Highly Sensitive Detection of Chemically Modified Thio-Organophosphates by an Enzymatic Biosensing Device: An Automated Robotic Approach

 , ,
, ,  and
and
Abstract
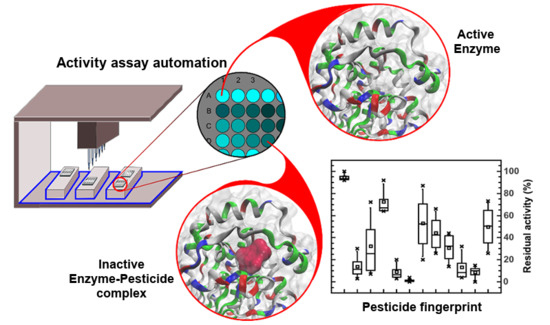
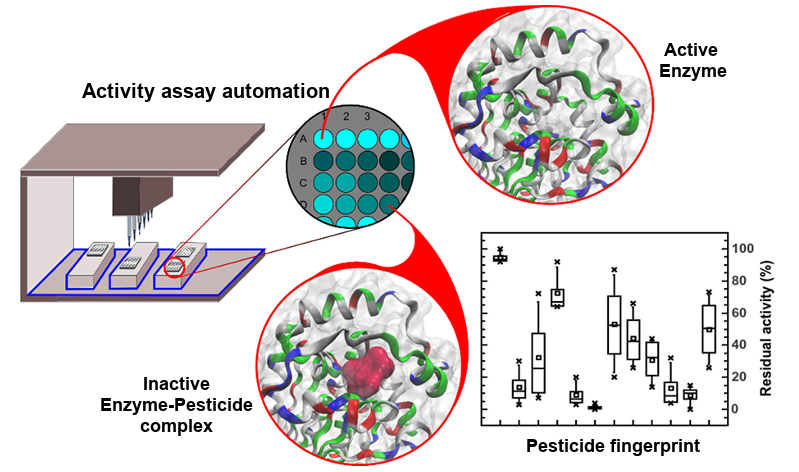
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Enzyme Purification
2.3. Fluorescence Standard Enzymatic Assay
2.4. Inhibition Assay of EST2 in Presence of Pesticides
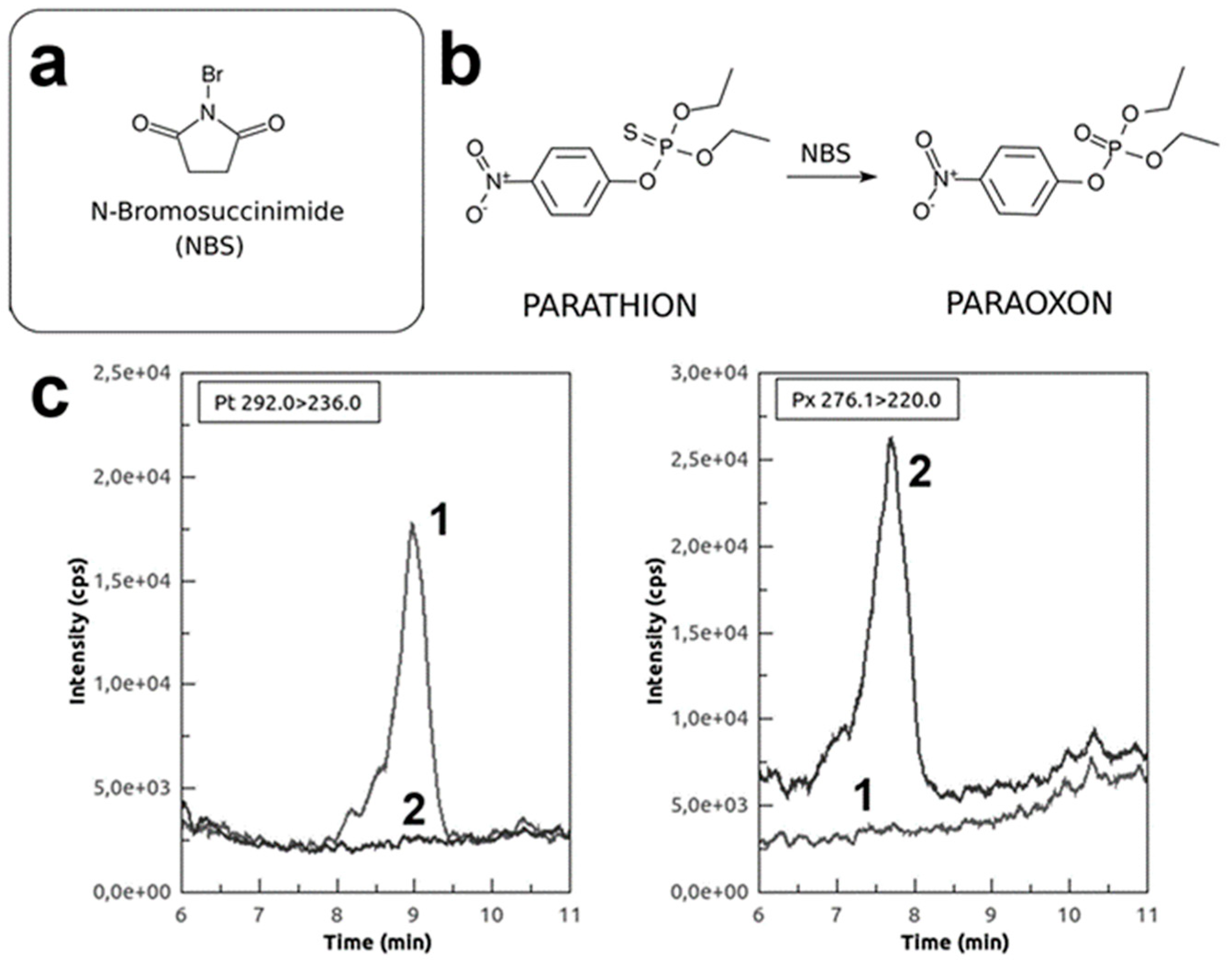
2.5. Phosphorothionate Pesticide Oxidation by NBS
2.6. MS Analysis of NBS-Oxidised Pesticides
2.7. Determination of EST2 Activity in Presence of NBS
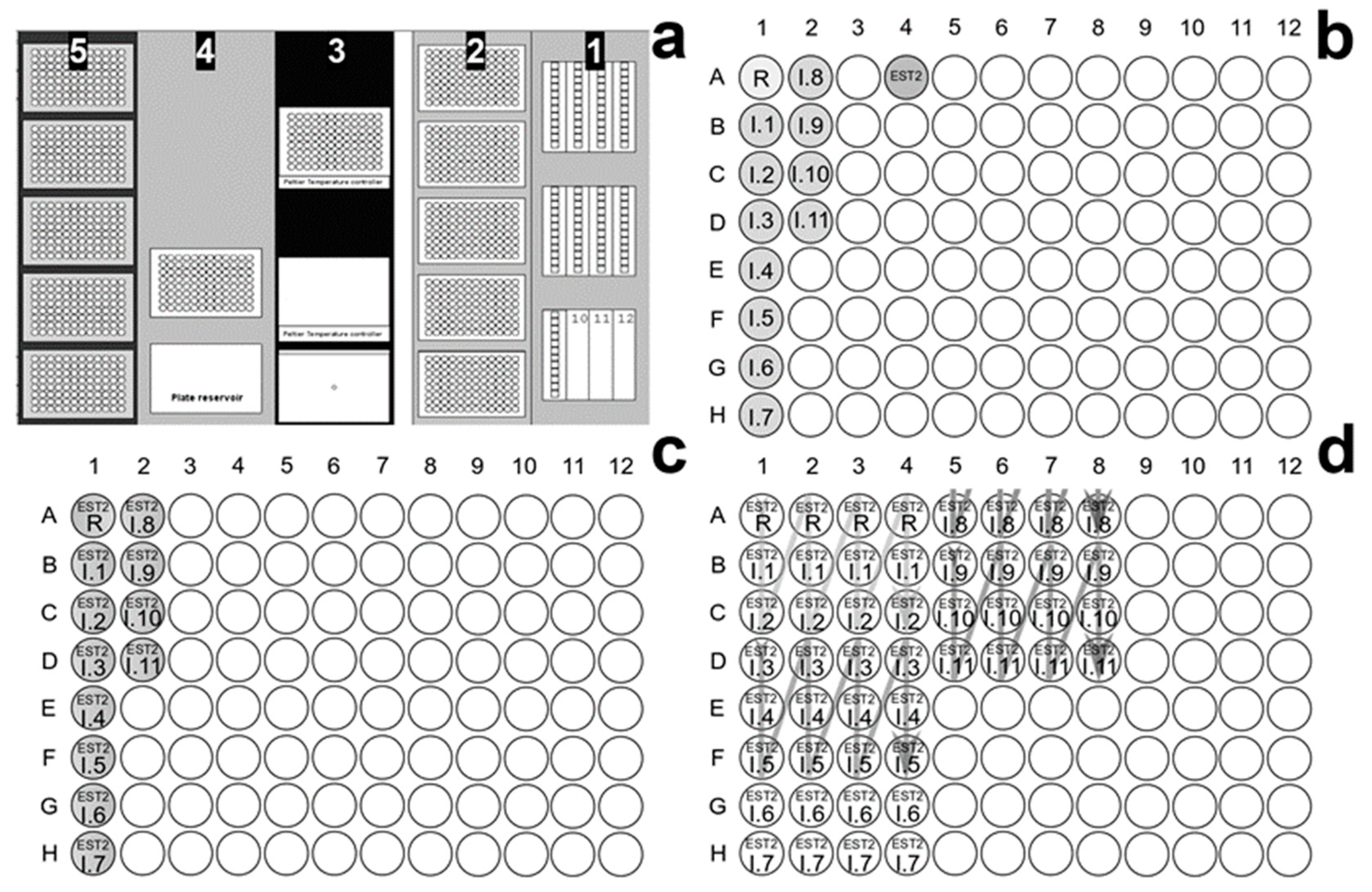
2.8. EST2 Assay on Robotic Workstation and OP Screening
3. Results and Discussion
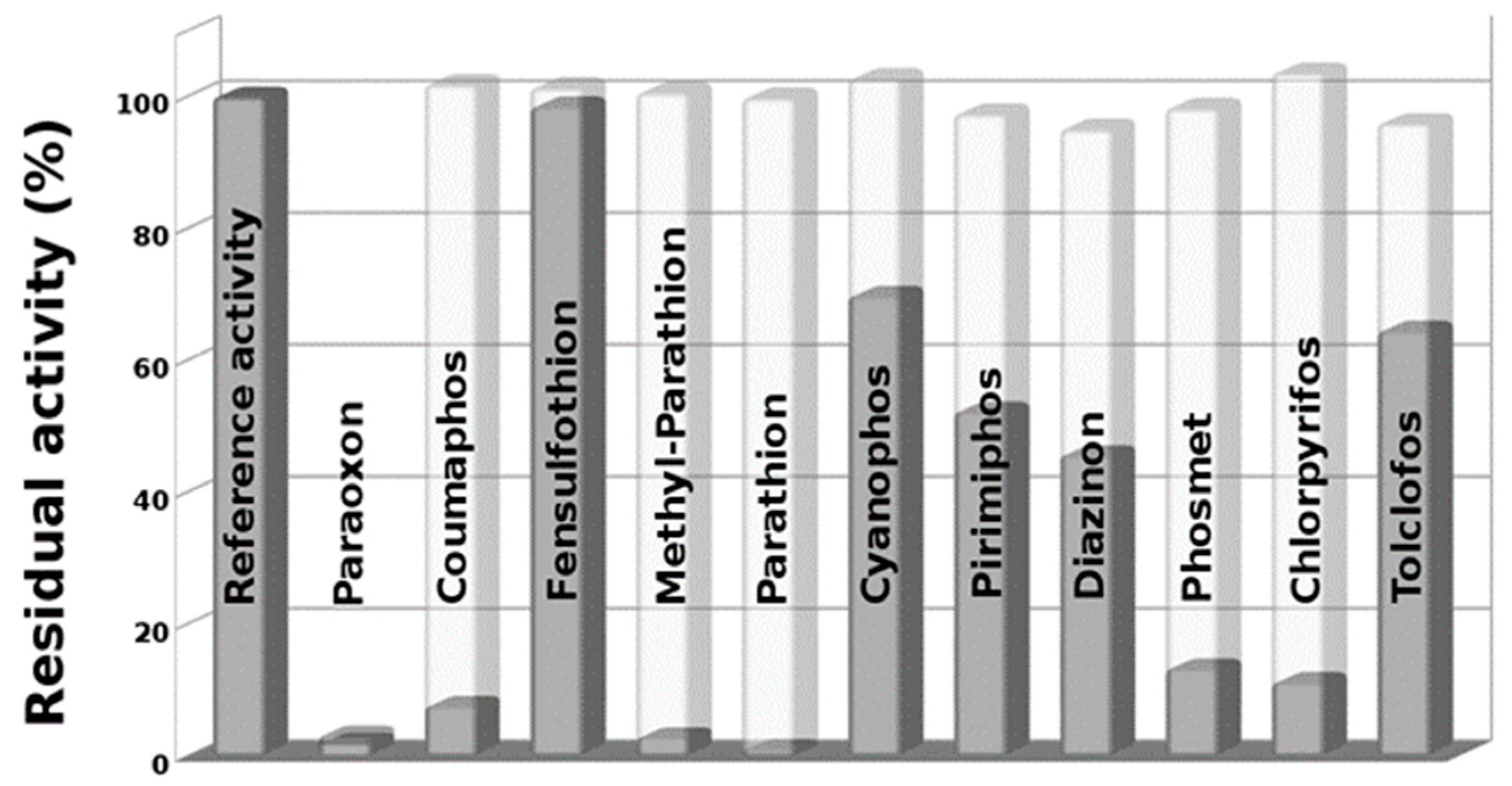
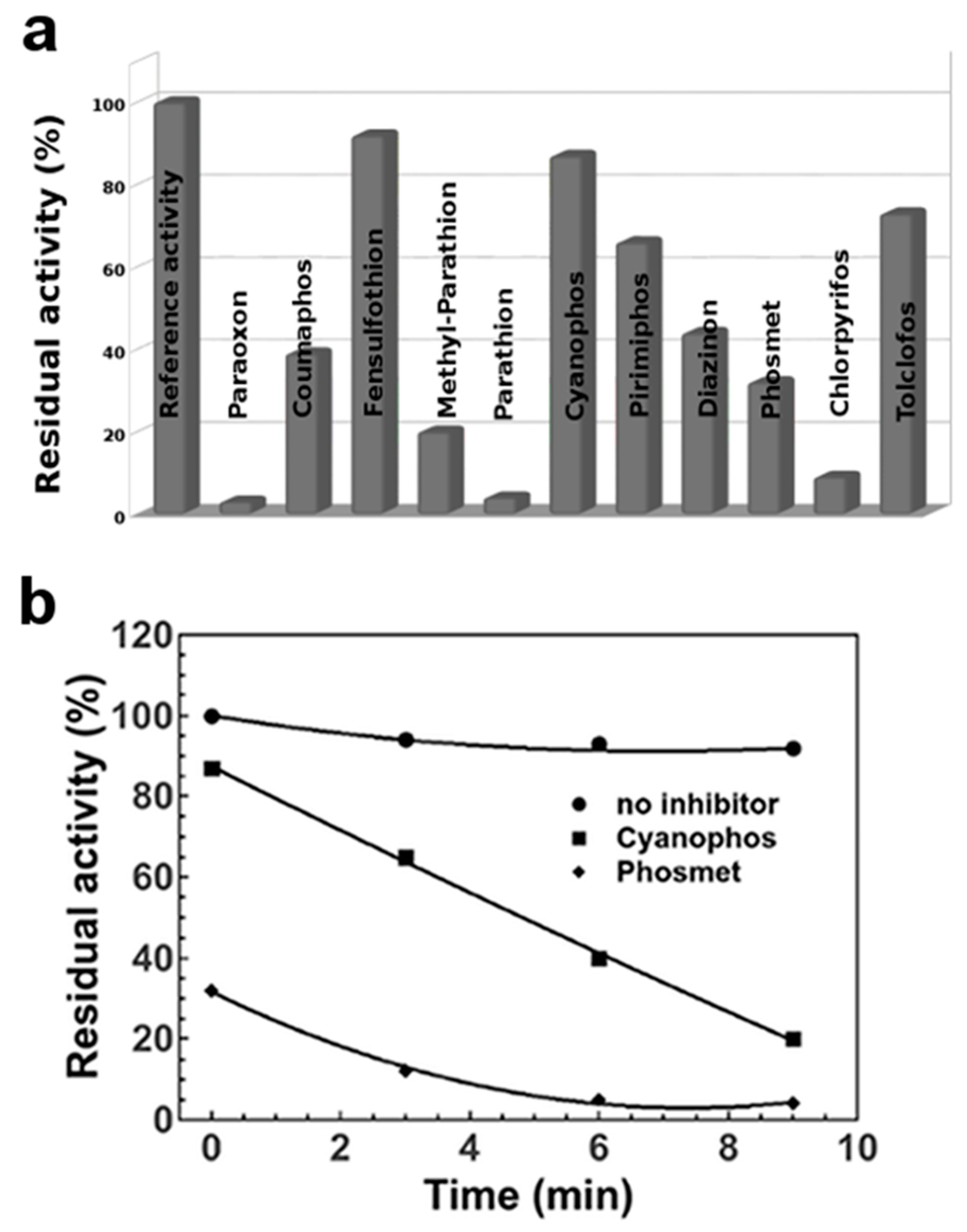
3.1. In Vitro Pesticides Activation by Using NBS
3.2. Effects of NBS on EST2 Activity
3.3. EST2 Inhibition Measurements on in Vitro NBS-Activated Pesticides
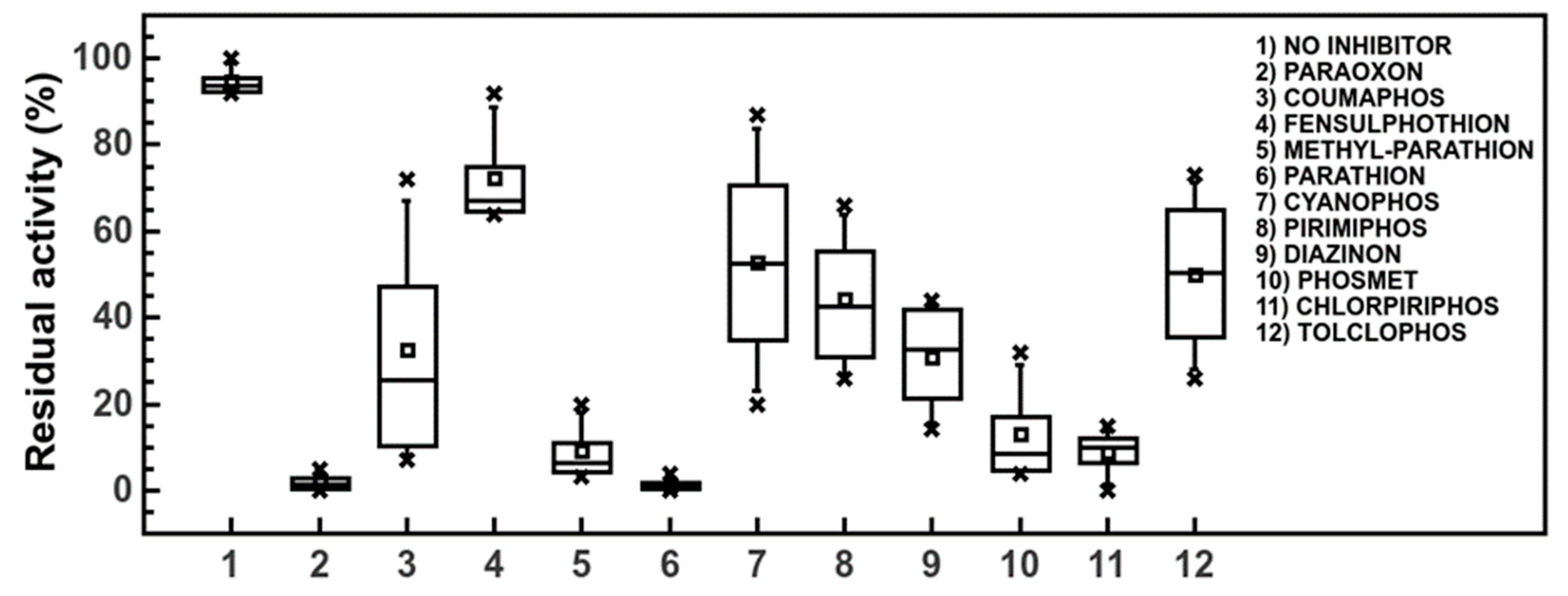
3.4. Automatic Approach in the Pesticide Determinations by Robotic System
3.5. Pesticide Pseudo Finger-Print
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bjørling-Poulsen, M.; Andersen, H.; Grandjean, P. Potential developmental neurotoxicity of pesticides used in Europe. Environ. Health 2008, 7, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- London, L.; Beseler, C.; Bouchard, M.F.; Bellinger, D.C.; Colosio, C.; Grandjean, P.; Harari, R.; Kootbodien, T.; Kromhout, H.; Little, F.; et al. Neurobehavioral and neurodevelopmental effects of pesticide exposures. Neurotoxicology 2012, 33, 887–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beard, J.D.; Hoppin, J.A.; Richards, M.; Alavanja, M.C.R.; Blair, A.; Sandler, D.P.; Kamel, F. Pesticide exposure and self-reported incident depression among wives in the Agricultural Health Study. Environ. Res. 2013, 126, 31–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, C.A.; Wilkinson, S.C.; Blain, P.G.; Dunn, M.; Aust, G.A.; Williams, F.M. Percutaneous absorption and distribution of organophosphates (chlorpyrifos and dichlorvos) following dermal exposure and decontamination scenarios using in vitro human skin model. Toxicol. Lett. 2014, 229, 66–72. [Google Scholar] [CrossRef]
- Furlong, M.A.; Engel, S.M.; Barr, D.B.; Wolff, M.S. Prenatal exposure to organophosphate pesticides and reciprocal social behavior in childhood. Environ. Int. 2014, 70, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Rauh, V.A.; Garfinkel, R.; Perera, F.P.; Andrews, H.F.; Hoepner, L.; Barr, D.B.; Whitehead, R.; Tang, D.; Whyatt, R.W. Impact of prenatal chlorpyrifos exposure on neurodevelopment in the first 3 years of life among inner-city children. Pediatrics 2006, 118, e1845–e1859. [Google Scholar] [CrossRef] [Green Version]
