Feasibility of Volatile Biomarker-Based Detection of Pythium Leak in Postharvest Stored Potato Tubers Using Field Asymmetric Ion Mobility Spectrometry

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Potato Tubers
2.2. Inoculum Preparation and Inoculation
2.3. Experimental Design
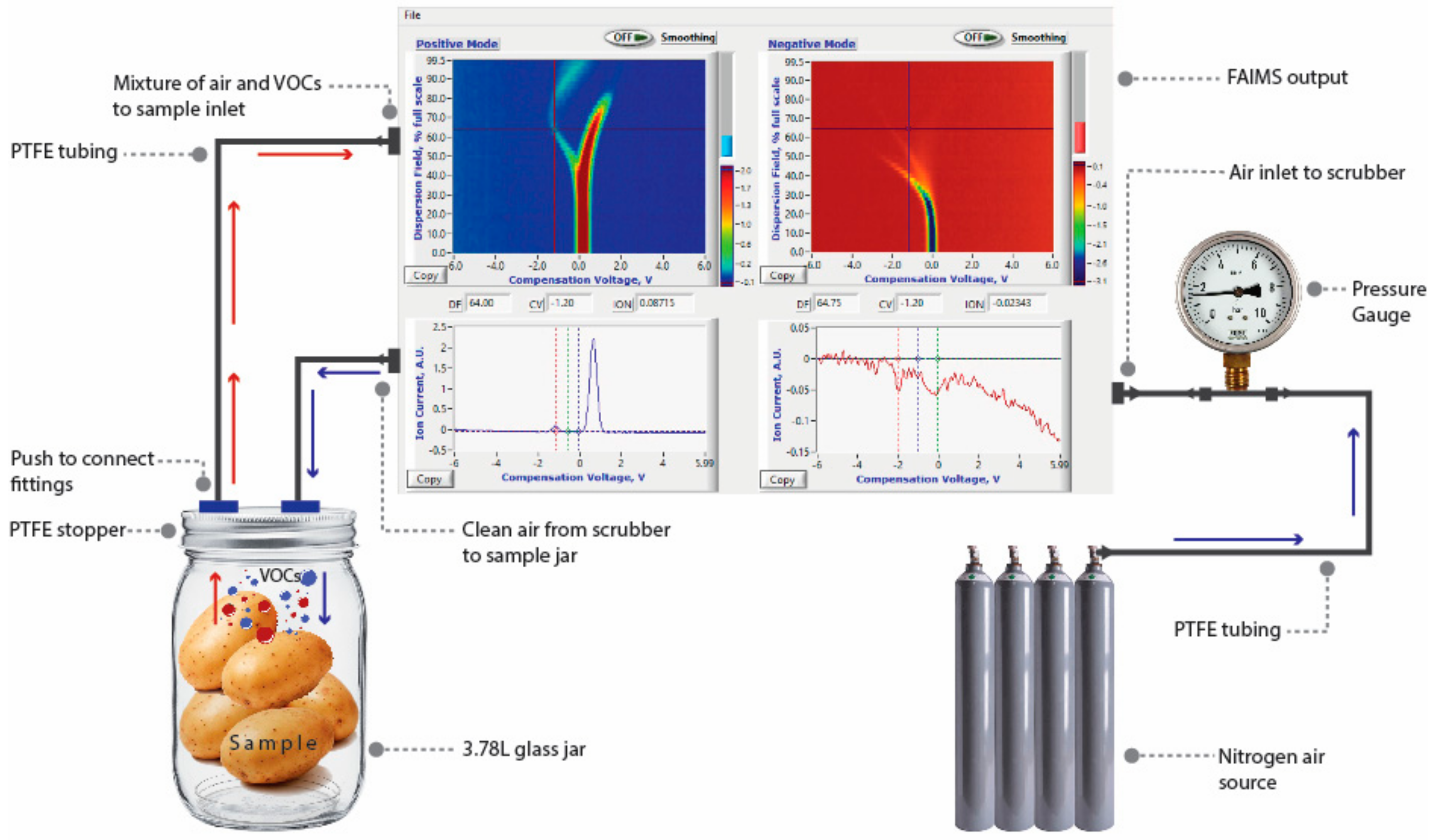
2.4. FAIMS Evaluations
2.5. Data Analysis
3. Results and Discussion
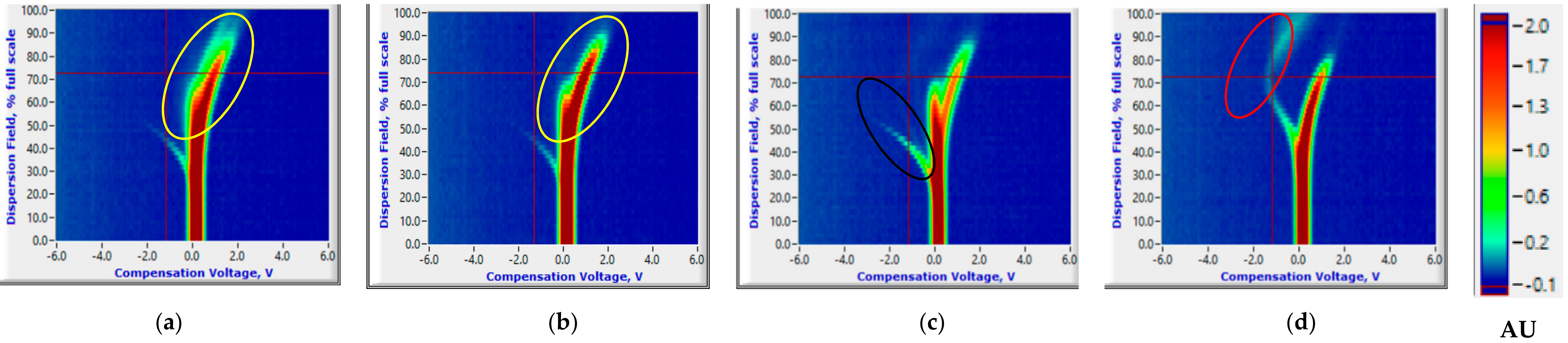
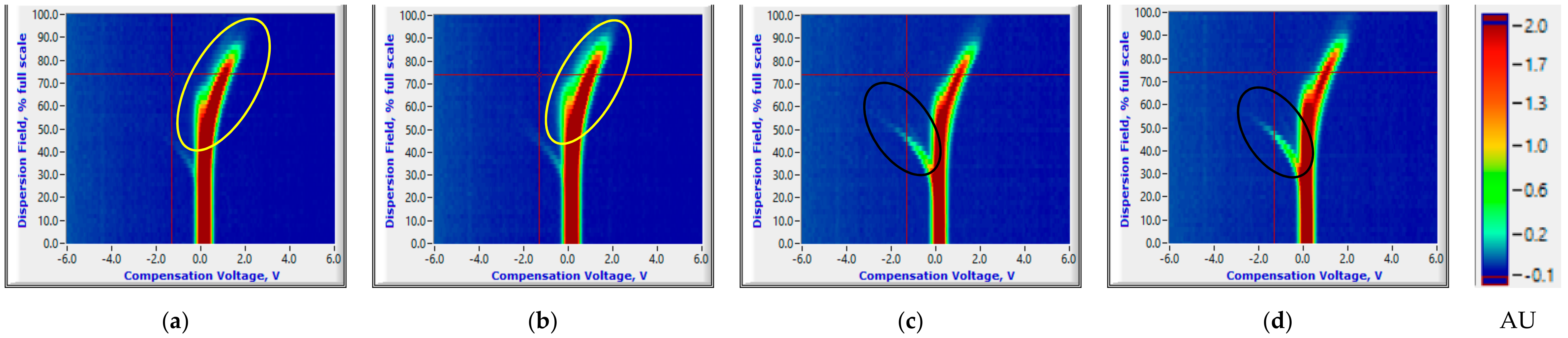
3.1. Salient FAIMS Signatures
3.1.1. Room Temperature Storage Experiments
3.1.2. Reduced Temperature Storage Experiments
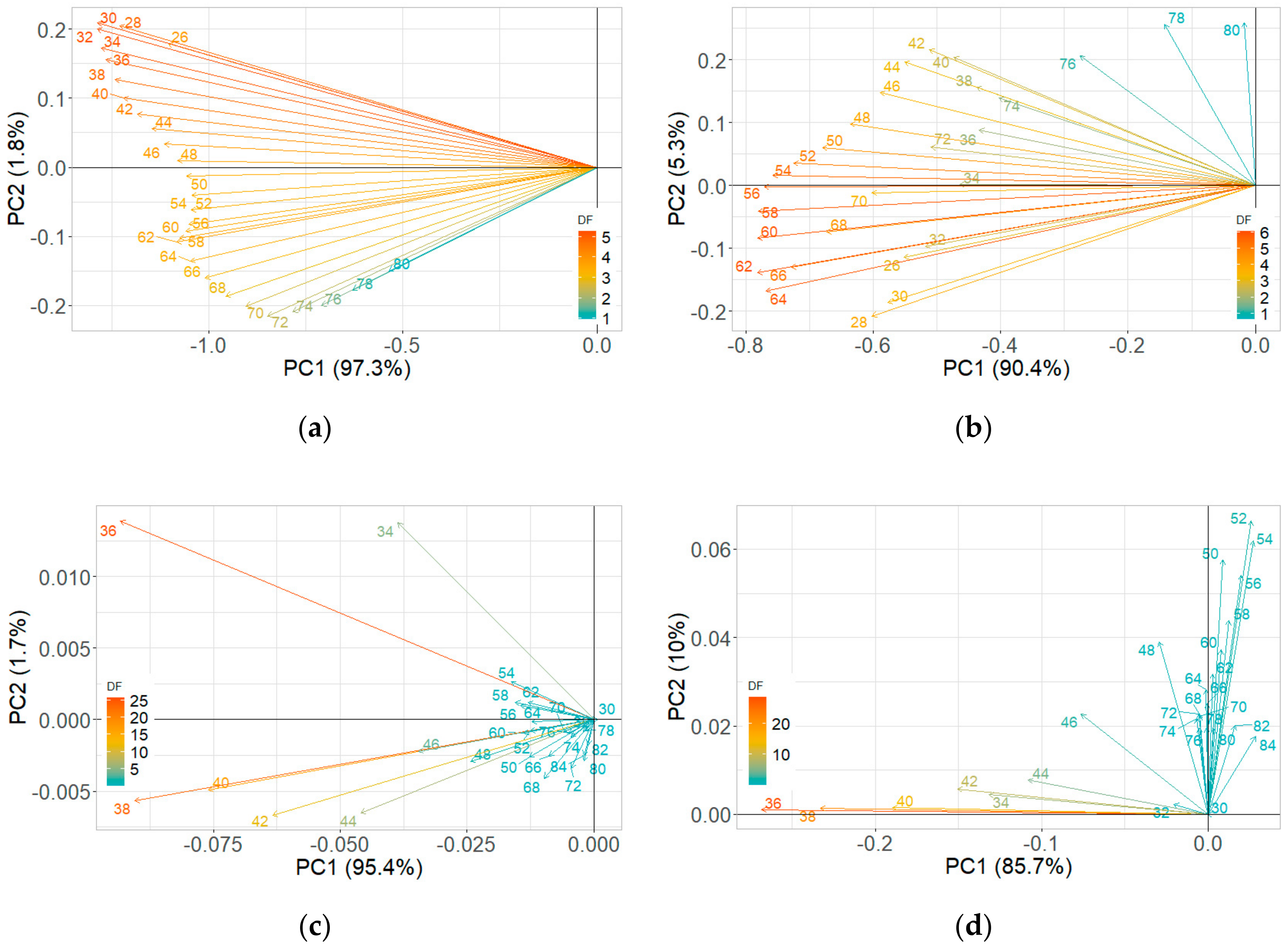
3.2. Pattern Recognition
3.2.1. Room Temperature Storage Experiments
3.2.2. Reduced Temperature Storage Experiments
3.3. Temporal Progression
3.3.1. Room Temperature Storage Experiments
3.3.2. Reduced Temperature Storage Experiments
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Uitdewilligen, J. Discovery and Genotyping of Existing and Induced DNA Sequence Variation in Potato; Wageningen University: Wageningen, The Netherlands, 2012. [Google Scholar]
- Johnson, D.A. Potato Health Management; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2008; p. 261. [Google Scholar]
- Gachango, E. Management of Postharvest Diseases of Potato (Solanum tuberosum L.); Plant Pathology, Michigan State University: East Lansing, MI, USA, 2011. [Google Scholar]
- Powelson, M.; Rowe, R. Managing diseases caused by seedborne and soilborne fungi and fungus-like pathogens. Potato Health Manag. 2008, 2183–2195. [Google Scholar]
- Stark, J.C.; Love, S.L.; Knowles, N.R. Tuber Quality. In Potato Production Systems; Springer: Berlin/Heidelberg, Germany, 2020; pp. 479–497. [Google Scholar]
- Ammar, N.; Jabnoun-Khiareddine, H.; Mejdoub-Trabelsi, B.; Nefzi, A.; Mahjoub, M.A.; Daami-Remadi, M. Pythium leak control in potato using aqueous and organic extracts from the brown alga Sargassum vulgare (C. Agardh, 1820). Postharvest Biol. Technol. 2017, 130, 81–93. [Google Scholar] [CrossRef]
