
Triptolide, a Cancer Cell Proliferation Inhibitor, Causes Zebrafish Muscle Defects by Regulating Notch and STAT3 Signaling Pathways

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
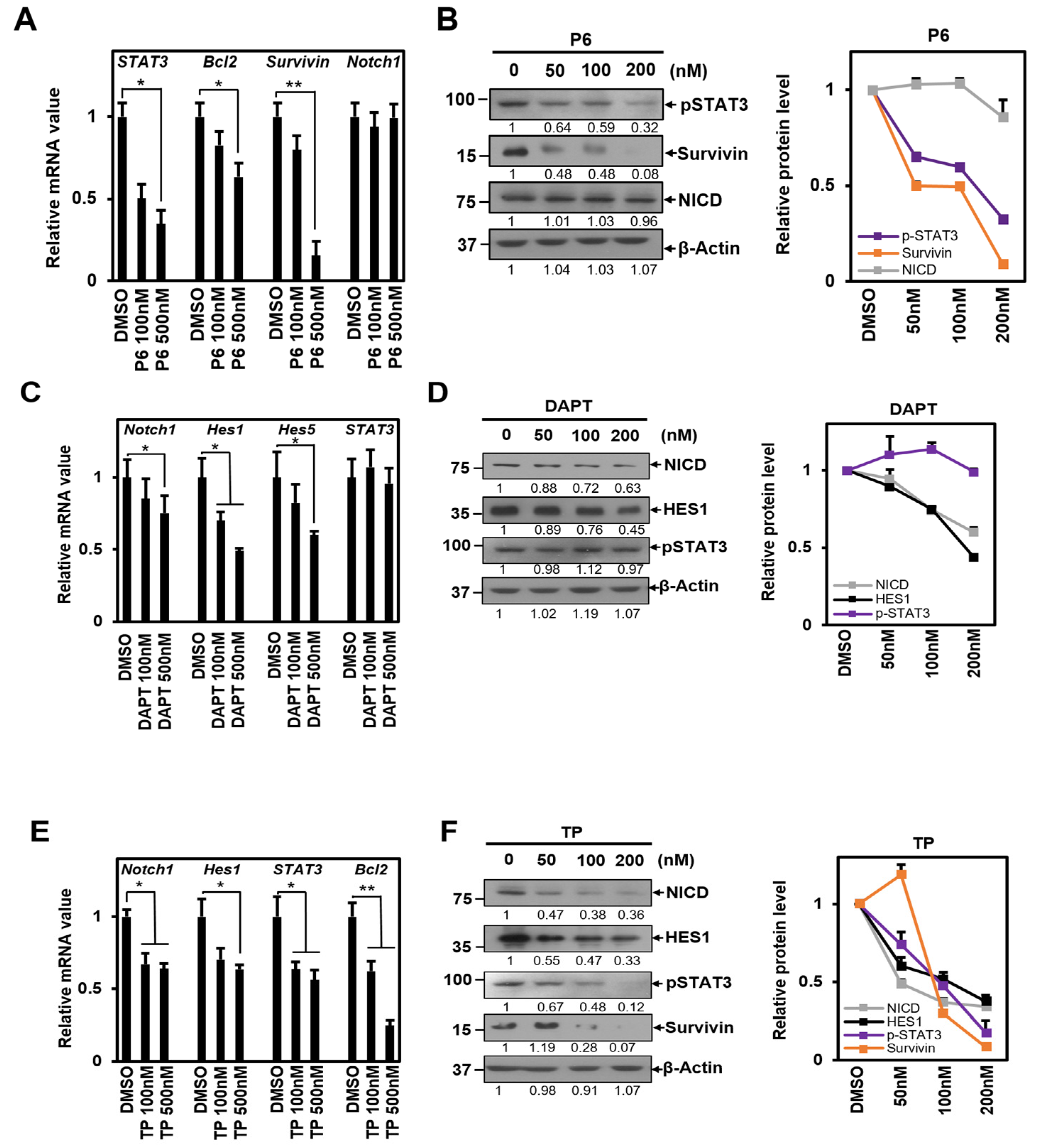
2.1. TP Inhibits Both STAT3 and Notch1 Signal
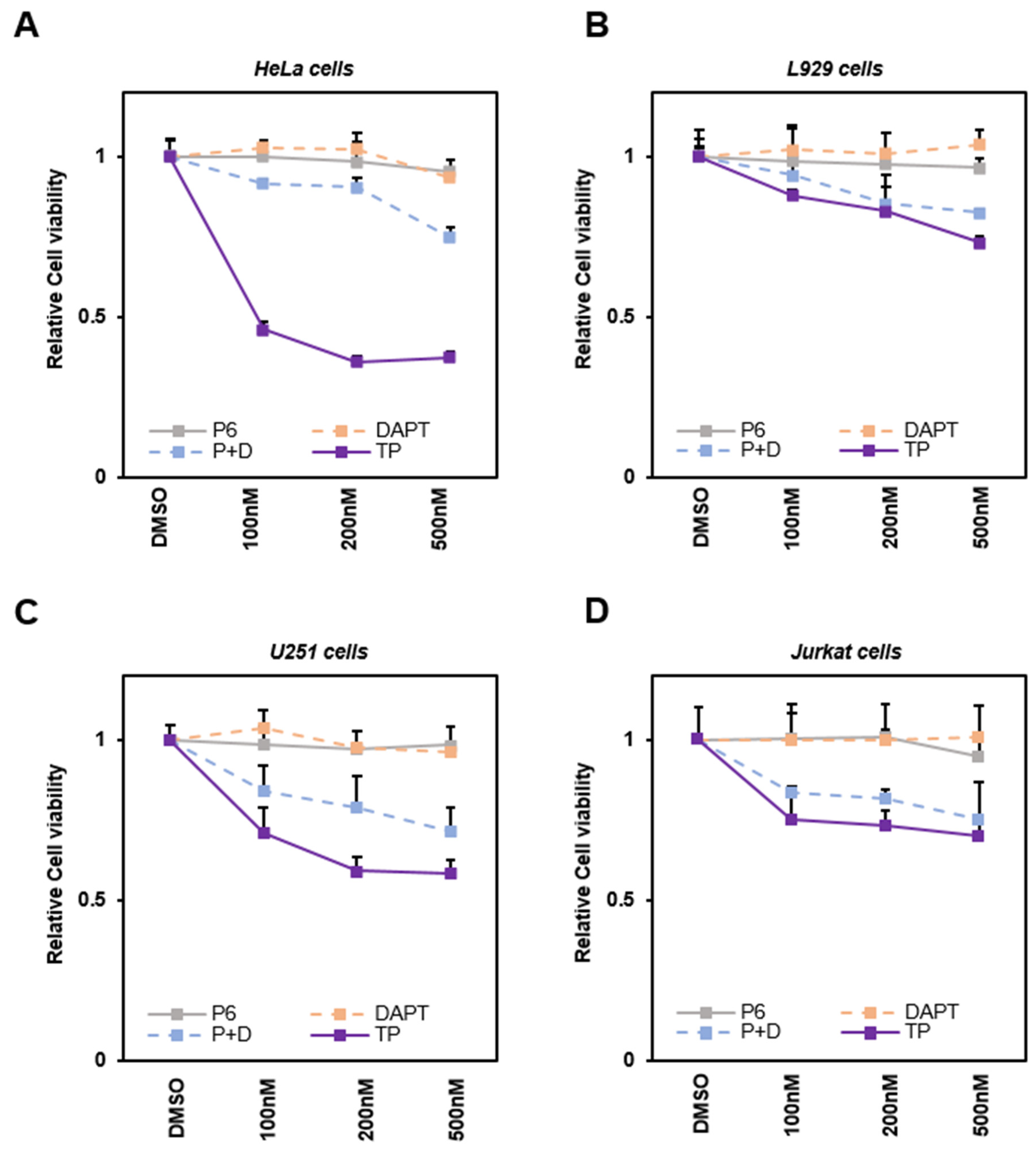
2.2. TP Reduces the Viability of Various Cancer Cells
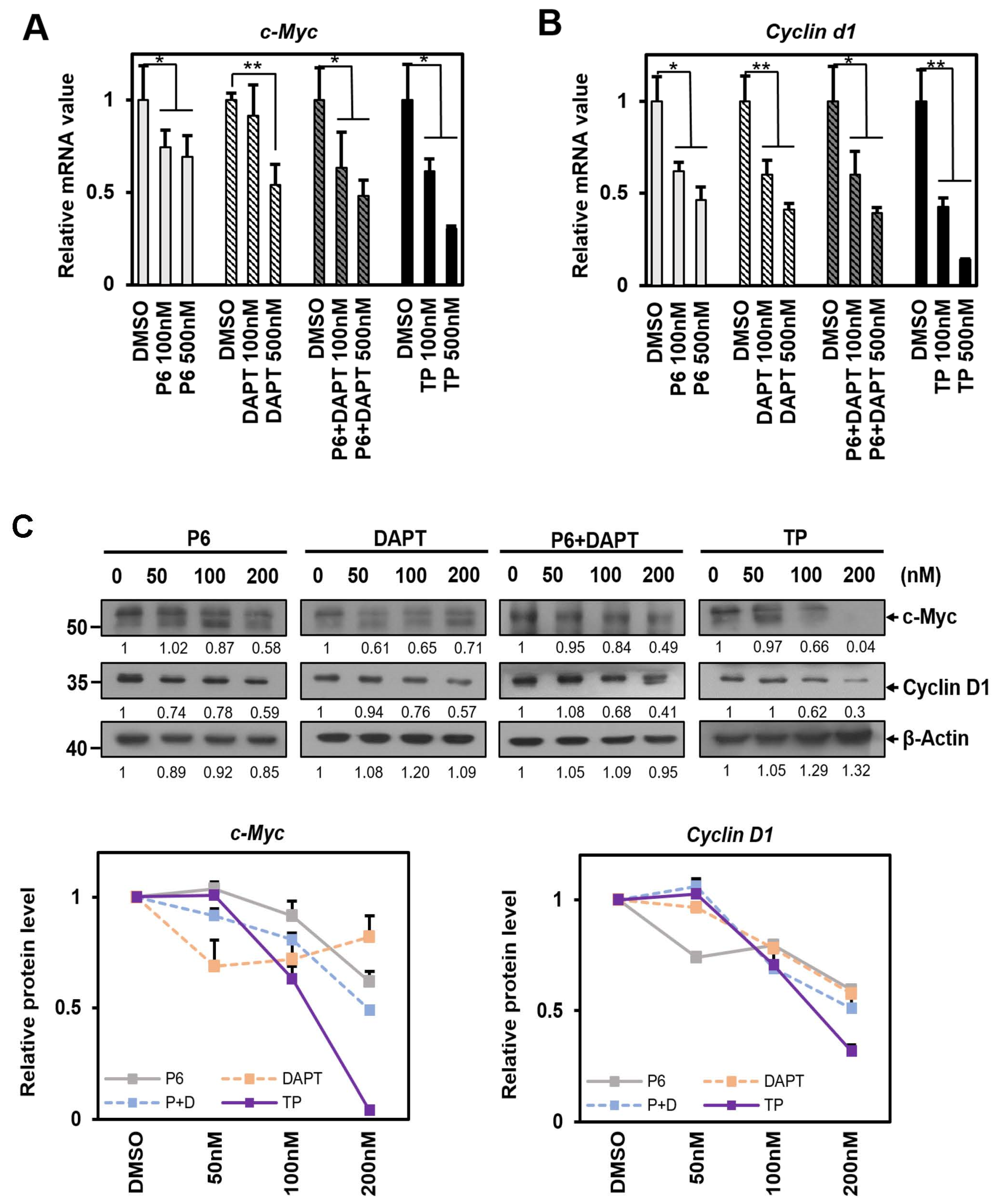
2.3. TP Reduces c-Myc and Cyclin d1 mRNA and These Proteins in Cell Proliferation
2.4. TP Reduces the Body Length of Zebrafish, Similar to the Combined Treatment with P6 and DAPT
2.5. TP Impairs Zebrafish Muscle Development
3. Discussion
4. Materials and Methods
4.1. Antibodies
4.2. Cell Maintenance
4.3. Zebrafish System
4.4. Chemical Treatment
4.5. Luciferase Reporter Assay
4.6. Quantitative RT-PCR (qRT-PCR)
4.7. Western Blotting
4.8. Cell Viability Assay
4.9. Measurement of Zebrafish Body Length
4.10. Immunohistochemistry
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TP | Triptolide |
| P6 | Pyridone 6, a pan Janus-activated kinase inhibitor |
| DAPT | γ-secretase inhibitor, (N-[N-(3,5-Difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester) |
| STAT3 | Signal transducer and activators of transcription3 |
| NICD | Notch Intracellular Domain |
| JAK | Janus-kinase |
References
- Wang, Q.; Meng, J.; Dong, A.; Yu, J.Z.; Zhang, G.X.; Ma, C.G. The Pharmacological Effects and Mechanism of Tripterygium wilfordii Hook F in Central Nervous System Autoimmunity. J. Altern. Complement. Med. 2016, 22, 496–502. [Google Scholar] [CrossRef]
- Kupchan, S.M.; Court, W.A.; Dailey, R.G., Jr.; Gilmore, C.J.; Bryan, R.F. Triptolide and tripdiolide, novel antileukemic diterpenoid triepoxides from Tripterygium wilfordii. J. Am. Chem. Soc. 1972, 94, 7194–7195. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Zhao, Y.; Zheng, Y. Therapeutic potential of triptolide in autoimmune diseases and strategies to reduce its toxicity. Chin. Med. 2021, 16, 114. [Google Scholar] [CrossRef]
- Ziaei, S.; Halaby, R. Immunosuppressive, anti-inflammatory and anti-cancer properties of triptolide: A mini review. Avicenna J. Phytomed. 2016, 6, 149. [Google Scholar] [PubMed]
- Tian, Y.; Li, P.; Xiao, Z.; Zhou, J.; Xue, X.; Jiang, N.; Peng, C.; Wu, L.; Tian, H.; Popper, H.; et al. Triptolide inhibits epithelial-mesenchymal transition phenotype through the p70S6k/GSK3/beta-catenin signaling pathway in taxol-resistant human lung adenocarcinoma. Transl. Lung Cancer Res. 2021, 10, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.D.; Lei, X.P.; Zhong, Y.H.; Chen, M.S.; Ke, Y.Y.; Li, Z.; Chen, J.; Huang, L.J.; Zhang, Y.; Liang, L.; et al. Triptolide suppresses the growth and metastasis of non-small cell lung cancer by inhibiting beta-catenin-mediated epithelial-mesenchymal transition. Acta Pharmacol. Sin. 2021, 42, 1486–1497. [Google Scholar] [CrossRef] [PubMed]