- Rauh, V.; Arunajadai, S.; Horton, M.; Perera, F.; Hoepner, L.; Barr, D.B.; Whyatt, R. Seven-year neurodevelopmental scores and prenatal exposure to chlorpyrifos, a common agricultural pesticide. Environ. Health Perspect. 2011, 119, 1196–1201. [Google Scholar] [CrossRef]
- Febbraio, F. Biochemical strategies for the detection and detoxification of toxic chemicals in the environment. World J. Biol. Chem. 2017, 8, 13–20. [Google Scholar] [CrossRef]
- Gervais, G.; Brosillon, S.; Laplanche, A.; Helen, C. Ultra-pressure liquid chromatography-electrospray tandem mass spectrometry for multiresidue determination of pesticides in water. J. Chromatogr. A 2008, 1202, 163–172. [Google Scholar] [CrossRef]
- Hengel, M.J.; Miller, M. Analysis of pesticides in dried hops by liquid chromatography-tandem mass spectrometry. J. Agric. Food Chem. 2008, 56, 6851–6856. [Google Scholar] [CrossRef]
- Manco, G.; Nucci, R.; Febbraio, F. Use of esterase activities for the detection of chemical neurotoxic agents. Protein Pept. Lett. 2009, 16, 1225–1234. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.K.; Dominguez, R.B.; Bhand, S.; Muñoz, R.; Marty, J.