- Daami-Remadi, M.; Dkhili, I.; Jabnoun-Khiareddine, H.; El Mahjoub, M. Biological control of potato leak with antagonistic fungi isolated from compost teas and solarized and non-solarized soils. Pest. Technol. 2012, 6, 32–40. [Google Scholar]
- Olsen, N.; Miller, J.; Nolte, P. Diagnosis & Management of Potato Storage Diseases; Idaho Agricultural Experiment Station: Moscow, ID, USA, 2006. [Google Scholar]
- Wang, T.; Gao, C.; Cheng, Y.; Li, Z.; Chen, J.; Guo, L.; Xu, J. Molecular Diagnostics and Detection of Oomycetes on Fiber Crops. Plants 2020, 9, 769. [Google Scholar] [CrossRef] [PubMed]
- Pettitt, T.; Wakeham, A.; Wainwright, M.; White, J. Comparison of serological, culture, and bait methods for detection of Pythium and Phytophthora zoospores in water. Plant Pathol. 2002, 51, 720–727. [Google Scholar] [CrossRef]
- Shen, D.; Li, Q.; Yu, J.; Zhao, Y.; Zhu, Y.; Xu, H.; Dou, D. Development of a loop-mediated isothermal amplification method for the rapid detection of Pythium ultimum. Australas. Plant Pathol. 2017, 46, 571–576. [Google Scholar] [CrossRef]
- Schroeder, K.L.; Martin, F.N.; de Cock, A.W.; Lévesque, C.A.; Spies, C.F.; Okubara, P.A.; Paulitz, T.C. Molecular detection and quantification of Pythium species: Evolving taxonomy, new tools, and challenges. Plant Dis. 2013, 97, 4–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, C.; Vitalis, E.A.; Baker, B.R.; Gardner, S.N.; Torres, M.W.; Dzenitis, J.M. LAVA: An open-source approach to designing LAMP (loop-mediated isothermal amplification) DNA signatures. BMC Bioinform. 2011, 12, 240. [Google Scholar] [CrossRef] [Green Version]
- Toth, I.K.; Bell, K.S.; Holeva, M.C.; Birch, P.R. Soft rot erwiniae: From genes to genomes. Mol. Plant Pathol. 2003, 4, 17–30. [Google Scholar] [CrossRef]
- Atallah, Z.; Stevenson, W. A methodology to detect and quantify five pathogens causing potato tuber decay using real-time quantitative polymerase chain reaction. Phytopathology 2006, 96, 1037–1045. [Google Scholar] [CrossRef] [Green Version]