- Furtek, S.L.; Backos, D.S.; Matheson, C.J.; Reigan, P. Strategies and Approaches of Targeting STAT3 for Cancer Treatment. ACS Chem. Biol. 2016, 11, 308–318. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Jove, R. The STATs of cancer--new molecular targets come of age. Nat. Rev. Cancer 2004, 4, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Shaw, P.E. Autocrine-mediated activation of STAT3 correlates with cell proliferation in breast carcinoma lines. J. Biol. Chem. 2002, 277, 17397–17405. [Google Scholar] [CrossRef]
- Tsareva, S.A.; Moriggl, R.; Corvinus, F.M.; Wiederanders, B.; Schutz, A.; Kovacic, B.; Friedrich, K. Signal transducer and activator of transcription 3 activation promotes invasive growth of colon carcinomas through matrix metalloproteinase induction. Neoplasia 2007, 9, 279–291. [Google Scholar] [CrossRef]
- Zhong, Y.; Le, F.; Cheng, J.; Luo, C.; Zhang, X.; Wu, X.; Xu, F.; Zuo, Q.; Tan, B. Triptolide inhibits JAK2/STAT3 signaling and induces lethal autophagy through ROS generation in cisplatin-resistant SKOV3/DDP ovarian cancer cells. Oncol. Rep. 2021, 45, 69. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Park, B. Triptolide blocks the STAT3 signaling pathway through induction of protein tyrosine phosphatase SHP-1 in multiple myeloma cells. Int. J. Mol. Med. 2017, 40, 1566–1572. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Jin, H.; Xu, R.; Mei, Q.; Fan, D. Triptolide downregulates Rac1 and the JAK/STAT3 pathway and inhibits colitis-related colon cancer progression. Exp. Mol. Med. 2009, 41, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Gurzov, E.N.; Stanley, W.J.; Pappas, E.G.; Thomas, H.E.; Gough, D.J. The JAK/STAT pathway in obesity and diabetes. FEBS J. 2016, 283, 3002–3015. [Google Scholar] [CrossRef] [PubMed]
- Kanda, N.; Seno, H.; Konda, Y.; Marusawa, H.; Kanai, M.; Nakajima, T.; Kawashima, T.; Nanakin, A.; Sawabu, T.; Uenoyama, Y.; et al. STAT3 is constitutively activated and supports cell survival in association with survivin expression in gastric cancer cells. Oncogene 2004, 23, 4921–4929. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Zhang, Z.G.; Tian, X.Q.; Sun, D.F.; Liang, Q.C.; Zhang, Y.J.; Lu, R.; Chen, Y.X.; Fang, J.Y. Inhibition of JAK1, 2/STAT3 signaling induces apoptosis, cell cycle arrest, and reduces tumor cell invasion in colorectal cancer cells. Neoplasia 2008, 10, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Koerdel, K.; Spitzner, M.; Meyer, T.; Engels, N.; Krause, F.; Gaedcke, J.; Conradi, L.C.; Haubrock, M.; Beissbarth, T.; Leha, A.; et al. NOTCH Activation via gp130/STAT3 Signaling Confers Resistance to Chemoradiotherapy. Cancers 2021, 13, 455. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, D.; Uhle, F.; Sahin, D.; Krauser, U.; Weigand, M.A.; Heeg, K. The Interplay of Notch Signaling and STAT3 in TLR-Activated Human Primary Monocytes. Front. Cell Infect. Microbiol. 2018, 8, 241. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.C.; Suriano, G.; Mendes, N.; Gomes, B.; Wen, X.; Carneiro, F.; Seruca, R.; Machado, J.C. E-cadherin impairment increases cell survival through Notch-dependent upregulation of Bcl-2. Human Mol. Genet. 2012, 21, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Cohen, B.; Shimizu, M.; Izrailit, J.; Ng, N.F.; Buchman, Y.; Pan, J.G.; Dering, J.; Reedijk, M. Cyclin D1 is a direct target of JAG1-mediated Notch signaling in breast cancer. Breast Cancer Res. Treat. 2010, 123, 113–124. [Google Scholar] [CrossRef]
- Li, M.; Chen, F.; Clifton, N.; Sullivan, D.M.; Dalton, W.S.; Gabrilovich, D.I.; Nefedova, Y. Combined inhibition of Notch signaling and Bcl-2/Bcl-xL results in synergistic antimyeloma effect. Mol. Cancer Ther. 2010, 9, 3200–3209. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.; Jolicoeur, P. Notch-1 signaling promotes the cyclinD1-dependent generation of mammary tumor-initiating cells that can revert to bi-potential progenitors from which they arise. Oncogene 2013, 32, 3410–3419. [Google Scholar] [CrossRef]
- Aoki, Y.; Feldman, G.M.; Tosato, G. Inhibition of STAT3 signaling induces apoptosis and decreases survivin expression in primary effusion lymphoma. Blood J. Am. Soc. Hematol. 2003, 101, 1535–1542. [Google Scholar] [CrossRef]
- Ramana, C.V.; Grammatikakis, N.; Chernov, M.; Nguyen, H.; Goh, K.C.; Williams, B.R.; Stark, G.R. Regulation of c-myc expression by IFN-γ through Stat1-dependent and-independent pathways. EMBO J. 2000, 19, 263–272. [Google Scholar] [CrossRef]
- Amaya, M.L.; Inguva, A.; Pei, S.; Jones, C.; Krug, A.; Ye, H.; Minhajuddin, M.; Winters, A.; Furtek, S.L.; Gamboni, F.; et al. The STAT3-MYC axis promotes survival of leukemia stem cells by regulating SLC1A5 and oxidative phosphorylation. Blood 2022, 139, 584–596. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Han, Z.C. STAT3: A critical transcription activator in angiogenesis. Med. Res. Rev. 2008, 28, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Limbourg, F.P.; Takeshita, K.; Radtke, F.; Bronson, R.T.; Chin, M.T.; Liao, J.K. Essential role of endothelial Notch1 in angiogenesis. Circulation 2005, 111, 1826–1832. [Google Scholar] [CrossRef]