-L. A novel automated flow-based biosensor for the determination of organophosphate pesticides in milk. Biosens. Bioelectron. 2012, 32, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Pundir, C.S.; Chauhan, N. Acetylcholinesterase inhibition-based biosensors for pesticide determination: A review. Anal. Biochem. 2012, 429, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zheng, Z.; Liu, X.; Zhao, S.; Liu, S. Nanostructured photoelectrochemical biosensor for highly sensitive detection of organophosphorous pesticides. Biosens. Bioelectron. 2015, 64, 1–5. [Google Scholar] [CrossRef]
- Hassani, S.; Momtaz, S.; Vakhshiteh, F.; Maghsoudi, A.S.; Ganjali, M.R.; Norouzi, P.; Abdollahi, M. Biosensors and their applications in detection of organophosphorus pesticides in the environment. Arch. Toxicol. 2017, 91, 109–130. [Google Scholar] [CrossRef]
- Whangsuk, W.; Thiengmag, S.; Dubbs, J.; Mongkolsuk, S.; Loprasert, S. Specific detection of the pesticide chlorpyrifos by a sensitive genetic-based whole cell biosensor. Anal. Biochem. 2016, 493, 11–13. [Google Scholar] [CrossRef]
- Warner, J.; Andreescu, S. An acetylcholinesterase (AChE) biosensor with enhanced solvent resistance based on chitosan for the detection of pesticides. Talanta 2016, 146, 279–284. [Google Scholar] [CrossRef]