- Cullen, D.; Toth, I.; Boonham, N.; Walsh, K.; Barker, I.; Lees, A. Development and validation of conventional and quantitative polymerase chain reaction assays for the detection of storage rot potato pathogens, Phytophthora erythroseptica, Pythium ultimum and Phoma foveata. J. Phytopathol. 2007, 155, 309–315. [Google Scholar] [CrossRef]
- Lui, L.; Kushalappa, A. Models to predict potato tuber infection by Pythium ultimum from duration of wetness and temperature, and leak-lesion expansion from storage duration and temperature. Postharvest Biol. Technol. 2003, 27, 313–322. [Google Scholar] [CrossRef]
- Jansen, R.; Wildt, J.; Kappers, I.; Bouwmeester, H.; Hofstee, J.; Van Henten, E. Detection of diseased plants by analysis of volatile organic compound emission. Annu. Rev. Phytopathol. 2011, 49, 157–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, R.; Khot, L.R.; Schroeder, B.K.; Sankaran, S. FAIMS based volatile fingerprinting for real-time postharvest storage infections detection in stored potatoes and onions. Postharvest Biol. Technol. 2018, 135, 83–92. [Google Scholar] [CrossRef]
- Cellini, A.; Biondi, E.; Buriani, G.; Farneti, B.; Rodriguez-Estrada, M.T.; Braschi, I.; Savioli, S.; Blasioli, S.; Rocchi, L.; Biasioli, F. Characterization of volatile organic compounds emitted by kiwifruit plants infected with Pseudomonas syringae pv. actinidiae and their effects on host defences. Trees 2016, 30, 795–806. [Google Scholar] [CrossRef]
- Sinha, R.; Khot, L.R.; Schroeder, B.K.; Si, Y. Rapid and non–destructive detection of Pectobacterium carotovorum causing soft rot in stored potatoes through volatile biomarkers sensing. Crop. Prot. 2017, 93, 122–131. [Google Scholar] [CrossRef] [Green Version]
- Rutolo, M.; Covington, J.A.; Clarkson, J.; Iliescu, D. Detection of potato storage disease via gas analysis: A pilot study using field asymmetric ion mobility spectrometry. Sensors 2014, 14, 15939–15952. [Google Scholar] [CrossRef] [Green Version]