- Teraoka, H.; Dong, W.; Hiraga, T. Zebrafish as a novel experimental model for developmental toxicology. Congenit. Anom. 2003, 43, 123–132. [Google Scholar] [CrossRef]
- Lee, H.C.; Lin, C.Y.; Tsai, H.J. Zebrafish, an In Vivo Platform to Screen Drugs and Proteins for Biomedical Use. Pharmaceuticals 2021, 14, 500. [Google Scholar] [CrossRef]
- Dumont, N.A.; Bentzinger, C.F.; Sincennes, M.C.; Rudnicki, M.A. Satellite Cells and Skeletal Muscle Regeneration. Compr. Physiol. 2015, 5, 1027–1059. [Google Scholar]
- Mallard, J.; Hucteau, E.; Hureau, T.J.; Pagano, A.F. Skeletal Muscle Deconditioning in Breast Cancer Patients Undergoing Chemotherapy: Current Knowledge and Insights from Other Cancers. Front. Cell Dev. Biol. 2021, 9, 719643. [Google Scholar] [CrossRef] [PubMed]
- Sandri, M. Protein breakdown in muscle wasting: Role of autophagy-lysosome and ubiquitin-proteasome. Int. J. Biochem. Cell Biol. 2013, 45, 2121–2129. [Google Scholar] [CrossRef] [PubMed]
- Sartori, R.; Romanello, V.; Sandri, M. Mechanisms of muscle atrophy and hypertrophy: Implications in health and disease. Nat. Commun. 2021, 12, 330. [Google Scholar] [CrossRef]
- Rodrigues, C.; Joy, L.R.; Sachithanandan, S.P.; Krishna, S. Notch signalling in cervical cancer. Exp. Cell Res. 2019, 385, 111682. [Google Scholar] [CrossRef] [PubMed]
- Maliekal, T.T.; Bajaj, J.; Giri, V.; Subramanyam, D.; Krishna, S. The role of Notch signaling in human cervical cancer: Implications for solid tumors. Oncogene 2008, 27, 5110–5114. [Google Scholar] [CrossRef] [PubMed]
- Yashiro-Ohtani, Y.; He, Y.; Ohtani, T.; Jones, M.E.; Shestova, O.; Xu, L.; Fang, T.C.; Chiang, M.Y.; Intlekofer, A.M.; Blacklow, S.C.; et al. Pre-TCR signaling inactivates Notch1 transcription by antagonizing E2A. Genes. Dev. 2009, 23, 1665–1676. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liao, X.; Agarwal, M.K.; Barnes, L.; Auron, P.E.; Stark, G.R. Unphosphorylated STAT3 accumulates in response to IL-6 and activates transcription by binding to NFkappaB. Genes. Dev. 2007, 21, 1396–1408. [Google Scholar] [CrossRef]
- Masters, J.R. HeLa cells 50 years on: The good, the bad and the ugly. Nat. Rev. Cancer 2002, 2, 315–319. [Google Scholar] [CrossRef]