- Guler, M.; Turkoglu, V.; Kivrak, A. Electrochemical detection of malathion pesticide using acetylcholinesterase biosensor based on glassy carbon electrode modified with conducting polymer film. Environ. Sci. Pollut. Res. Int. 2016, 23, 12343–12351. [Google Scholar] [CrossRef]
- Febbraio, F.; Merone, L.; Cetrangolo, G.P.; Rossi, M.; Nucci, R.; Manco, G. Thermostable esterase 2 from Alicyclobacillus acidocaldarius as biosensor for the detection of organophosphate pesticides. Anal. Chem. 2011, 83, 1530–1536. [Google Scholar] [CrossRef]
- De Simone, G.; Mandrich, L.; Menchise, V.; Giordano, V.; Febbraio, F.; Rossi, M.; Pedone, C.; Manco, G. A substrate-induced switch in the reaction mechanism of a thermophilic esterase—Kinetic evidences and structural basis. J. Biol. Chem. 2004, 279, 6815–6823. [Google Scholar] [CrossRef]
- Febbraio, F.; D’Andrea, S.E.; Mandrich, L.; Merone, L.; Rossi, M.; Nucci, R.; Manco, G. Irreversible inhibition of the thermophilic esterase EST2 from Alicyclobacillus acidocaldarius. Extremophiles 2008, 12, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Cetrangolo, G.P.; Gori, C.; Rusko, J.; Terreri, S.; Manco, G.; Cimmino, A.; Febbraio, F. Determination of Picomolar Concentrations of Paraoxon in Human Urine by Fluorescence-Based Enzymatic Assay. Sensors 2019, 19, 4582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bavcon Kralj, M.; Trebse, P.; Franko, M. Oxidation as a Pre-step in Determination of Organophosphorus Compounds by the AChE-TLS Bioassay. Acta Chimica Slovenica 2006, 53, 43–51. [Google Scholar]
