
Have All of the Phytohormonal Properties of Melatonin Been Verified?

Department of Molecular Biology, Dankook University, Cheonan-si 31116, Republic of Korea
Int. J. Mol. Sci. 2024, 25(6), 3550; https://doi.org/10.3390/ijms25063550
Submission received: 17 February 2024
/
Revised: 15 March 2024
/
Accepted: 19 March 2024
/
Published: 21 March 2024
(This article belongs to the Special Issue The Role of Melatonin in Plants 2.0)
Abstract
:Melatonin is a ubiquitous regulator in plants and performs a variety of physiological roles, including resistance to abiotic stress, regulation of growth and development, and enhancement of plant immunity. Melatonin exhibits the characteristics of a phytohormone with its pleiotropic effects, biosynthesis, conjugation, catabolism, effective concentration, and the shape and location of its dose–response curves. In addition, CAND2/PMTR1, a phytomelatonin receptor candidate belonging to the G protein-coupled receptors (GPCRs), supports the concept of melatonin as a phytohormone. However, the biochemistry of plant melatonin receptors needs to be further characterized. In particular, some of the experimental findings to date cannot be explained by known GPCR signaling mechanisms, so further studies are needed to explore the possibility of novel signaling mechanisms.
1. Introduction
Since melatonin (N-acetyl-5-methoxytrypamine) was found in plants in 1995 [1,2,3], increasing efforts have been focused on phytomelatonin research. I found more than 860 papers in the most recent 5 years through a web search with the combined key words “melatonin” and “plant” (https://pubmed.ncbi.nlm.nih.gov (accessed on 24 January 2024)). Interestingly, about 200 papers among them are reviews. This high number of review papers may indicate the great interest of the community but also not enough research verifying the action mechanism.
Melatonin is originally known as a neurohormone found in the bovine pineal gland that regulates circadian rhythm, sleep, and other physiological events in the animal system, but it is now recognized as a multifunctional regulator in diverse life groups including invertebrates, plants, and procaryotes [4]. In plants, melatonin is involved in diverse physiological events (Table 1), including the mitigation of abiotic stress and the regulation of growth and development [5]. Furthermore, melatonin strengthens plant immunity and protects plants against pathogens, supplying new possibilities for agricultural application [6,7,8].
Melatonin has strong antioxidant activity of its own [9], so it can protect plants simply by being present at the right concentration. This intrinsic antioxidant activity is due to the chemical properties of the melatonin molecule, so that plant protection by high concentrations of melatonin is a response independent of cellular signaling. However, during evolution, plants appear to have acquired additional ways of using this substance that have been present since the earliest days of life on Earth [6]. By perceiving melatonin as a signal and linking it to intracellular signal transduction processes, plants were able to achieve a higher level of regulation through the amplification or diversification of responses to signal detection and integration of signals from different signaling pathways.

{kind=link}
{kind=link}
Table 1.
Examples of the pleiotropic effects of melatonin in plants.
| Plants | Responses | Effective Concentration | References |
|---|---|---|---|
| Arabidopsis | Alleviation of cold stress | 10, 30 µM | [10] |
| Upregulation of stress and defense genes and others | 1 mM | [11] | |
| Mediation of innate immunity against bacterial pathogens | 20 µM | [12] | |
| Expression of CBF/DREB1s genes involved in stress response | 50 µM | [13] | |
| Induction of nitric oxide and enhancement of innate immunity | 20 µM | [14] | |
| Cell wall strengthening and callose accumulation against bacteria | 50 µM | [15] | |
| Suppression of root meristem, auxin biosynthesis, and transport | 100 µM–1 mM | [16] | |
| Repression of the floral transition by stabilizing DELLA proteins | 0.5, 1.0 mM | [17] | |
| Improvement of iron deficiency tolerance | 5 µM | [18] | |
| Inhibition of brassinosteroid synthesis and decrease in hypocotyl growth | 0.1–1.0 mM | [19] | |
| Promotion of lateral root development (synergism with auxin) | 50–300 µM | [20] | |
| Regulation of stomatal closure | 0.1–80 µM | [21] | |
| Inhibition of seedling growth and regulation of abscisic acid homeostasis | 100, 300 µM | [22] | |
| Induction of pathogenesis-related proteins and other defense genes | 10 µM | [23] | |
| Activation of mitogen-activated protein kinases (MPK3, MPK6) | 1 µM | [24] | |
| Maize | Improvement of germination by priming seeds with melatonin | 50, 100 µM | [25] |
| Delay of leaf senescence and improvement of antioxidant defense | 25–75 µM | [26] | |
| Induction of resistance to a fungal pathogen, Fusarium graminearum | 50–400 µM | [27] | |
| Enhancement of thermotolerance through modulation of antioxidant defense | 10–70 µM | [28] | |
| Increase in drought stress tolerance | 0.25–1.0 mM | [29] | |
| Rice | Mitigation of cold-stress-induced reactive oxygen species (ROS) accumulation | 20, 100 µM | [30] |
| Regulation of root architecture and modulation of auxin response | 10–50 µM | [31] | |
| Suppression of a pathogenic bacterial growth in rice | 200 µg/mL | [32] | |
| Improvement of resistance to rice stripe virus | 0.1–10 µM | [33] | |
| Reduction of fluoride uptake and toxicity | 20 µM | [34] | |
| Broad-spectrum antifungal activity | 0.1–10 mM | [35] | |
| Soybean | Enhancement of growth and resistance to abiotic stress | 50, 100 µM | [36] |
| Activation of auxin biosynthesis and signal transduction | 20 µM | [37] | |
| Alleviation of salt-alkali stress by reducing oxidative damage of DNA | 300 µM | [38] | |
| Mitigation of arsenate stress | 100 µM | [39] | |
| Tomato | Promotion of adventitious root development | 50 µM | [40] |
| Improvement of tomato fruit quality and more ascorbic acid and lycopene | 0.1 mM | [41] | |
| Mitigation of acid rain stress and modulation of leaf ultrastructure | 50–250 µM | [42] | |
| Acclimation to a combination of abiotic stresses | 100 µM | [43] | |
| Alleviation of photosynthetic apparatus under cold stress | 5–250 µM | [44] | |
| Promotion of salicylic acid and nitric oxide accumulation and viral resistance | 50–400 µM | [45] | |
| Improvement of cadmium tolerance | 100 µM | [46] | |
| Delay of leaf senescence in darkness | 250 µM | [47] | |
| Alleviation of heat-indued damage by balancing redox homeostasis | 100 µM | [48] | |
| Improvement of cold tolerance | 100 µM | [49] | |
| Alleviation of nickel toxicity | 100 µM | [50] | |
| Ethylene-dependent enhancement of carotenoid biosynthesis | 50 µM | [51] | |
| Increase in the resistance to the fungal pathogen Botrytis cinerea | 1–100 µM | [52] | |
| Wheat | Mitigation of salt stress through modulation of polyamine metabolism | 1 µM | [53] |
| Increase in photosynthetic capacity and salt tolerance | 100 µM | [54] | |
| Enhancement of seed germination under salt stress | 50–250 µM | [55] | |
| Reduction of chromium uptake and toxicity | 1, 2 mM | [56] |
Because melatonin has existed as a biomolecule since the dawn of evolution, its role in plants is complex, ranging from simple chemical action to receptor-mediated signaling. While this situation is advantageous for plants, it creates considerable confusion for melatonin researchers. The discovery of the melatonin receptor in plants [21] has led to great advances in understanding how melatonin works as a signal molecule and leads to the suggestion of melatonin as a new phytohormone [57,58], but the details of the action mechanism still remain poorly understood.
This review examines the melatonin responses in plants and aims to identify which of these are characteristic of phytohormone responses. It examines the effective concentration, biosynthesis, catabolism, transport, and dose–response curves of melatonin in plants and compares them with those of other phytohormones. Additionally, this paper reviews the current state of knowledge regarding the molecular biology and biochemistry of the plant melatonin receptor. This review examines the adequacy of available information in explaining the function of the melatonin receptor and discusses future works.
