
Generation of Slco1a4-CreERT2-tdTomato Knock-in Mice for Specific Cerebrovascular Endothelial Cell Targeting


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Slco1a4 Is a Candidate Marker of Cerebrovascular ECs
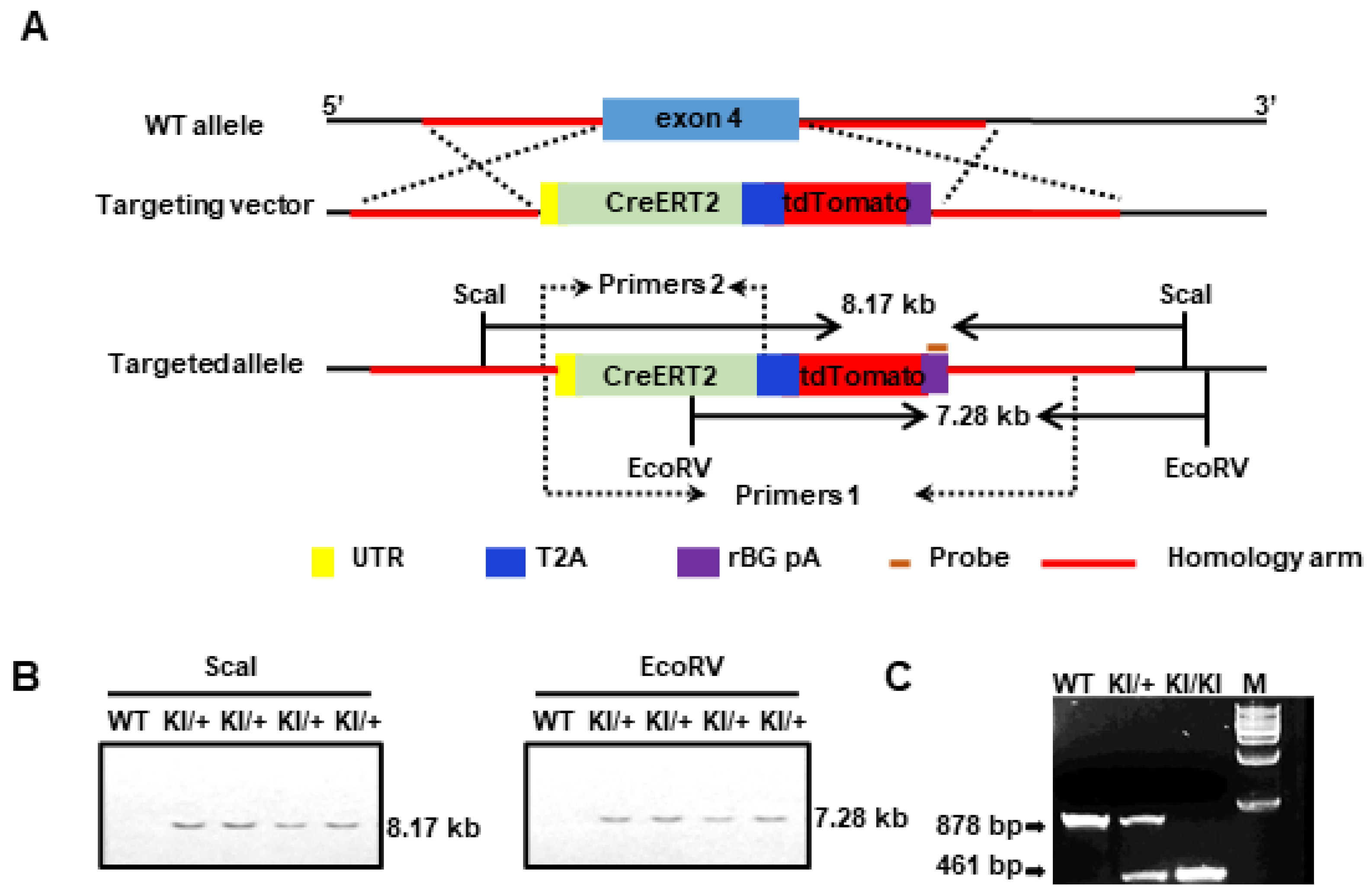
2.2. Generation of the Slco1a4-CreERT2-tdTomato Knock-in Mouse Line
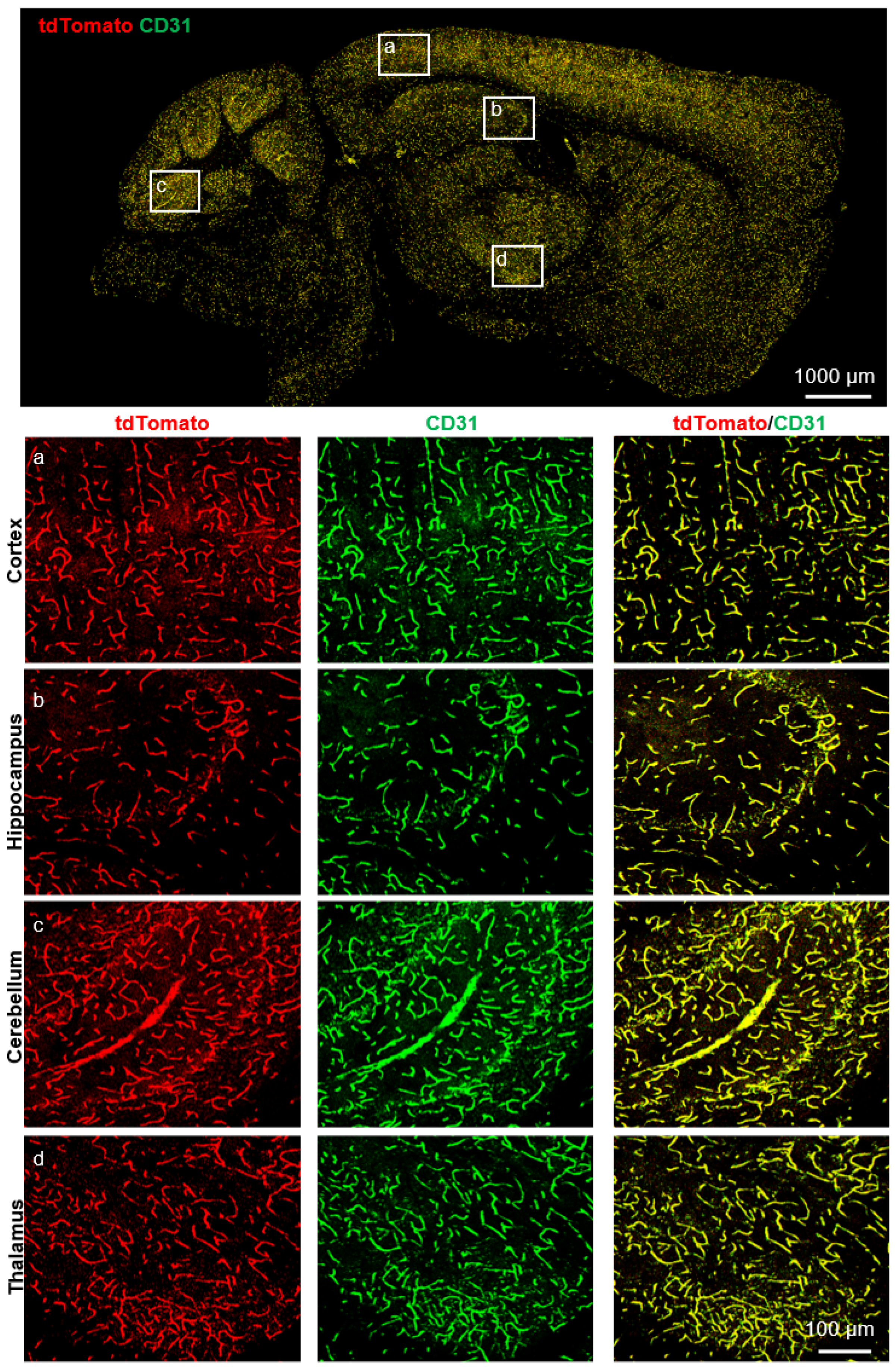
2.3. Cerebrovascular ECs-Specific Expression of Slco1a4
2.4. Cerebrovascular Endothelial Cells-Specific Cre Activity of Slco1a4-CreERT2 Mice
3. Discussion
4. Materials and Methods
4.1. Generation of Slco1a4-CreERT2-T2A-tdTomato
4.2. Mouse Strains
4.3. Experimental Design
4.4. Tamoxifen Administration
4.5. Genotyping by PCR
4.6. Immunostaining
4.7. Southern Blot
4.8. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Zierfuss, B.; Larochelle, C.; Prat, A. Blood–brain barrier dysfunction in multiple sclerosis: Causes, consequences, and potential effects of therapies. Lancet Neurol. 2024, 23, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Shue, F.; Bu, G.; Kanekiyo, T. Pathophysiology and probable etiology of cerebral small vessel disease in vascular dementia and Alzheimer’s disease. Mol. Neurodegener. 2023, 18, 46. [Google Scholar] [CrossRef] [PubMed]
- Ben-Zvi, A.; Liebner, S. Developmental regulation of barrier- and non-barrier blood vessels in the CNS. J. Intern. Med. 2021, 292, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Andreone, B.J.; Chow, B.W.; Tata, A.; Lacoste, B.; Ben-Zvi, A.; Bullock, K.; Deik, A.A.; Ginty, D.D.; Clish, C.B.; Gu, C. Blood-Brain Barrier Permeability Is Regulated by Lipid Transport-Dependent Suppression of Caveolae-Mediated Transcytosis. Neuron 2017, 94, 581–594.e5. [Google Scholar] [CrossRef] [PubMed]