- Schulze, H.; Schmid, R.D.; Bachmann, T.T. Activation of phosphorothionate pesticides based on a cytochrome P450 BM-3 (CYP102 A1) mutant for expanded neurotoxin detection in food using acetylcholinesterase biosensors. Anal. Chem. 2004, 76, 1720–1725. [Google Scholar] [CrossRef] [PubMed]
- Fukuto, T.R. Mechanism of action of organophosphorus and carbamate insecticides. Environ. Health Perspect. 1990, 87, 245–254. [Google Scholar] [CrossRef]
- Manco, G.; Adinolfi, E.; Pisani, F.M.; Ottolina, G.; Carrea, G.; Rossi, M. Overexpression and properties of a new thermophilic and thermostable esterase from Bacillus acidocaldarius with sequence similarity to hormone-sensitive lipase subfamily. Biochem. J. 1998, 332, 203–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sotiropoulou, S.; Fournier, D.; Chaniotakis, N.A. Genetically engineered acetylcholinesterase-based biosensor for attomolar detection of dichlorvos. Biosens. Bioelectron. 2005, 20, 2347–2352. [Google Scholar] [CrossRef]
- Carullo, P.; Cetrangolo, G.P.; Mandrich, L.; Manco, G.; Febbraio, F. Fluorescence spectroscopy approaches for the development of a real-time organophosphate detection system using an enzymatic sensor. Sensors 2015, 15, 3932–3951. [Google Scholar] [CrossRef] [Green Version]