2. Pleiotropy
A characteristic of phytohormones is their pleiotropic action. The pleiotropy of phytohormone action is probably one of the reasons that it is possible to regulate a wide range of physiological responses with a limited number of phytohormones. In plants, the effects of melatonin are pleiotropic (Table 1).
The most widely accepted effect of melatonin in plants is its activity against oxidative stress. While melatonin itself is a potent antioxidant, it has the interesting property of inducing the expression of genes that alleviate oxidative stress. Melatonin is also known to interact with reactive oxygen species (ROS) and reactive nitrogen species (RNS) [6]. Not only against oxidative stress but also against diverse environmental stresses, e.g., salt, heat, drought, heavy metal, and strong light, melatonin protects plants [59]. Initially, melatonin attracted attention for its alleviating effects on abiotic stress, but now its protective roles against biotic stress are being illustrated. Melatonin reduces damage caused by viral, bacterial, and fungal infections in plants [7], giving the possibility to protect plants using melatonin as a defense stimulator [60,61,62]. Melatonin also affects plant growth and development, including root development, hypocotyl growth, germination, flowering, and parthenocarpy [57]. In many cases, these pleiotropic effects of melatonin do not occur directly but through a network of interactions with other phytohormones, including auxin, gibberellin (GA), cytokinin, abscisic acid (ABA), ethylene, brassinosteroids, salicylic acid, and jasmonic acid [6,63,64]. Melatonin and other phytohormones seem to share their signaling pathways very often. Melatonin can also change the concentration of other phytohormones, and the inverse has been known, too [64]. Interestingly, melatonin increases the concentration of itself [65]. All of melatonin’s effects concerning abiotic stress [5,6,66,67,68,69,70], biotic stress [6,7,8,71,72], and growth and development [5,66,69,70] have been extensively reviewed in recent years.
3. Biosynthesis, Conjugation, and Catabolism
Melatonin is biosynthesized and catabolized like other phytohormones, and its metabolic pathways consist of a regulatory network together with other phytohormones [64]. Melatonin is synthesized in plants from tryptophan (Trp) mainly via trypamine, serotonin, and N-acetylserotonin [66,73]. However, under some environmental stresses, melatonin is produced via alternative pathways using 5-hydroxytryptophan or 5-methoxytryptophan or both as intermediates. Indole-3-acetic acid (IAA), the representative auxin with an indole ring like melatonin, is essentially produced from Trp via indole pyruvic acid in Arabidopsis [74]. In parallel, several biosynthetic pathways to produce IAA have been suggested in diverse plants, including even a Trp-independent pathway based on the increase in IAA in the Trp-deficient maize mutant orange pericarp [75]. Melatonin and auxin share some intermediates, and this is one reason for the close relationship between melatonin and auxin [37].
The biosynthesis of melatonin is a part of the regulatory network. For example, ABA stimulates melatonin biosynthesis in watermelon by inducing gene expression of ClASMT encoding N-acetylserotonin methyltransferase, ClCOMT encoding caffeic acid O-methyltransferase, and ClSNAT encoding serotonin N-acetyltransferase [76].
To meet the physiological demands by optimizing hormone levels, plants have evolved sophisticated regulatory mechanisms of biosynthesis, catabolism, and conjugation. Melatonin participates in the regulation of homeostasis of other hormones in various modes. When plants experience water stress, ABA levels sharply rise to cope with the drought by closing stomata [77]. The increased ABA is reduced through degradation and conjugation [77]. In contrast to the cooperative effect of melatonin and ABA on plant responses to environmental stress, melatonin contributes to the maintenance of hormonal balance in Malus plants by decreasing the expression of the NECD gene encoding 9-cis-epoxycarotenoid dioxygenase, which is involved in ABA biosynthesis, or by promoting the expression of enzymes that catabolize ABA [68,78]. Melatonin also affects GA biosynthesis. Active GA homologues are produced by GA-3 oxidase, and the increased hormone is inactivated by GA-2 oxidase [79]. Melatonin regulates this fine-tuning of GA biosynthesis [80], suggesting a complex network in the regulation of hormone levels.
Melatonin promotes ethylene biosynthesis during tomato fruit ripening by inducing the expression of both 1-aminocyclopropane-1-carboxylic acid (ACC) synthase and ACC oxidase, which are involved in ethylene biosynthesis [51]. In this case, the apparent phenotypic change is fruit ripening, but the direct effect of melatonin is on ethylene biosynthesis; therefore, the observed effect of melatonin is indirect. These results suggest that mechanistic studies are needed to understand the action of melatonin. In the ethylene biosynthesis described above, there is a lack of information on how melatonin is perceived and linked to gene expression, although it is known that a MYB transcription factor binds directly to the promoter of the ACC synthase gene in another plant, grapevine [81]. Melatonin also affects the biosynthesis of melatonin itself. Exogenous melatonin increased endogenous melatonin levels in cassava [82] and maize [83], suggesting that self-stimulatory mechanisms may operate under certain circumstances.
Not only biosynthesis, but also conjugation and catabolism, are important tools to regulate hormone levels. All phytohormones are active in free form and lose their activity when conjugated to sugars or amino acids, except jasmonic acid, which is only active when conjugated to isoleucine [84]. Conjugation of melatonin was expected [85], but it has not yet been demonstrated in plants. When melatonin encounters hydroxyl radicals, melatonin can be chemically oxidized. In addition, melatonin is also enzymatically oxidized by melatonin 2-hydroylase [86] and melatonin 3-hydroxylase [87] to 2-hydroxy- melatonin and 3-hydroxymelatonin, respectively. Catabolism generally inactivates phytohormones. 2-oxoglutarate-dependent Fe(II) dioxygenase (DIOXYGENASE FOR AUXIN OXIDATION; DAO) oxidizes IAA to 2-oxindole-3-acetic acid (oxIAA) [88]. C26 hydroxylase (encoded by BAS1) and some other enzymes inactivate brassinosteroids [89]. GA2-oxidase inactivates GA [79]. Interestingly, unlike the oxidation products of other hormones, oxidized melatonin still exhibits antioxidant activity, although the activity is reduced to about 50%.
Multiple crosstalk among the regulatory mechanisms of phytohormone levels is an important tool to harmonize the diverse responses to cope with complicated situations for efficient survival. Melatonin, like other phytohormones, plays a role in this regulatory network, although the detailed mechanism remains to be elucidated.
4. Transport
Transport is an important means of regulating the local distribution of plant hormones. In particular, the polar transport of auxin by influx and efflux carriers is of great importance for the establishment of polarity during plant development. Fine-tuning the direction of auxin transport by redistributing efflux carriers (PINs) during development is an efficient way to establish a new axis [90]. Strigolactone moves from root to shoot and regulates apical dominance by suppressing axillary bud growth [91]. Melatonin transport is still poorly understood. Melatonin supplied to maize roots accumulates in the leaves, and the amount of accumulated melatonin decreases as the stomata closes [92], so it appears that melatonin moves along the transpiration stream in the xylem, but the details are unknown.
5. Dose Relationships
5.1. Effective Concentrations
Hormones are regulators that work at low concentrations, but the question is what range of concentrations is meant by low in this case. To see if the concentrations at which melatonin works are within the range of phytohormones, the working concentrations of other phytohormones are checked as well as that of melatonin. Auxin shows activity in the range of 0.1–10 µM to promote cell elongation in maize coleoptiles [93]. Cytokinin suppresses Arabidopsis root growth in the range of 0.01–10 µM [94]. Melatonin shows activity on stomatal closure in the range of 0.1–40 µM [21]. However, some effects of melatonin appear at much higher concentrations, sometimes even higher than 1 mM [95,96]. These differences in effective concentrations of melatonin for different effects could be due to whether the effects are via receptor signaling or not. Melatonin is a powerful antioxidant and can directly counteract oxidants generated during plant stress without the need for receptors at high concentrations.