- Engert, S.; Liao, W.P.; Burtscher, I.; Lickert, H. Sox17-2A-iCre: A knock-in mouse line expressing Cre recombinase in endoderm and vascular endothelial cells. Genesis 2009, 47, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Kisanuki, Y.Y.; Hammer, R.E.; Miyazaki, J.-i.; Williams, S.C.; Richardson, J.A.; Yanagisawa, M. Tie2-Cre Transgenic Mice: A New Model for Endothelial Cell-Lineage Analysis In Vivo. Dev. Biol. 2001, 230, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Monvoisin, A.; Alva, J.A.; Hofmann, J.J.; Zovein, A.C.; Lane, T.F.; Iruela-Arispe, M.L. VE-cadherin-CreERT2 transgenic mouse: A model for inducible recombination in the endothelium. Dev. Dyn. 2006, 235, 3413–3422. [Google Scholar] [CrossRef]
- Meng, F.; Shi, L.; Cheng, X.; Hou, N.; Wang, Y.; Teng, Y.; Meng, A.; Yang, X. Surfactant protein A promoter directs the expression of Cre recombinase in brain microvascular endothelial cells of transgenic mice. Matrix Biol. 2007, 26, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Lan, Y.; Wang, Y.; Wang, J.; Yang, G.; Meng, F.; Han, H.; Meng, A.; Wang, Y.; Yang, X. Endothelial Smad4 maintains cerebrovascular integrity by activating N-cadherin through cooperation with Notch. Dev. Cell 2011, 20, 291–302. [Google Scholar] [CrossRef]
- Vanlandewijck, M.; He, L.; Mäe, M.A.; Andrae, J.; Ando, K.; Del Gaudio, F.; Nahar, K.; Lebouvier, T.; Laviña, B.; Gouveia, L.; et al. A molecular atlas of cell types and zonation in the brain vasculature. Nature 2018, 554, 475–480. [Google Scholar] [CrossRef]
- Schaum, N.; Karkanias, J.; Neff, N.F.; May, A.P.; Quake, S.R.; Wyss-Coray, T.; Darmanis, S.; Batson, J.; Botvinnik, O.; Chen, M.B.; et al. Single-cell transcriptomics of 20 mouse organs creates a Tabula Muris. Nature 2018, 562, 367–372. [Google Scholar] [CrossRef]