- Carullo, P.; Chino, M.; Cetrangolo, G.P.; Terreri, S.; Lombardi, A.; Manco, G.; Cimmino, A.; Febbraio, F. Direct detection of organophosphate compounds in water by a fluorescence-based biosensing device. Sens. Actuators B Chem. 2018, 255, 3257–3266. [Google Scholar] [CrossRef]
- Del Carlo, M.; Pepe, A.; Sergi, M.; Mascini, M.; Tarentini, A.; Compagnone, D. Detection of coumaphos in honey using a screening method based on an electrochemical acetylcholinesterase bioassay. Talanta 2010, 81, 76–81. [Google Scholar] [CrossRef]
- Hou, C.; He, K.; Yang, L.; Huo, D.; Yang, M.; Huang, S.; Zhang, L.; Shen, C. Catalytic characteristics of plant-esterase from wheat flour. World J. Microbiol. Biotechnol. 2012, 28, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Dixon, M.; Webb, E.C. Enzymes, 3rd ed.; Longman Group Limited London: New York, NY, USA, 1979; ISBN 0-12-218358-4. [Google Scholar]
- Krstić, D.; Colović, M.; Krinulović, K.; Djurić, D.; Vasić, V. Inhibition of AChE by single and simultaneous exposure to malathion and its degradation products. Gen. Physiol. Biophys. 2007, 26, 247–253. [Google Scholar] [PubMed]