5.2. Patterns of Dose–Response Curves
In phytohormone science, “dose–response” generally means “concentration gradient response”. Numerically, the “hormone dose” can be assumed to be “the concentration of a given hormone” times “the duration of the treatment”, as the light fluence is calculated as the multiplication of “photons per unit area” and “the duration time”. However, such an assumption is physiologically inappropriate for phytohormones, because the treatment with 1 µM IAA for 10 h and that with 10 µM IAA for 1 h gave qualitatively and quantitatively different results in stem growth [93]. Therefore, concentration gradient response curves obtained under the same experimental conditions are generally accepted as dose–response curves for phytohormones.
Because the binding of a phytohormone to its receptor is an equilibrium between bound and unbound states of the molecules, most of the dose–response curves show changes over several orders of magnitude of the hormone concentrations [97]. Because dose–response curves of phytohormones reflect the status of receptor and changes in signal transduction, observation of dose–response curves is very useful to obtain information about the hormone action.
As shown by the dose–response curves for phytohormones, responses vary depending on the level of hormone, providing an efficient way to regulate hormone action (Table 1). They typically show a saturating sigmoid or inverse sigmoid curve when the hormone response is plotted against hormone concentration [98,99,100] or a bell-shaped characteristic [93,101] where the response peaks at a certain concentration and then declines. If the hormone response is suppressive, the bell-shaped dose–response curve can also be expressed as a U-shaped curve [21] (Table 2). The shape of dose–response curves for the same hormone can vary depending on the type of response, tissue, organ, plant species, and many other conditions.
The dose–response curve of melatonin for stomatal closure is U-shaped [21], but the curve for Arabidopsis root growth is inverse sigmoidal [16]. Because the dose–response curve of melatonin over 10−8 M to 10−4 M for coleoptile and root growth in some monocot plants was not simply bell-shaped and was even somewhat inhibitory at high concentrations [102], the character was interpreted as hormetic [103].
Table 2.
Types of dose–response curves for some phytohormones.
| Dose–Response | Hormone | Response | References |
|---|---|---|---|
| Sigmoidal | |||
 | Auxin | Elongation of maize coleoptiles and pea stems | [98] |
| Petiole elongation in Ranunculus sceleratus | [104] | ||
| Gibberellin | Leaf elongation in the dwarf mutants of barley | [99] | |
| Cytokinin | Amaranthin accumulation | [105] | |
| Strigolactone | Germination in some parasite plants | [106] | |
| Inverse sigmoidal | |||
 | Auxin | Root growth in Arabidopsis | [107] |
| Cytokinin | Root growth in Arabidopsis | [94] | |
| Abscisic acid | Germination in Arabidopsis | [108] | |
| Brassinosteroid | Growth of etiolated pea seedling | [109] | |
| Melatonin | Root growth in Arabidopsis | [16] | |
| Bell-shaped | |||
 | Auxin | Maize coleoptile elongation by IAA and 4-Cl-IAA | [101] |
| Pea epicotyl protoplast swelling | [110] | ||
| Maize coleoptile elongation | [93,111] | ||
| Strigolactone | Seed germination in some Striga plants | [112] | |
| Salicylic acid | PR1 accumulation in tobacco cell culture | [113] | |
| Melatonin | The maximum quantum yield of photosystem II (Fv/Fm) | [114] | |
| U-shaped | |||
 | Melatonin | Stomatal closure in Arabidopsis Malondialdehyde (MDA) content | [21,114] |
5.3. Changes in Dose–Response Curves and Regulatory Implications
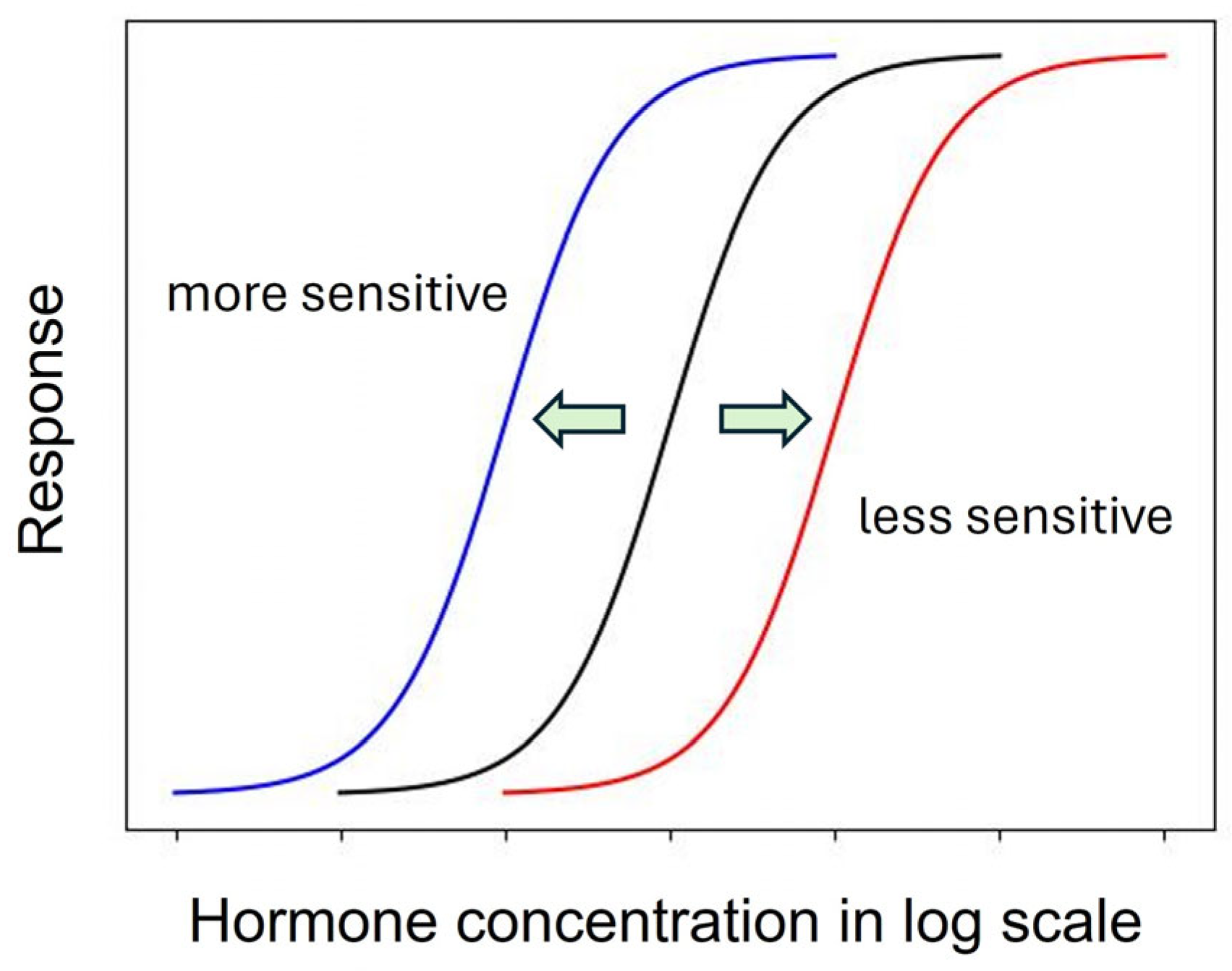
As described above, the shape of the dose–response curve is important, but another equally important characteristic is the location of the dose–response curve [97]. A dose–response curve on the left indicates a more sensitive response than a dose–response curve on the right (Figure 1).
A leftward shift of a dose–response curve has been suggested to indicate an increased affinity of the receptor for its ligand [115]. However, the rightward shift of the dose–response curve of root growth to auxin in the aux1 mutant [107], which has a defective auxin influx carrier, suggests that some cause other than a change in receptor affinity may induce the leftward or rightward shift of the dose–response curve.