- Sinha, R.; Khot, L.R.; Schroeder, B.K. FAIMS based sensing of Burkholderia cepacia caused sour skin in onions under bulk storage condition. J. Food Meas. Charact. 2017, 11, 1578–1585. [Google Scholar] [CrossRef]
- Rutolo, M.; Clarkson, J.P.; Covington, J.A. The use of an electronic nose to detect early signs of soft-rot infection in potatoes. Biosyst. Eng. 2018, 167, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Chang, Z.; Lv, J.; Qi, H.; Ma, Y.; Chen, D.; Xie, J.; Sun, Y. Bacterial infection potato tuber soft rot disease detection based on electronic nose. Open Life Sci. 2017, 12, 379–385. [Google Scholar] [CrossRef]
- Chavarria, M.A.; Matheoud, A.V.; Marmillod, P.; Liu, Y.; Kong, D.; Brugger, J.; Boero, G. High sensitivity field asymmetric ion mobility spectrometer. Rev. Sci. Instrum. 2017, 88, 035115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feider, C.L.; Elizondo, N.; Eberlin, L.S. Ambient ionization and FAIMS mass spectrometry for enhanced imaging of multiply charged molecular ions in biological tissues. Anal. Chem. 2016, 88, 11533–11541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borsdorf, H.; Nazarov, E.G.; Miller, R.A. Time-of-flight ion mobility spectrometry and differential mobility spectrometry: A comparative study of their efficiency in the analysis of halogenated compounds. Talanta 2007, 71, 1804–1812. [Google Scholar] [CrossRef] [PubMed]
- Tufariello, J.A. Development of a Breathalyzer: Ion. Mobility Spectrometer for the Detection of Cannabis from Human Breath; Washington State University: Pullman, WA, USA, 2016. [Google Scholar]
- Shvartsburg, A.A. Differential Ion Mobility Spectrometry: Nonlinear ion Transport and Fundamentals of FAIMS; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Karpas, Z. Applications of ion mobility spectrometry (IMS) in the field of foodomics. Food Res. Int. 2013, 54, 1146–1151. [Google Scholar] [CrossRef]