- Caserta, G.; Chino, M.; Firpo, V.; Zambrano, G.; Leone, L.; D’Alonzo, D.; Nastri, F.; Maglio, O.; Pavone, V.; Lombardi, A. Enhancement of Peroxidase Activity in Artificial Mimochrome VI Catalysts through Rational Design. Chembiochem 2018, 19, 1823–1826. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, J.; Robledo, N.R.; Velasco, L.; Quintero, R.; Pickard, M.A.; Vazquez-Duhalt, R. Chloroperoxidase-Mediated Oxidation of Organophosphorus Pesticides. Pestic. Biochem. Physiol. 1998, 61, 87–94. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cetrangolo, G.P.; Rusko, J.; Gori, C.; Carullo, P.; Manco, G.; Chino, M.; Febbraio, F. Highly Sensitive Detection of Chemically Modified Thio-Organophosphates by an Enzymatic Biosensing Device: An Automated Robotic Approach. Sensors 2020, 20, 1365. https://doi.org/10.3390/s20051365
Cetrangolo GP, Rusko J, Gori C, Carullo P, Manco G, Chino M, Febbraio F. Highly Sensitive Detection of Chemically Modified Thio-Organophosphates by an Enzymatic Biosensing Device: An Automated Robotic Approach. Sensors. 2020; 20(5):1365. https://doi.org/10.3390/s20051365
Chicago/Turabian StyleCetrangolo, Giovanni Paolo, Janis Rusko, Carla Gori, Paola Carullo, Giuseppe Manco, Marco Chino, and Ferdinando Febbraio. 2020. "Highly Sensitive Detection of Chemically Modified Thio-Organophosphates by an Enzymatic Biosensing Device: An Automated Robotic Approach" Sensors 20, no. 5: 1365. https://doi.org/10.3390/s20051365