- Nies, A.T.; Schaeffeler, E.; Schwab, M. Hepatic solute carrier transporters and drug therapy: Regulation of expression and impact of genetic variation. Pharmacol. Ther. 2022, 238, 108268. [Google Scholar] [CrossRef]
- Takano, J.; Maeda, K.; Kusuhara, H.; Sugiyama, Y. Organic Anion Transporting Polypeptide 1a4 is Responsible for the Hepatic Uptake of Cardiac Glycosides in Mice. Drug Metab. Dispos. 2018, 46, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Csanaky, I.L.; Selwyn, F.P.; Lehman-McKeeman, L.D.; Klaassen, C.D. Organic anion-transporting polypeptide 1a4 (Oatp1a4) is important for secondary bile acid metabolism. Biochem. Pharmacol. 2013, 86, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuki, S.; Ito, S.; Matsuda, A.; Hori, S.; Abe, T.; Terasaki, T. Brain-to-blood elimination of 24S-hydroxycholesterol from rat brain is mediated by organic anion transporting polypeptide 2 (oatp2) at the blood-brain barrier. J. Neurochem. 2007, 103, 1430–1438. [Google Scholar] [CrossRef]
- Miyajima, M.; Kusuhara, H.; Fujishima, M.; Adachi, Y.; Sugiyama, Y. Organic anion transporter 3 mediates the efflux transport of an amphipathic organic anion, dehydroepiandrosterone sulfate, across the blood-brain barrier in mice. Drug Metab. Dispos. 2011, 39, 814–819. [Google Scholar] [CrossRef] [PubMed]
- Kühn, R.; Schwenk, F.; Aguet, M.; Rajewsky, K. Inducible gene targeting in mice. Science 1995, 269, 1427–1429. [Google Scholar] [CrossRef]
- Zhang, Y.; Riesterer, C.; Ayrall, A.M.; Sablitzky, F.; Littlewood, T.D.; Reth, M. Inducible site-directed recombination in mouse embryonic stem cells. Nucleic Acids Res. 1996, 24, 543–548. [Google Scholar] [CrossRef]