- Costanzo, M.T.; Boock, J.J.; Kemperman, R.H.; Wei, M.S.; Beekman, C.R.; Yost, R.A. Portable FAIMS: Applications and future perspectives. Int. J. Mass Spectrom. 2017, 422, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Vernon, A.S.; Covington, J.A.; Arasaradnam, R.P.; Esfahani, S.; O’connell, N.; Kyrou, I.; Savage, R.S. An improved machine learning pipeline for urinary volatiles disease detection: Diagnosing diabetes. PLoS ONE 2018, 13, e0204425. [Google Scholar] [CrossRef]
- Song, X.J.; Beckman, T.; Scherm, H.; Kemenova, O.; McMurray, G. Detection of Armillaria Root Rot Disease by Analysis of Volatile Organic Compounds. In Proceedings of the 2015 ASABE Annual International Meeting, New Orleans, LA, USA, 26–29 July 2015; p. 1. [Google Scholar]
- Cui, S.; Ling, P.; Zhu, H.; Keener, H.M. Plant pest detection using an artificial nose system: A review. Sensors 2018, 18, 378. [Google Scholar] [CrossRef] [Green Version]
- Hollingshead, A.K.; Olsen, N.L.; Thornton, M.; Miller, J.S.; Lin, A.H.M. Potato cultivar susceptibility to Pythium leak as influenced by harvest and early storage temperatures. Am. J. Potato Res. 2020, 97, 393–403. [Google Scholar] [CrossRef]
- Zrodnikov, Y.; Rajapakse, M.Y.; Peirano, D.J.; Aksenov, A.A.; Kenyon, N.J.; Davis, C.E. High asymmetric longitudinal field ion mobility spectrometry device for low power mobile chemical separation and detection. Anal. Chem. 2019, 91, 5523–5529. [Google Scholar] [CrossRef]
- Przybylski, C.; Bonnet, V. Discrimination of isomeric trisaccharides and their relative quantification in honeys using trapped ion mobility spectrometry. Food Chem. 2020, 341, 128182. [Google Scholar] [CrossRef]
- Nema, P.K.; Ramayya, N.; Duncan, E.; Niranjan, K. Potato glycoalkaloids: Formation and strategies for mitigation. J. Sci. Food Agric. 2008, 88, 1869–1881. [Google Scholar] [CrossRef]
- Kuznetsova, M.; Rogozhin, A.; Smetanina, T.; Demidova, V.; Denisenkov, I.; Statsyuk, N. Pythium-Induced Root Rot of Potato and Its Control. Entomol. Appl. Sci. Lett. 2018, 5, 55–61. [Google Scholar]