Generally, this type of change is described by the term “sensitivity change”, which has a broad meaning and includes changes in affinity, receptivity (changes in the number of receptors), endogenous hormone levels, or any other physiological changes that affect hormone responses [115]. The dose–response curves of maize coleoptiles to auxin were different when treated with auxin 0.5 h or 2.5 h after tissue excision, indicating a time-dependent change in auxin sensitivity [116]. Furthermore, in a semiaquatic plant, Ranunculus sceleratus, petiole elongated auxin dependently, and the dose–response curve of the growth was shifted to the left by ethylene [104]. Interestingly, ethylene desensitization is mediated by a gene family, ARGOS, whose expression is induced by auxin in Arabidopsis [117]. Trewavas (1983) recognized such a great potential to regulate hormonal action by modulating sensitivity without changing phytohormone levels [75]. However, the regulation mechanism of phytohormone sensitivity remains largely unexplored. Despite its importance, the sensitivity regulation of melatonin action also remains to be investigated.
6. Receptors
6.1. Finding of Receptors and Disputes
Probably one of the most important proofs that a bioactive substance is a hormone is the presence of its receptor. Even after it was widely recognized that melatonin exists in plants and has a wide range of bioactivities, how plants perceive it remained a mystery. Therefore, the first paper suggesting that PMTR1 in Arabidopsis is the melatonin receptor [21] has received a lot of attention. PMTR1 was a CAND2 (candidate GPCR2) identified in Arabidopsis from previous studies, but its function was unknown [118]. The CAND2 protein (Q94AH1.1) consists of 300 amino acid residues, has a molecular weight of 34.1 kDa and an isoelectric point (pI) of 6.38, and contains seven transmembrane domains. Sequence analysis indicated that this protein belongs to a branch of GPCRs. Because the animal melatonin receptors, M1 and M2, are high-affinity GPCRs, the idea that the plant melatonin receptor is also a GPCR was intriguing. Since the first report, proteins with similarity to CAND2/PMTR1 have been identified in Panax notoginseng [119], Nicotiana benthamiana [120], Medicago sativa [121], Zea mays [122], and Gossypium hirsutum [123]. In CAND2/PMTR1 knock-out mutants or overexpressed plants, stomatal closure [21,124], immunity [119,120], osmotic stress tolerance [125], salt and osmotic stress tolerance [121,122] and mitochondrial gene expression [126] were affected, supporting the idea that the CAND2/PMTR1 is the actual phytomelatonin receptor. However, some researchers questioned the conclusion that CAND2/PMTR1 is a receptor for phytomelatonin because they obtained different results in Arabidopsis [127], whereas CAND2/PMTR1 was first reported to be the receptor for phytomelatonin [21].
The workers double-checking the first report observed slight differences in the localization of CAND2/PMTR1-mCherry and FLS-2-GFP (as a plasma membrane marker), melatonin-induced MAPK (MPK3/6) activation and several gene expressions in cand2 mutants, and MPK3/6 activation in the gpa1 mutant [127]. The first report showed the localization of CAND2/PMTR1-YFP and YFP (as a cytoplasmic marker) in separate cells without the use of a plasma membrane maker [21]. Unfortunately, the location of YFP was confusingly similar to that of CAND2/PMTR1-YFP [21], although the authors interpreted the result as confirming the plasma membrane localization of CAND2/PMTR1. Later, it was shown that the localization of GFP-PMTR1 and PIP2-mCherry (a plasma membrane marker) coincided with another group [119]. The plasma membrane localization of CAND2/PMTR1 was also confirmed in alfalfa [121].
The next argument was the activation of MPK3/6 by melatonin in the cand2-1 and cand 2-2 mutants [127]. The cand2-1 was used in the first report to show the aberrations in the stomatal closure and ion fluxes regulated by melatonin [21]. To investigate this point, the researchers who double-checked the first report measured the transcript level of CAND2 in cand2-1 and cand2-2 and found that the CAND2 mRNA accumulation was abolished in the cand2-2 but not in the cand2-1 [127], which was used to obtain evidence for the receptor function of CAND2/PMTR1 [21]. However, they did not check the CAND2/PMTR1 protein level. Because the transcript level does not necessarily correspond to the protein level, this question remains open.
Another argument was that melatonin-induced gene expression and MPK3/6 activation were normal in the gpa1 mutant [127]. GPA1 encodes Gα, which is part of a heterotrimeric GTP-binding protein that interacts with GPCRs and is known to play a key role in signal transduction. Because CAND2/PMTR1 was identified as a GPCR, the normal function of melatonin in the gpa1 mutant seemed to make no sense, casting doubt on CAND2/PMTR1 as the melatonin receptor. However, melatonin-induced activation of MPK3/6 was impaired in pmtr1 mutant but was normal in the gpa1-4 mutant [119], strongly suggesting the involvement of another signaling pathway that is GPA1-independent. In contrast, melatonin-induced stomatal closure was aberrant in the gpa1-4 mutant [119].
The evidence collected to date suggests that CAND2/PMTR1 acts as a receptor for phytomelatonin. However, if one were to add another question, it would be about the biological benefits of localizing the melatonin receptor to the plasma membrane. What would be the advantages of signal perception at the cell surface for melatonin, which easily passes through membranes?
6.2. Biochemistry of the Receptors
The main feature of a hormone receptor is binding to its ligand. The binding can be biochemically described following a saturating competitive binding assay. Several papers on CAND2/PMTR1 included the binding assay results and reported that the Kd of CAND2/TMPR1 was about 0.73 nM in Arabidopsis [37] and about 1.026 nM (with Bmax 0.93 pmol/6 nmol protein) in alfalfa [128]. Another paper reported the EC50 value of 47.8 nM in maize [122]. Unfortunately, the procedures for the binding assay and the results were described only briefly. Thus, it remains unclear how the melatonin was labeled, whether the equilibrium was properly achieved, whether there was adequate competition with unlabeled melatonin, how the specific binding was estimated, and how Kd and Bmax were determined. Because the molecular weight of the CAND2/PMTR1 protein is known, it should be possible to calculate the number of ligand binding sites on the receptor from Bmax, but the experimental results were presented too simplistically to make the necessary calculations.
One paper presented a nice saturation curve of melatonin binding to CAND2/PMTR1 as evidence for ligand binding to the candidate receptor in cassava [128]. The shape of the saturation curve looked similar to that observed for other hormone binding but was in fact unique when the fold change of melatonin from basal to saturation level was considered. In their hands, the binding was saturated when melatonin was increased from 0.5 to 1.5 nmol in the presence of 20 mmol of purified MePMTR1 [128]. In general, saturation of ligand binding to a hormone receptor occurs over at least two or more orders of magnitude of increase in ligand concentration. Therefore, it is very difficult to observe the saturation of ligand binding with only the labeled ligand due to the difficulty of preparing high concentrations of labeled ligand. Instead, the binding characteristics of a ligand to its receptor are measured by preparing a certain concentration of labeled ligand and allowing it to compete with various concentrations of unlabeled ligand for binding to the receptor. The competitor ligand decreases the measured indicator of the labeled ligand, such as radioactivity, in a concentration-dependent manner. However, the specific activity of the ligand mixture provides the actual amount of increasing ligand binding to the receptor, including both the labeled and unlabeled ligand. The binding saturation occurs over several orders of magnitude of the ligand. Unfortunately, the description of the unique aspect of melatonin binding to MePMTR1 in a narrow range was omitted in the paper [128].