- Yang, F.; Bai, X.J.; Hu, D.; Li, Z.F.; Liu, K.J. Effect of triptolide on secretion of inflammatory cellular factors TNF-alpha and IL-8 in peritoneal macrophages of mice activated by lipopolysaccharide. World J. Emerg. Med. 2010, 1, 70–74. [Google Scholar]
- Meng, H.T.; Zhu, L.; Ni, W.M.; You, L.S.; Jin, J.; Qian, W.B. Triptolide inhibits the proliferation of cells from lymphocytic leukemic cell lines in association with downregulation of NF-kappaB activity and miR-16-1*. Acta Pharmacol. Sin. 2011, 32, 503–511. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, P.; Wang, X.; Wang, L.; Zhu, Y.; Gao, W. Triptolide Induces Glioma Cell Autophagy and Apoptosis via Upregulating the ROS/JNK and Downregulating the Akt/mTOR Signaling Pathways. Front. Oncol. 2019, 9, 387. [Google Scholar] [CrossRef]
- Brinker, A.M.; Ma, J.; Lipsky, P.E.; Raskin, I. Medicinal chemistry and pharmacology of genus Tripterygium (Celastraceae). Phytochemistry 2007, 68, 732–766. [Google Scholar] [CrossRef] [PubMed]
- Daya, A.; Donaka, R.; Karasik, D. Zebrafish models of sarcopenia. Dis. Model. Mech. 2020, 13, dmm042689. [Google Scholar] [CrossRef]
- Nicolas, H.A.; Hua, K.; Quigley, H.; Ivare, J.; Tesson, F.; Akimenko, M.A. A CRISPR/Cas9 zebrafish lamin A/C mutant model of muscular laminopathy. Dev. Dyn. 2022, 251, 645–661. [Google Scholar] [CrossRef]
- Kwon, S.; Ki, S.M.; Park, S.E.; Kim, M.-J.; Hyung, B.; Lee, N.K.; Shim, S.; Choi, B.-O.; Na, D.L.; Lee, J.E. Anti-apoptotic effects of human Wharton’s jelly-derived mesenchymal stem cells on skeletal muscle cells mediated via secretion of XCL1. Mol. Ther. 2016, 24, 1550–1560. [Google Scholar] [CrossRef]
- Gallagher, T.L.; Arribere, J.A.; Geurts, P.A.; Exner, C.R.; McDonald, K.L.; Dill, K.K.; Marr, H.L.; Adkar, S.S.; Garnett, A.T.; Amacher, S.L. Rbfox-regulated alternative splicing is critical for zebrafish cardiac and skeletal muscle functions. Dev. Biol. 2011, 359, 251–261. [Google Scholar] [CrossRef]
- Wen, Y.; Bi, P.; Liu, W.; Asakura, A.; Keller, C.; Kuang, S. Constitutive Notch activation upregulates Pax7 and promotes the self-renewal of skeletal muscle satellite cells. Mol. Cell Biol. 2012, 32, 2300–2311. [Google Scholar] [CrossRef] [PubMed]
- Tajbakhsh, S.; Rocancourt, D.; Cossu, G.; Buckingham, M. Redefining the genetic hierarchies controlling skeletal myogenesis: Pax-3 and Myf-5 act upstream of MyoD. Cell 1997, 89, 127–138. [Google Scholar] [CrossRef]
