 2005, 350, 277–295. [Google Scholar] [CrossRef] [Green Version]
- Muller, T.; Lauk, M.; Reinhard, M.; Hetzel, A.; Lucking, C.H.; Timmer, J. Estimation of Delay times in Biological systems. Ann. Biomed. Eng. 2003, 31, 1423–1439. [Google Scholar] [CrossRef] [Green Version]
- Nicholas Riddle, C.; Stuart, N.B. Manipulation of peripheral neural feedback loops alters human corticomuscular coherence. J. Physiol. 2005, 566, 625–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreas, W.; Marc de, L.; Markus, L. Time-delay Mutual Information of the phase as a Measure of Functional Connectivity. PLoS ONE 2012, 7, 1–22. [Google Scholar] [CrossRef]
- Andrew, M.F.; Harry, L.S. Independent coordinates for strange attractors from mutual information. A Gen. Phys. 1986, 33, 1134–1140. [Google Scholar]
- Laine, C.M.; Yavuz, S.U.; Farina1, D. Task-related changes in sensorimotor integration influence the common synaptic input to motor neurones. Acta Physiol. 2014, 211, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Christopher, M.; de Lussanet, M.; Valero-Cuevas, J. Intermuscular coherence reflects functional coordination. J. Neurophys. 2017, 118, 1775–1783. [Google Scholar] [CrossRef]
- Gert, P.; Lopes da Silva, F.H. Event-related EEG/MEG synchronization and desynchronization: Basic principles. Clin. Neurophys. 1999, 110, 1842–1857. [Google Scholar]
- Stephan, S.; Riitta, S.; Christa, N.; Gert, P.; Riitta, H. Human cortical 40 Hz rhythms is closely related to EMG rhythmicity. Neurosci. Lett. 1996, 213, 75–78. [Google Scholar]
- Pascal, F. A mechanism for cognitive dynamics: A neuronal communication through neuronal coherence. Trends Cogn. Sci. 2006, 9, 474–480. [Google Scholar]
- Tatsuya, M.; Nicole, S.; Tomi, O.; Mark, H. Force level modulates human cortical oscillatory activities. Neurosci. Lett. 1999, 275, 77–88. [Google Scholar]
- Jacobs, J.V.; Wu, G.; Kelly, K.M. Evidence for beta corticomuscular coherence during human standing balance: Effects of stance width, vision, and support surface. Neuroscience 2015, 298, 1–11. [Google Scholar] [CrossRef] [PubMed]
- David, P.; Juliana, D. Non-linear EEG dynamic changes and their probable relation to voluntary movement organization. Cog. Neu. Neu. Rep. 1999, 10, 1397–1401. [Google Scholar] [CrossRef]
- Tatsuya, M.; Takahiro, M.; Mark, H. Information flow from the sensorimotor cortex to muscles in humans. Clin. Neurophysiol. 2001, 112, 122–126. [Google Scholar]
- Panzica, F.; Canafoglia, L.; Franceschetti, S.; Binelli, S.; Ciano, C.; Visani, E.; Avanzini, G. Movement-activated myoclonus in genetically defined progressive myoclonic epilepsies: EEG-EMG relationship estimated using autoregressive models. Clin. Neurophy. 2003, 114, 1041–1052. [Google Scholar] [CrossRef]
- Firas, S.M.; Mohammed, D.; Shahab, A.; Siuly, S.; Jonathan, H.G.; Ravinesh, C.D. A new framework for classification of multi-category hand grasps using EMG signals. Art. Intel. Med. 2021, 112, 102005. [Google Scholar]
- Robert, H.; Larrabee, M.G.; German, W. The human electromyogram in response to nerve stimulation and the conduction velocity of motor axons: Studies on normal and on injured peripheral nerves. Arch. Neurol. Psychiatr. 1948, 60, 340–365. [Google Scholar] [CrossRef]
- Fay, B.H. Assumption underlying motor control for neurologic rehabilitation, Contemporary Management of Motor Problems, R.S.D. Neurol. Scien. Instit. 1991, 4, 11–24. [Google Scholar]
- Giovanni, T.; Micheal, J.; Jenny, V.S.J.; WooSurng, L.; Devin, D.; Dan, J.Z.; Eric, R.W.; Paul, A.E. Limitations of surface EMG estimate of parasternal intercostal to infer neural respiratory drive. Respir. Physiol. Neurobiol. 2021, 285, 1–9. [Google Scholar]












| Delay Time (ms) Obtained by Maximizing Mutual Information 1 | ||||||||
|---|---|---|---|---|---|---|---|---|
| Subject | RM | Inten | MI | OL | ||||
| β | γ | β | γ | β | γ | β | γ | |
| 1 | 20 | 15 | 20 | 20 | 40 | 15 | 45 | 30 |
| 2 | 20 | 15 | 20 | 20 | 30 | 20 | 45 | 39 |
| 3 | 25 | 20 | 25 | 20 | 25 | 45 | 35 | 35 |
| 4 | 30 | 23 | 25 | 25 | 30 | 28 | 35 | 40 |
| 5 | 25 | 30 | 0 | 30 | 35 | 30 | 20 | 39 |
| 6 | 20 | 30 | 35 | 15 | 25 | 30 | 43 | 25 |
| 7 | 25 | 20 | 35 | 20 | 40 | 20 | 43 | 20 |
| 8 | 30 | 0 | 25 | 20 | 30 | 30 | 35 | 25 |
| 9 | 15 | 15 | 30 | 25 | 40 | 28 | 35 | 20 |
| 10 | 25 | 30 | 15 | 30 | 35 | 35 | 25 | 25 |
| 11 | 16 | 23 | 35 | 28 | 39 | 43 | 43 | 39 |
| 12 | 20 | 15 | 30 | 30 | 35 | 40 | 25 | 20 |
| 13 | 25 | 15 | 30 | 0 | 30 | 45 | 30 | 15 |
| Mean | 22.76 | 19.31 | 25.00 | 21.76 | 33.38 | 31.46 | 35.31 | 28.61 |
| SD | 4.83 | 8.35 | 9.78 | 8.16 | 5.45 | 9.78 | 8.37 | 8.86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tun, N.N.; Sanuki, F.; Iramina, K. Electroencephalogram-Electromyogram Functional Coupling and Delay Time Change Based on Motor Task Performance. Sensors 2021, 21, 4380. https://doi.org/10.3390/s21134380
Tun NN, Sanuki F, Iramina K. Electroencephalogram-Electromyogram Functional Coupling and Delay Time Change Based on Motor Task Performance. Sensors. 2021; 21(13):4380. https://doi.org/10.3390/s21134380
Chicago/Turabian StyleTun, Nyi Nyi, Fumiya Sanuki, and Keiji Iramina. 2021. "Electroencephalogram-Electromyogram Functional Coupling and Delay Time Change Based on Motor Task Performance" Sensors 21, no. 13: 4380. https://doi.org/10.3390/s21134380