- Indra, A.K.; Warot, X.; Brocard, J.; Bornert, J.M.; Xiao, J.H.; Chambon, P.; Metzger, D. Temporally-controlled site-specific mutagenesis in the basal layer of the epidermis: Comparison of the recombinase activity of the tamoxifen-inducible Cre-ER(T) and Cre-ER(T2) recombinases. Nucleic Acids Res. 1999, 27, 4324–4327. [Google Scholar] [CrossRef]
- Hamada, K.; Sasaki, T.; Koni, P.A.; Natsui, M.; Kishimoto, H.; Sasaki, J.; Yajima, N.; Horie, Y.; Hasegawa, G.; Naito, M.; et al. The PTEN/PI3K pathway governs normal vascular development and tumor angiogenesis. Genes Dev. 2005, 19, 2054–2065. [Google Scholar] [CrossRef]
- Wang, J.; Cui, Y.; Yu, Z.; Wang, W.; Cheng, X.; Ji, W.; Guo, S.; Zhou, Q.; Wu, N.; Chen, Y.; et al. Brain Endothelial Cells Maintain Lactate Homeostasis and Control Adult Hippocampal Neurogenesis. Cell Stem Cell 2019, 25, 754–767.e759. [Google Scholar] [CrossRef] [PubMed]
- Lang, M.F.; Salinin, S.; Ridder, D.A.; Kleesiek, J.; Hroudova, J.; Berger, S.; Schütz, G.; Schwaninger, M. A transgenic approach to identify thyroxine transporter-expressing structures in brain development. J. Neuroendocrinol. 2011, 23, 1194–1203. [Google Scholar] [CrossRef] [PubMed]
- Ren, A.A.; Snellings, D.A.; Su, Y.S.; Hong, C.C.; Castro, M.; Tang, A.T.; Detter, M.R.; Hobson, N.; Girard, R.; Romanos, S.; et al. PIK3CA and CCM mutations fuel cavernomas through a cancer-like mechanism. Nature 2021, 594, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Detter, M.R.; Shenkar, R.; Benavides, C.R.; Neilson, C.A.; Moore, T.; Lightle, R.; Hobson, N.; Shen, L.; Cao, Y.; Girard, R.; et al. Novel Murine Models of Cerebral Cavernous Malformations. Angiogenesis 2020, 23, 651–666. [Google Scholar] [CrossRef] [PubMed]
- Ridder, D.A.; Lang, M.F.; Salinin, S.; Röderer, J.P.; Struss, M.; Maser-Gluth, C.; Schwaninger, M. TAK1 in brain endothelial cells mediates fever and lethargy. J. Exp. Med. 2011, 208, 2615–2623. [Google Scholar] [CrossRef] [PubMed]