- Relaix, F.; Montarras, D.; Zaffran, S.; Gayraud-Morel, B.; Rocancourt, D.; Tajbakhsh, S.; Mansouri, A.; Cumano, A.; Buckingham, M. Pax3 and Pax7 have distinct and overlapping functions in adult muscle progenitor cells. J. Cell Biol. 2006, 172, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Lee, T.H.; Shim, J. Emerin suppresses Notch signaling by restricting the Notch intracellular domain to the nuclear membrane. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 303–313. [Google Scholar] [CrossRef]
- Andersson, E.R.; Sandberg, R.; Lendahl, U. Notch signaling: Simplicity in design, versatility in function. Development 2011, 138, 3593–3612. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, Q.; Shang, T.; Song, W.; Xu, D.; Allen, T.D.; Wang, X.; Jeong, J.; Lobe, C.G.; Liu, J. Aberrant Activation of Notch1 Signaling in Glomerular Endothelium Induces Albuminuria. Circ. Res. 2021, 128, 602–618. [Google Scholar] [CrossRef] [PubMed]
- Horita, N.; Keeley, T.M.; Hibdon, E.S.; Delgado, E.; Lafkas, D.; Siebel, C.W.; Samuelson, L.C. Delta-like 1-Expressing Cells at the Gland Base Promote Proliferation of Gastric Antral Stem Cells in Mouse. Cell Mol. Gastroenterol. Hepatol. 2022, 13, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Capaccione, K.M.; Pine, S.R. The Notch signaling pathway as a mediator of tumor survival. Carcinogenesis 2013, 34, 1420–1430. [Google Scholar] [CrossRef] [PubMed]
- Owen, D.H.; Giffin, M.J.; Bailis, J.M.; Smit, M.D.; Carbone, D.P.; He, K. DLL3: An emerging target in small cell lung cancer. J. Hematol. Oncol. 2019, 12, 61. [Google Scholar] [CrossRef] [PubMed]
- Pitulescu, M.E.; Schmidt, I.; Giaimo, B.D.; Antoine, T.; Berkenfeld, F.; Ferrante, F.; Park, H.; Ehling, M.; Biljes, D.; Rocha, S.F.; et al. Dll4 and Notch signalling couples sprouting angiogenesis and artery formation. Nat. Cell Biol. 2017, 19, 915–927. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.; Xiong, Z.; Kong, J.; Wang, L.; Cheng, Y.; Jin, J.; Huang, Z. Triptolide impairs thioredoxin system by suppressing Notch1-mediated PTEN/Akt/Txnip signaling in hepatocytes. Toxicol. Lett. 2019, 300, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Wang, S.; Chang, Y.; Li, C.; Yang, J.; Han, Z.; Chang, B.; Sun, B.; Chen, L. Triptolide prevents extracellular matrix accumulation in experimental diabetic kidney disease by targeting microRNA-137/Notch1 pathway. J. Cell Physiol. 2018, 233, 2225–2237. [Google Scholar] [CrossRef] [PubMed]
- Weber, S.; Niessen, M.T.; Prox, J.; Lullmann-Rauch, R.; Schmitz, A.; Schwanbeck, R.; Blobel, C.P.; Jorissen, E.; de Strooper, B.; Niessen, C.M.; et al. The disintegrin/metalloproteinase Adam10 is essential for epidermal integrity and Notch-mediated signaling. Development 2011, 138, 495–505. [Google Scholar] [CrossRef]