The long debate over auxin-binding protein 1 (ABP1) [129,130,131,132,133,134] shows that hormone-binding proteins must be thoroughly validated before they can be accepted as receptors. After the first discovery of auxin binding to membranes [135], efforts were made to find proteins that specifically bind to auxin. A protein isolated from maize shoots, which have been extensively used to monitor auxin activities, showed specific auxin binding with a Kd value of 6 × 10−8 M [136]. Finally, the gene for the auxin-binding protein was cloned, and the molecular characteristics of ABP1 were revealed [137]. ABP1 had a molecular mass of 22 kDa and contained the endoplasmic reticulum retention signal KDEL, which could explain the fact that auxin binds to the endoplasmic reticulum, as reported in previous papers [138]. However, based on discrepancies between the dose–response curve of 1-naphthalene acetic acid (1-NAA; an artificial auxin) and the Kd value of ABP1, along with several other uncertainties, skeptical questions have been raised about the receptor function of ABP1, even calling it a red herring [129] or an outsider [131]. Further molecular biological and biochemical evidence supporting the physiological function of ABP1 has been presented and repeatedly rejected [134], leaving the role of ABP1 as an auxin receptor uncertain. As seen in the ABP1 debates, further efforts are needed to confirm the physiological function of CAND2/PMTR1 as a phytomelatonin receptor.
To establish the physiologically relevant melatonin receptor function of CAND2/PMTR1, the first step would be an elaborated biochemical description of the binding properties, followed by a good match between the biochemical properties of the hormone-binding protein and the molecular physiological responses induced by melatonin. With regard to this point, I still have questions about the reason for the difference between the melatonin dose–response and the melatonin binding characteristics of CAND2/PMTR1; for example, stomatal closure showed maximum activity at 1 to 10 µM melatonin, but melatonin binding was already saturated at 100 nM [21]. The next step would be to determine whether the binding of melatonin alters the conformation of CAND2/PMTR1 and whether these changes are associated with subsequent signaling. In particular, the interaction between CAND2/PMTR1 and GPA1 is important because there are some conflicting results. Scrutinizing the action of GPA1 may provide new clues that lead us to a different signaling pathway or even an unexpected new receptor.
7. Perspectives
Phytomelatonin is biosynthesized and degraded in plants and induces a variety of responses. Some of the responses induced by low concentrations of melatonin show characteristics of hormone responses, while other responses to high concentrations of melatonin may be due to the chemical nature of melatonin. One of the most critical criteria for recognizing melatonin as a phytohormone is the existence of the receptor. CAND2/PMTR1 is currently considered the primary candidate for the phytomelatonin receptor. Mutations in CAND2/PMTR1 have been found to either abolish or reduce several melatonin-induced responses in plants, indicating its critical involvement in melatonin actions. However, there are melatonin responses that cannot be explained by the general action mode of the G protein-coupled receptors to which CAND2/PMTR1 belongs. There may be signaling pathways of phytomelatonin that are dependent on GPA1, and others that are independent of it (Figure 2). Furthermore, the signaling mechanisms for many other physiological responses regulated by CAND2/PMTR1 remain unclear. To establish CAND2/PMTR1 as a receptor for phytomelatonin, it is necessary to thoroughly characterize its biochemical properties. Then, the mechanisms of signal transduction that induce the molecular physiological responses to melatonin must be elucidated. The melatonin responses in plants that are not accounted for by CAND2/PMTR1 may be attributed to the chemical properties of melatonin itself. However, it is also possible that plants have a distinct signaling process that differs from the classical GPCR signaling. It is also possible that plants even have another type of phytomelatonin receptor in addition to CAND2/PMTR1.
Funding
The present research was supported by the research fund of Dankook University in 2021.
Acknowledgments
I would like to thank Jae Yong Park for his great help in line drawings.
Conflicts of Interest
The author declares no conflict of interest.
References
- Dubbels, R.; Reiter, R.J.; Klenke, E.; Goebel, A.; Schnakenberg, E.; Ehlers, C.; Schiwara, H.W.; Schloot, W. Melatonin in edible plants identified by radioimmunoassay and by high performance liquid chromatography-mass spectrometry. J. Pineal Res. 1995, 18, 28–31. [Google Scholar] [CrossRef]
- Hattori, A.; Migitaka, H.; Iigo, M.; Itoh, M.; Yamamoto, K.; Ohtani-Kaneko, R.; Hara, M.; Suzuki, T.; Reiter, R.J. Identification of melatonin in plants and its effects on plasma melatonin levels and binding to melatonin receptors in vertebrates. Biochem. Mol. Biol. Int. 1995, 35, 627–634. [Google Scholar]
- Kolar, J.; Johnson, C.H.; Machackova, I. Presence and possible role of melatonin in a short-day flowering plant, Chenopodium rubrum. Adv. Exp. Med. Biol. 1999, 460, 391–393. [Google Scholar]
- Reiter, R.J.; Sharma, R.; Rosales-Corral, S.; de Campos Zuccari, D.A.P.; de Almeida Chuffa, L.G. Melatonin: A mitochondrial resident with a diverse skill set. Life Sci. 2022, 301, 120612. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Xu, X.; Li, L.; Sun, Q.; Wang, Q.; Huang, H.; Tong, Z.; Zhang, J. Melatonin-mediated development and abiotic stress tolerance in plants. Front. Plant Sci. 2023, 14, 1100827. [Google Scholar] [CrossRef]
- Khan, M.S.S.; Ahmed, S.; Ikram, A.U.; Hannan, F.; Yasin, M.U.; Wang, J.; Zhao, B.; Islam, F.; Chen, J. Phytomelatonin: A key regulator of redox and phytohormones signaling against biotic/abiotic stresses. Redox Biol. 2023, 64, 102805. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Ruiz, J.; Giraldo-Acosta, M.; El Mihyaoui, A.; Cano, A.; Arnao, M.B. Melatonin as a Possible Natural Anti-Viral Compound in Plant Biocontrol. Plants 2023, 12, 781. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Lal, M.K.; Kumar, R.; Mangal, V.; Altaf, M.A.; Sharma, S.; Singh, B.; Kumar, M. Insight into melatonin-mediated response and signaling in the regulation of plant defense under biotic stress. Plant Mol. Biol. 2022, 109, 385–399. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Tan, D.X.; Reiter, R.J. Melatonin as a natural ally against oxidative stress: A physicochemical examination. J. Pineal Res. 2011, 51, 1–16. [Google Scholar] [CrossRef]