- Pu, W.; Zhang, H.; Huang, X.; Tian, X.; He, L.; Wang, Y.; Zhang, L.; Liu, Q.; Li, Y.; Li, Y.; et al. Mfsd2a+ hepatocytes repopulate the liver during injury and regeneration. Nat. Commun. 2016, 7, 13369. [Google Scholar] [CrossRef] [PubMed]
- Pu, W.; He, L.; Han, X.; Tian, X.; Li, Y.; Zhang, H.; Liu, Q.; Huang, X.; Zhang, L.; Wang, Q.D.; et al. Genetic Targeting of Organ-Specific Blood Vessels. Circ. Res. 2018, 123, 86–99. [Google Scholar] [CrossRef]
- Ose, A.; Kusuhara, H.; Endo, C.; Tohyama, K.; Miyajima, M.; Kitamura, S.; Sugiyama, Y. Functional characterization of mouse organic anion transporting peptide 1a4 in the uptake and efflux of drugs across the blood-brain barrier. Drug Metab. Dispos. 2010, 38, 168–176. [Google Scholar] [CrossRef]
- Soriano, P. Generalized lacZ expression with the ROSA26 Cre reporter strain. Nat. Genet. 1999, 21, 70–71. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, C.; Li, S.; Cai, Y.; Lu, J.; Teng, Y.; Yang, X.; Wang, J. Generation of Slco1a4-CreERT2-tdTomato Knock-in Mice for Specific Cerebrovascular Endothelial Cell Targeting. Int. J. Mol. Sci. 2024, 25, 4666. https://doi.org/10.3390/ijms25094666
Xu C, Li S, Cai Y, Lu J, Teng Y, Yang X, Wang J. Generation of Slco1a4-CreERT2-tdTomato Knock-in Mice for Specific Cerebrovascular Endothelial Cell Targeting. International Journal of Molecular Sciences. 2024; 25(9):4666. https://doi.org/10.3390/ijms25094666
Chicago/Turabian StyleXu, Chengfang, Shounian Li, Yunting Cai, Jinjin Lu, Yan Teng, Xiao Yang, and Jun Wang. 2024. "Generation of Slco1a4-CreERT2-tdTomato Knock-in Mice for Specific Cerebrovascular Endothelial Cell Targeting" International Journal of Molecular Sciences 25, no. 9: 4666. https://doi.org/10.3390/ijms25094666