- Voss, R.; Baghott, K.; Timm, H. Proper environment for potato storage. In Vegetable Research and Information Center; The University of California: Davis, CA, USA, 2001. [Google Scholar]
- Eiceman, G.; Nazarov, E.; Rodriguez, J.; Bergloff, J. Positive reactant ion chemistry for analytical, high temperature ion mobility spectrometry (IMS): Effects of electric field of the drift tube and moisture, temperature, and flow of the drift gas. Int. J. Ion Mobil. Spectrom 1998, 1, 28–37. [Google Scholar]
- Salas, B.; Secor, G.A.; Taylor, R.J.; Gudmestad, N.C. Assessment of resistance of tubers of potato cultivars to Phytophthora erythroseptica and Pythium ultimum. Plant Dis. 2003, 87, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Tedla, T.; Stanghellini, M. Bacterial population dynamics and interactions with Pythium aphanidermatum in intact rhizosphere soil. Phytopathology 1992, 82, 652–656. [Google Scholar] [CrossRef]
- Biondi, E.; Blasioli, S.; Galeone, A.; Spinelli, F.; Cellini, A.; Lucchese, C.; Braschi, I. Detection of potato brown rot and ring rot by electronic nose: From laboratory to real scale. Talanta 2014, 129, 422–430. [Google Scholar] [CrossRef]
- El-Marzoky, H.A. Studies on Pythium leak rot disease on potato tubers during storage in Egypt. J. Plant Prot. Pathol. 2014, 5, 251–264. [Google Scholar] [CrossRef]
- Lui, L.; Vikram, A.; Abu-Nada, Y.; Kushalappa, A.; Raghavan, G.; Al-Mughrabi, K. Volatile metabolic profiling for discrimination of potato tubers inoculated with dry and soft rot pathogens. Am. J. Potato Res. 2005, 82, 1–8. [Google Scholar] [CrossRef]
- Barr, D.; Warwick, S.; Desaulniers, N. Isozyme variation, morphology, and growth response to temperature in Pythium ultimum. Can. J. Bot. 1996, 74, 753–761. [Google Scholar] [CrossRef]
- Kihurani, A.W. Factors Associated with Postharvest Deterioration of Sweetpotato (Ipomoea batatas L.) Roots in Kenya; University of Nairobi: Nairobi, Kenya, 2004. [Google Scholar]