- Bajwa, V.S.; Shukla, M.R.; Sherif, S.M.; Murch, S.J.; Saxena, P.K. Role of melatonin in alleviating cold stress in Arabidopsis thaliana. J. Pineal Res. 2014, 56, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Weeda, S.; Zhang, N.; Zhao, X.; Ndip, G.; Guo, Y.; Buck, G.A.; Fu, C.; Ren, S. Arabidopsis transcriptome analysis reveals key roles of melatonin in plant defense systems. PLoS ONE 2014, 9, e93462. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Tan, D.X.; Reiter, R.J.; Shi, H. Comparative metabolomic analysis highlights the involvement of sugars and glycerol in melatonin-mediated innate immunity against bacterial pathogen in Arabidopsis. Sci. Rep. 2015, 5, 15815. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Qian, Y.; Tan, D.X.; Reiter, R.J.; He, C. Melatonin induces the transcripts of CBF/DREB1s and their involvement in both abiotic and biotic stresses in Arabidopsis. J. Pineal Res. 2015, 59, 334–342. [Google Scholar] [CrossRef]
- Shi, H.; Chen, Y.; Tan, D.X.; Reiter, R.J.; Chan, Z.; He, C. Melatonin induces nitric oxide and the potential mechanisms relate to innate immunity against bacterial pathogen infection in Arabidopsis. J. Pineal Res. 2015, 59, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Xu, L.; Su, T.; Jiang, Y.; Hu, L.; Ma, F. Melatonin regulates carbohydrate metabolism and defenses against Pseudomonas syringae pv. tomato DC3000 infection in Arabidopsis thaliana. J. Pineal Res. 2015, 59, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; An, B.; Wei, Y.; Reiter, R.J.; Shi, H.; Luo, H.; He, C. Melatonin Regulates Root Meristem by Repressing Auxin Synthesis and Polar Auxin Transport in Arabidopsis. Front. Plant Sci. 2016, 7, 1882. [Google Scholar] [CrossRef]
- Shi, H.; Wei, Y.; Wang, Q.; Reiter, R.J.; He, C. Melatonin mediates the stabilization of DELLA proteins to repress the floral transition in Arabidopsis. J. Pineal Res. 2016, 60, 373–379. [Google Scholar] [CrossRef]
- Zhou, C.; Liu, Z.; Zhu, L.; Ma, Z.; Wang, J.; Zhu, J. Exogenous Melatonin Improves Plant Iron Deficiency Tolerance via Increased Accumulation of Polyamine-Mediated Nitric Oxide. Int. J. Mol. Sci. 2016, 17, 1777. [Google Scholar] [CrossRef]
- Xiong, F.; Zhuo, F.; Reiter, R.J.; Wang, L.; Wei, Z.; Deng, K.; Song, Y.; Qanmber, G.; Feng, L.; Yang, Z.; et al. Hypocotyl Elongation Inhibition of Melatonin Is Involved in Repressing Brassinosteroid Biosynthesis in Arabidopsis. Front. Plant Sci. 2019, 10, 1082. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Rutto, L.; Katuuramu, D. Melatonin acts synergistically with auxin to promote lateral root development through fine tuning auxin transport in Arabidopsis thaliana. PLoS ONE 2019, 14, e0221687. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Li, D.X.; Zhang, J.R.; Shan, C.; Rengel, Z.; Song, Z.B.; Chen, Q. Phytomelatonin receptor PMTR1-mediated signaling regulates stomatal closure in Arabidopsis thaliana. J. Pineal Res. 2018, 65, e12500. [Google Scholar] [CrossRef]
- Yin, X.; Bai, Y.L.; Gong, C.; Song, W.; Wu, Y.; Ye, T.; Feng, Y.Q. The phytomelatonin receptor PMTR1 regulates seed development and germination by modulating abscisic acid homeostasis in Arabidopsis thaliana. J. Pineal Res. 2022, 72, e12797. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Byeon, Y.; Back, K. Melatonin as a signal molecule triggering defense responses against pathogen attack in Arabidopsis and tobacco. J. Pineal Res. 2014, 57, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Back, K. Mitogen-activated protein kinase pathways are required for melatonin-mediated defense responses in plants. J. Pineal Res. 2016, 60, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Li, G.; Cui, Z.; Yang, F.; Jiang, X.; Diallo, L.; Kong, F. Seed Priming with Melatonin Improves the Seed Germination of Waxy Maize under Chilling Stress via Promoting the Antioxidant System and Starch Metabolism. Sci. Rep. 2019, 9, 15044. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Su, W.; Kamran, M.; Ahmad, I.; Meng, X.; Wu, X.; Javed, T.; Han, Q. Foliar application of melatonin delay leaf senescence in maize by improving the antioxidant defense system and enhancing photosynthetic capacity under semi-arid regions. Protoplasma 2020, 257, 1079–1092. [Google Scholar] [CrossRef] [PubMed]
- Kong, M.; Ali, Q.; Jing, H.; Hussain, A.; Wang, F.; Liu, X.; Gao, X.; Xu, H.L. Exogenous Melatonin Regulates Plant-Disease Interaction by Inducing Maize Resistance and Decreasing the Pathogenicity of Fusarium graminearum. Physiol. Plant 2023, 175, e14108. [Google Scholar] [CrossRef]
- Li, Z.G.; Xu, Y.; Bai, L.K.; Zhang, S.Y.; Wang, Y. Melatonin enhances thermotolerance of maize seedlings (Zea mays L.) by modulating antioxidant defense, methylglyoxal detoxification, and osmoregulation systems. Protoplasma 2019, 256, 471–490. [Google Scholar] [CrossRef]
- Muhammad, I.; Yang, L.; Ahmad, S.; Farooq, S.; Khan, A.; Muhammad, N.; Ullah, S.; Adnan, M.; Ali, S.; Liang, Q.P.; et al. Melatonin-priming enhances maize seedling drought tolerance by regulating the antioxidant defense system. Plant Physiol. 2023, 191, 2301–2315. [Google Scholar] [CrossRef]
- Han, Q.H.; Huang, B.; Ding, C.B.; Zhang, Z.W.; Chen, Y.E.; Hu, C.; Zhou, L.J.; Huang, Y.; Liao, J.Q.; Yuan, S.; et al. Effects of Melatonin on Anti-oxidative Systems and Photosystem II in Cold-Stressed Rice Seedlings. Front. Plant Sci. 2017, 8, 785. [Google Scholar] [CrossRef]
- Liang, C.; Li, A.; Yu, H.; Li, W.; Liang, C.; Guo, S.; Zhang, R.; Chu, C. Melatonin Regulates Root Architecture by Modulating Auxin Response in Rice. Front. Plant Sci. 2017, 8, 134. [Google Scholar] [CrossRef]
- Chen, X.; Sun, C.; Laborda, P.; Zhao, Y.; Palmer, I.; Fu, Z.Q.; Qiu, J.; Liu, F. Melatonin Treatment Inhibits the Growth of Xanthomonas oryzae pv. oryzae. Front. Microbiol. 2018, 9, 2280. [Google Scholar] [CrossRef]
- Lu, R.; Liu, Z.; Shao, Y.; Sun, F.; Zhang, Y.; Cui, J.; Zhou, Y.; Shen, W.; Zhou, T. Melatonin is responsible for rice resistance to rice stripe virus infection through a nitric oxide-dependent pathway. Virol. J. 2019, 16, 141. [Google Scholar] [CrossRef]
- Banerjee, A.; Roychoudhury, A. Melatonin application reduces fluoride uptake and toxicity in rice seedlings by altering abscisic acid, gibberellin, auxin and antioxidant homeostasis. Plant Physiol. Biochem. 2019, 145, 164–173. [Google Scholar] [CrossRef]
- Li, R.; Bi, R.; Cai, H.; Zhao, J.; Sun, P.; Xu, W.; Zhou, Y.; Yang, W.; Zheng, L.; Chen, X.L.; et al. Melatonin functions as a broad-spectrum antifungal by targeting a conserved pathogen protein kinase. J. Pineal Res. 2023, 74, e12839. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Li, Q.T.; Chu, Y.N.; Reiter, R.J.; Yu, X.M.; Zhu, D.H.; Zhang, W.K.; Ma, B.; Lin, Q.; Zhang, J.S.; et al. Melatonin enhances plant growth and abiotic stress tolerance in soybean plants. J. Exp. Bot. 2015, 66, 695–707. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Tao, J.J.; Yin, C.C.; Chen, S.Y.; Zhang, J.S.; Zhang, W.K. Melatonin regulates gene expressions through activating auxin synthesis and signaling pathways. Front. Plant Sci. 2022, 13, 1057993. [Google Scholar] [CrossRef]