- Soundararajan, R.; Sayat, R.; Robertson, G.S.; Marignani, P.A. Triptolide: An inhibitor of a disintegrin and metalloproteinase 10 (ADAM10) in cancer cells. Cancer Biol. Ther. 2009, 8, 2054–2062. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, Z.; Wang, Y.; Ba, X.; Huang, Y.; Shen, P.; Wang, H.; Tu, S. Triptolide exerts an anti-tumor effect on non-small cell lung cancer cells by inhibiting activation of the IL-6/STAT3 axis. Int. J. Mol. Med. 2019, 44, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.Y.; Tseng, Y.T.; Lee, T.Y.; Fu, Y.C.; Chang, W.H.; Lo, W.W.; Lin, C.L.; Lo, Y.C. Triptolide prevents LPS-induced skeletal muscle atrophy via inhibiting NF-kappaB/TNF-alpha and regulating protein synthesis/degradation pathway. Br. J. Pharmacol. 2021, 178, 2998–3016. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Li, Z.-H.; Liu, T.; Jiang, C.-H.; Zhang, Y.-J.; Li, H.; Jiang, Y.; Zhao, J.; Guo, W.-J.; Guo, J.-Y. Progress in research on antitumor drugs and dynamic changes in skeletal muscles. Front. Pharmacol. 2022, 13, 893333. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.J.; Chiu, G.N. Role of oxidative stress, endoplasmic reticulum stress and ERK activation in triptolide-induced apoptosis. Int. J. Oncol. 2013, 42, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.T.; Kang, J.J.; Lee, K.Y.; Wei, K.; Anderson, E.; Gotmare, S.; Ross, J.A.; Rosen, G.D. Triptolide and chemotherapy cooperate in tumor cell apoptosis. A role for the p53 pathway. J. Biol. Chem. 2001, 276, 2221–2227. [Google Scholar] [CrossRef] [PubMed]
- Polge, C.; Heng, A.E.; Jarzaguet, M.; Ventadour, S.; Claustre, A.; Combaret, L.; Bechet, D.; Matondo, M.; Uttenweiler-Joseph, S.; Monsarrat, B.; et al. Muscle actin is polyubiquitinylated in vitro and in vivo and targeted for breakdown by the E3 ligase MuRF1. FASEB J. 2011, 25, 3790–3802. [Google Scholar] [CrossRef] [PubMed]
- Tajbakhsh, S. Skeletal muscle stem cells in developmental versus regenerative myogenesis. J. Intern. Med. 2009, 266, 372–389. [Google Scholar] [CrossRef] [PubMed]
- Brack, A.S.; Conboy, I.M.; Conboy, M.J.; Shen, J.; Rando, T.A. A temporal switch from notch to Wnt signaling in muscle stem cells is necessary for normal adult myogenesis. Cell Stem Cell 2008, 2, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Tierney, M.T.; Aydogdu, T.; Sala, D.; Malecova, B.; Gatto, S.; Puri, P.L.; Latella, L.; Sacco, A. STAT3 signaling controls satellite cell expansion and skeletal muscle repair. Nat. Med. 2014, 20, 1182–1186. [Google Scholar] [CrossRef]