- Olson, J.; Damicone, J.; Kahn, B. Identification and characterization of isolates of Pythium and Phytophthora spp. from snap beans with cottony leak. Plant Dis. 2016, 100, 1446–1453. [Google Scholar] [CrossRef]
- Almohammedy, M.A.B.; Al-maliky, B.S.A. Morphological, molecular and pathogenicity characterization of Pythium spp. on Zucchini isolated from soil and diseased plants in Iraq. Plant. Arch. 2020, 20, 767–775. [Google Scholar]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exp. | Cultivar | Inoculation | Replicates | Storage Conditions | Sampling Days (DAI) | |
|---|---|---|---|---|---|---|
| Temperature (°C) | Humidity (%) | |||||
| 1 | RR | P. ultimum | 5 | 25 | 30 | 0, 1, 3, 5, 7, 14 |
| RR | sterile PDA plug | 5 | 25 | 30 | ||
| 2 | RB | P. ultimum | 5 | 25 | 30 | |
| RB | sterile PDA plug | 5 | 25 | 30 | ||
| 3 | RR | P. ultimum | 5 | 4 | 95 | 0, 1, 5, 10, 15, 31 |
| RR | sterile PDA plug | 5 | 4 | 95 | ||
| 4 | RB | P. ultimum | 5 | 4 | 95 | |
| RB | sterile PDA plug | 5 | 4 | 95 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kothawade, G.S.; Sankaran, S.; Bates, A.A.; Schroeder, B.K.; Khot, L.R. Feasibility of Volatile Biomarker-Based Detection of Pythium Leak in Postharvest Stored Potato Tubers Using Field Asymmetric Ion Mobility Spectrometry. Sensors 2020, 20, 7350. https://doi.org/10.3390/s20247350
Kothawade GS, Sankaran S, Bates AA, Schroeder BK, Khot LR. Feasibility of Volatile Biomarker-Based Detection of Pythium Leak in Postharvest Stored Potato Tubers Using Field Asymmetric Ion Mobility Spectrometry. Sensors. 2020; 20(24):7350. https://doi.org/10.3390/s20247350
Chicago/Turabian StyleKothawade, Gajanan S., Sindhuja Sankaran, Austin A. Bates, Brenda K. Schroeder, and Lav R. Khot. 2020. "Feasibility of Volatile Biomarker-Based Detection of Pythium Leak in Postharvest Stored Potato Tubers Using Field Asymmetric Ion Mobility Spectrometry" Sensors 20, no. 24: 7350. https://doi.org/10.3390/s20247350