- Zhao, Q.; Shen, W.; Gu, Y.; Hu, J.; Ma, Y.; Zhang, X.; Du, Y.; Zhang, Y.; Du, J. Exogenous melatonin mitigates saline-alkali stress by decreasing DNA oxidative damage and enhancing photosynthetic carbon metabolism in soybean (Glycine max [L.] Merr.) leaves. Physiol. Plant 2023, 175, e13983. [Google Scholar] [CrossRef] [PubMed]
- Bhat, J.A.; Faizan, M.; Bhat, M.A.; Huang, F.; Yu, D.; Ahmad, A.; Bajguz, A.; Ahmad, P. Defense interplay of the zinc-oxide nanoparticles and melatonin in alleviating the arsenic stress in soybean (Glycine max L.). Chemosphere 2022, 288, 132471. [Google Scholar] [CrossRef] [PubMed]
- Wen, D.; Gong, B.; Sun, S.; Liu, S.; Wang, X.; Wei, M.; Yang, F.; Li, Y.; Shi, Q. Promoting Roles of Melatonin in Adventitious Root Development of Solanum lycopersicum L. by Regulating Auxin and Nitric Oxide Signaling. Front. Plant Sci. 2016, 7, 718. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, R.; Sun, Y.; Liu, Z.; Jin, W.; Sun, Y. The beneficial effects of exogenous melatonin on tomato fruit properties. Sci. Hortic. 2016, 207, 14–20. [Google Scholar] [CrossRef]
- Debnath, B.; Hussain, M.; Irshad, M.; Mitra, S.; Li, M.; Liu, S.; Qiu, D. Exogenous Melatonin Mitigates Acid Rain Stress to Tomato Plants through Modulation of Leaf Ultrastructure, Photosynthesis and Antioxidant Potential. Molecules 2018, 23, 388. [Google Scholar] [CrossRef] [PubMed]
- Martinez, V.; Nieves-Cordones, M.; Lopez-Delacalle, M.; Rodenas, R.; Mestre, T.C.; Garcia-Sanchez, F.; Rubio, F.; Nortes, P.A.; Mittler, R.; Rivero, R.M. Tolerance to Stress Combination in Tomato Plants: New Insights in the Protective Role of Melatonin. Molecules 2018, 23, 535. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xu, H.; Li, D.; Gao, X.; Li, T.; Wang, R. Effect of melatonin priming on photosynthetic capacity of tomato leaves under low-temperature stress. Photosynthetica 2018, 56, 884–892. [Google Scholar] [CrossRef]
- Zhao, L.; Chen, L.; Gu, P.; Zhan, X.; Zhang, Y.; Hou, C.; Wu, Z.; Wu, Y.F.; Wang, Q.C. Exogenous application of melatonin improves plant resistance to virus infection. Plant Pathol. 2019, 68, 1287–1295. [Google Scholar] [CrossRef]
- Hasan, M.K.; Ahammed, G.J.; Sun, S.; Li, M.; Yin, H.; Zhou, J. Melatonin Inhibits Cadmium Translocation and Enhances Plant Tolerance by Regulating Sulfur Uptake and Assimilation in Solanum lycopersicum L. J. Agric. Food Chem. 2019, 67, 10563–10576. [Google Scholar] [CrossRef]
- Wang, K.; Cai, S.; Xing, Q.; Qi, Z.; Fotopoulos, V.; Yu, J.; Zhou, J. Melatonin delays dark-induced leaf senescence by inducing miR171b expression in tomato. J. Pineal Res. 2022, 72, e12792. [Google Scholar] [CrossRef]
- Jahan, M.S.; Shu, S.; Wang, Y.; Chen, Z.; He, M.; Tao, M.; Sun, J.; Guo, S. Melatonin alleviates heat-induced damage of tomato seedlings by balancing redox homeostasis and modulating polyamine and nitric oxide biosynthesis. BMC Plant Biol. 2019, 19, 414. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Luo, Z.; Jannatizadeh, A.; Sheikh-Assadi, M.; Sharafi, Y.; Farmani, B.; Fard, J.R.; Razavi, F. Employing exogenous melatonin applying confers chilling tolerance in tomato fruits by upregulating ZAT2/6/12 giving rise to promoting endogenous polyamines, proline, and nitric oxide accumulation by triggering arginine pathway activity. Food Chem. 2019, 275, 549–556. [Google Scholar] [CrossRef]
- Jahan, M.S.; Guo, S.; Baloch, A.R.; Sun, J.; Shu, S.; Wang, Y.; Ahammed, G.J.; Kabir, K.; Roy, R. Melatonin alleviates nickel phytotoxicity by improving photosynthesis, secondary metabolism and oxidative stress tolerance in tomato seedlings. Ecotoxicol. Environ. Saf. 2020, 197, 110593. [Google Scholar] [CrossRef]
- Sun, Q.; Liu, L.; Zhang, L.; Lv, H.; He, Q.; Guo, L.; Zhang, X.; He, H.; Ren, S.; Zhang, N.; et al. Melatonin promotes carotenoid biosynthesis in an ethylene-dependent manner in tomato fruits. Plant Sci. 2020, 298, 110580. [Google Scholar] [CrossRef]
- Liu, C.; Chen, L.; Zhao, R.; Li, R.; Zhang, S.; Yu, W.; Sheng, J.; Shen, L. Melatonin Induces Disease Resistance to Botrytis cinerea in Tomato Fruit by Activating Jasmonic Acid Signaling Pathway. J. Agric. Food Chem. 2019, 67, 6116–6124. [Google Scholar] [CrossRef] [PubMed]
- Ke, Q.; Ye, J.; Wang, B.; Ren, J.; Yin, L.; Deng, X.; Wang, S. Melatonin Mitigates Salt Stress in Wheat Seedlings by Modulating Polyamine Metabolism. Front. Plant Sci. 2018, 9, 914. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Wang, J.; Lu, Z.; Liu, R.; Hong, Y.; Su, B.; Wang, Y.; Peng, Z.; Yu, C.; Gao, Y.; et al. Melatonin-Mediated Enhancement of Photosynthetic Capacity and Photoprotection Improves Salt Tolerance in Wheat. Plants 2023, 12, 3984. [Google Scholar] [CrossRef]
- Wang, J.; Lv, P.; Yan, D.; Zhang, Z.; Xu, X.; Wang, T.; Wang, Y.; Peng, Z.; Yu, C.; Gao, Y.; et al. Exogenous Melatonin Improves Seed Germination of Wheat (Triticum aestivum L.) under Salt Stress. Int. J. Mol. Sci. 2022, 23, 8436. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Ali, S.; Refay, Y.; Rizwan, M.; Alhammad, B.A.; El-Hendawy, S.E. Chromium resistant microbes and melatonin reduced Cr uptake and toxicity, improved physio-biochemical traits and yield of wheat in contaminated soil. Chemosphere 2020, 250, 126239. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernandez-Ruiz, J. Melatonin: A New Plant Hormone and/or a Plant Master Regulator? Trends Plant Sci. 2019, 24, 38–48. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Is Phytomelatonin a New Plant Hormone? Agronomy 2020, 10, 95. [Google Scholar] [CrossRef]
- Khan, T.A.; Fariduddin, Q.; Nazir, F.; Saleem, M. Melatonin in business with abiotic stresses in plants. Physiol. Mol. Biol. Plants 2020, 26, 1931–1944. [Google Scholar] [CrossRef]
- Li, L.; Du, C.; Wang, L.; Lai, M.; Fan, H. Exogenous melatonin improves the resistance to cucumber bacterial angular leaf spot caused by Pseudomonas syringae pv. Lachrymans. Physiol. Plant 2022, 174, e13724. [Google Scholar] [CrossRef]
- Li, C.; Zhao, Q.; Gao, T.; Wang, H.; Zhang, Z.; Liang, B.; Wei, Z.; Liu, C.; Ma, F. The mitigation effects of exogenous melatonin on replant disease in apple. J. Pineal Res. 2018, 65, e12523. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Wang, P.; Li, M.; Ke, X.; Li, C.; Liang, D.; Wu, S.; Ma, X.; Li, C.; Zou, Y.; et al. Exogenous melatonin improves Malus resistance to Marssonina apple blotch. J. Pineal Res. 2013, 54, 426–434. [Google Scholar] [CrossRef]