- Price, F.D.; Von Maltzahn, J.; Bentzinger, C.F.; Dumont, N.A.; Yin, H.; Chang, N.C.; Wilson, D.H.; Frenette, J.; Rudnicki, M.A. Inhibition of JAK-STAT signaling stimulates adult satellite cell function. Nat. Med. 2014, 20, 1174–1181. [Google Scholar] [CrossRef]
- Chen, J.A.; Splenser, A.; Guillory, B.; Luo, J.; Mendiratta, M.; Belinova, B.; Halder, T.; Zhang, G.; Li, Y.P.; Garcia, J.M. Ghrelin prevents tumour- and cisplatin-induced muscle wasting: Characterization of multiple mechanisms involved. J. Cachexia Sarcopenia Muscle 2015, 6, 132–143. [Google Scholar] [CrossRef]
- Huo, J.; Yu, Q.; Zhang, Y.; Liu, K.; Hsiao, C.D.; Jiang, Z.; Zhang, L. Triptolide-induced hepatotoxicity via apoptosis and autophagy in zebrafish. J. Appl. Toxicol. 2019, 39, 1532–1540. [Google Scholar] [CrossRef]
- Jeong, H.S.; Kim, H.J.; Kim, D.H.; Chung, K.W.; Choi, B.O.; Lee, J.E. Therapeutic Potential of CKD-504, a Novel Selective Histone Deacetylase 6 Inhibitor, in a Zebrafish Model of Neuromuscular Junction Disorders. Mol. Cells 2022, 45, 231–242. [Google Scholar] [CrossRef]
- Chahardehi, A.M.; Arsad, H.; Lim, V. Zebrafish as a Successful Animal Model for Screening Toxicity of Medicinal Plants. Plants 2020, 9, 1345. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.P.; Feng, F.; Zhang, X.Q.; Liu, X.X.; Wang, Y.B.; She, J.X.; He, Z.H.; He, M.F. Toxicity assessment of 7 anticancer compounds in zebrafish. Int. J. Toxicol. 2014, 33, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, P.; Dandawate, P.; Jensen, R.A.; Anant, S. Celastrol and Triptolide Suppress Stemness in Triple Negative Breast Cancer: Notch as a Therapeutic Target for Stem Cells. Biomedicines 2021, 9, 482. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, B.; Park, Y.; Lee, Y.; Kwon, S.; Shim, J. Triptolide, a Cancer Cell Proliferation Inhibitor, Causes Zebrafish Muscle Defects by Regulating Notch and STAT3 Signaling Pathways. Int. J. Mol. Sci. 2024, 25, 4675. https://doi.org/10.3390/ijms25094675
Lee B, Park Y, Lee Y, Kwon S, Shim J. Triptolide, a Cancer Cell Proliferation Inhibitor, Causes Zebrafish Muscle Defects by Regulating Notch and STAT3 Signaling Pathways. International Journal of Molecular Sciences. 2024; 25(9):4675. https://doi.org/10.3390/ijms25094675
Chicago/Turabian StyleLee, Byongsun, Yongjin Park, Younggwang Lee, Seyoung Kwon, and Jaekyung Shim. 2024. "Triptolide, a Cancer Cell Proliferation Inhibitor, Causes Zebrafish Muscle Defects by Regulating Notch and STAT3 Signaling Pathways" International Journal of Molecular Sciences 25, no. 9: 4675. https://doi.org/10.3390/ijms25094675