- Khan, M.; Ali, S.; Manghwar, H.; Saqib, S.; Ullah, F.; Ayaz, A.; Zaman, W. Melatonin Function and Crosstalk with Other Phytohormones under Normal and Stressful Conditions. Genes 2022, 13, 1699. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernandez-Ruiz, J. Melatonin as a regulatory hub of plant hormone levels and action in stress situations. Plant Biol. 2021, 23 (Suppl. S1), 7–19. [Google Scholar] [CrossRef]
- Jahan, M.S.; Shu, S.; Wang, Y.; Hasan, M.M.; El-Yazied, A.A.; Alabdallah, N.M.; Hajjar, D.; Altaf, M.A.; Sun, J.; Guo, S. Melatonin Pretreatment Confers Heat Tolerance and Repression of Heat-Induced Senescence in Tomato Through the Modulation of ABA- and GA-Mediated Pathways. Front. Plant Sci. 2021, 12, 650955. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tanveer, M.; Wang, H.; Arnao, M.B. Melatonin as a key regulator in seed germination under abiotic stress. J. Pineal Res. 2024, 76, e12937. [Google Scholar] [CrossRef]
- Colombage, R.; Singh, M.B.; Bhalla, P.L. Melatonin and Abiotic Stress Tolerance in Crop Plants. Int. J. Mol. Sci. 2023, 24, 7447. [Google Scholar] [CrossRef]
- Ali, M.; Pan, Y.; Liu, H.; Cheng, Z. Melatonin interaction with abscisic acid in the regulation of abiotic stress in Solanaceae family plants. Front. Plant Sci. 2023, 14, 1271137. [Google Scholar] [CrossRef]
- Ahmad, I.; Zhu, G.; Zhou, G.; Liu, J.; Younas, M.U.; Zhu, Y. Melatonin Role in Plant Growth and Physiology under Abiotic Stress. Int. J. Mol. Sci. 2023, 24, 8759. [Google Scholar] [CrossRef]
- Ahmad, I.; Song, X.; Hussein Ibrahim, M.E.; Jamal, Y.; Younas, M.U.; Zhu, G.; Zhou, G.; Adam Ali, A.Y. The role of melatonin in plant growth and metabolism, and its interplay with nitric oxide and auxin in plants under different types of abiotic stress. Front. Plant Sci. 2023, 14, 1108507. [Google Scholar] [CrossRef]
- Moustafa-Farag, M.; Almoneafy, A.; Mahmoud, A.; Elkelish, A.; Arnao, M.B.; Li, L.; Ai, S. Melatonin and Its Protective Role against Biotic Stress Impacts on Plants. Biomolecules 2019, 10, 54. [Google Scholar] [CrossRef]
- Zeng, H.; Bai, Y.; Wei, Y.; Reiter, R.J.; Shi, H. Phytomelatonin as a central molecule in plant disease resistance. J. Exp. Bot. 2022, 73, 5874–5885. [Google Scholar] [CrossRef]
- Khan, D.; Cai, N.; Zhu, W.; Li, L.; Guan, M.; Pu, X.; Chen, Q. The role of phytomelatonin receptor 1-mediated signaling in plant growth and stress response. Front. Plant Sci. 2023, 14, 1142753. [Google Scholar] [CrossRef]
- Cao, X.; Yang, H.; Shang, C.; Ma, S.; Liu, L.; Cheng, J. The Roles of Auxin Biosynthesis YUCCA Gene Family in Plants. Int. J. Mol. Sci. 2019, 20, 6343. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.D.; Sampson, M.B.; Neuffer, M.G.; Michalczuk, L.; Slovin, J.P.; Cohen, J.D. Indole-3-Acetic Acid Biosynthesis in the Mutant Maize Orange pericarp, a Tryptophan Auxotroph. Science 1991, 254, 998–1000. [Google Scholar] [CrossRef]
- Yan, M.; Li, M.; Ding, Z.; Qiao, F.; Jiang, X. Plant Hormone Signals Mediate Melatonin Synthesis to Enhance Osmotic Stress Tolerance in Watermelon Cells. Horticulturae 2023, 9, 927. [Google Scholar] [CrossRef]
- Ma, Y.; Cao, J.; He, J.; Chen, Q.; Li, X.; Yang, Y. Molecular Mechanism for the Regulation of ABA Homeostasis During Plant Development and Stress Responses. Int. J. Mol. Sci. 2018, 19, 3643. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Tan, D.-X.; Liang, D.; Chang, C.; Jia, D.; Ma, F. Melatonin mediates the regulation of ABA metabolism, free-radical scavenging, and stomatal behaviour in two Malus species under drought stress. J. Exp. Bot. 2015, 66, 669–680. [Google Scholar] [CrossRef]
- Hedden, P. The Current Status of Research on Gibberellin Biosynthesis. Plant Cell Physiol. 2020, 61, 1832–1849. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Zhang, N.; Yang, R.C.; Wang, L.; Sun, Q.Q.; Li, D.B.; Cao, Y.Y.; Weeda, S.; Zhao, B.; Ren, S.; et al. Melatonin promotes seed germination under high salinity by regulating antioxidant systems, ABA and GA₄ interaction in cucumber (Cucumis sativus L.). J. Pineal Res. 2014, 57, 269–279. [Google Scholar] [CrossRef]
- Xu, L.; Xiang, G.; Sun, Q.; Ni, Y.; Jin, Z.; Gao, S.; Yao, Y. Melatonin enhances salt tolerance by promoting MYB108A-mediated ethylene biosynthesis in grapevines. Hortic. Res. 2019, 6, 114. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Tie, W.; Ou, W.; Yan, Y.; Kong, H.; Zuo, J.; Ding, X.; Ding, Z.; Liu, Y.; Wu, C.; et al. Crosstalk between calcium and melatonin affects postharvest physiological deterioration and quality loss in cassava. Postharvest Biol. Technol. 2018, 140, 42–49. [Google Scholar] [CrossRef]
- Huang, B.; Chen, Y.-E.; Zhao, Y.-Q.; Ding, C.-B.; Liao, J.-Q.; Hu, C.; Zhou, L.-J.; Zhang, Z.-W.; Yuan, S.; Yuan, M. Exogenous Melatonin Alleviates Oxidative Damages and Protects Photosystem II in Maize Seedlings Under Drought Stress. Front. Plant Sci. 2019, 10, 677. [Google Scholar] [CrossRef]
- Monte, I. Jasmonates and salicylic acid: Evolution of defense hormones in land plants. Curr. Opin. Plant Biol. 2023, 76, 102470. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Functions of melatonin in plants: A review. J. Pineal Res. 2015, 59, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Byeon, Y.; Lee, H.Y.; Hwang, O.J.; Lee, H.J.; Lee, K.; Back, K. Coordinated regulation of melatonin synthesis and degradation genes in rice leaves in response to cadmium treatment. J. Pineal Res. 2015, 58, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Zawadzka, A.; Czarnocki, Z.; Reiter, R.J.; Back, K. Molecular cloning of melatonin 3-hydroxylase and its production of cyclic 3-hydroxymelatonin in rice (Oryza sativa). J. Pineal Res. 2016, 61, 470–478. [Google Scholar] [CrossRef]
- Zhang, J.; Lin, J.E.; Harris, C.; Campos Mastrotti Pereira, F.; Wu, F.; Blakeslee, J.J.; Peer, W.A. DAO1 catalyzes temporal and tissue-specific oxidative inactivation of auxin in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2016, 113, 11010–11015. [Google Scholar] [CrossRef]
- Wei, Z.; Li, J. Regulation of Brassinosteroid Homeostasis in Higher Plants. Front. Plant Sci. 2020, 11, 583622. [Google Scholar] [CrossRef]
- Marhava, P. Recent developments in the understanding of PIN polarity. New Phytol. 2022, 233, 624–630. [Google Scholar] [CrossRef]
- Mashiguchi, K.; Seto, Y.; Yamaguchi, S. Strigolactone biosynthesis, transport and perception. Plant J. 2021, 105, 335–350. [Google Scholar] [CrossRef] [PubMed]
- Yoon, Y.H.; Kim, M.; Park, W.J. Foliar Accumulation of Melatonin Applied to the Roots of Maize (Zea mays) Seedlings. Biomolecules 2019, 9, 26. [Google Scholar] [CrossRef] [PubMed]
- Polak, M.; Tukaj, Z.; Karcz, W. Effect of temperature on the dose–response curves for auxin-induced elongation growth in maize coleoptile segments. Acta Physiol. Plant. 2011, 33, 437–442. [Google Scholar] [CrossRef]
- Berkova, V.; Kameniarova, M.; Ondriskova, V.; Berka, M.; Mensikova, S.; Kopecka, R.; Luklova, M.; Novak, J.; Spichal, L.; Rashotte, A.M.; et al. Arabidopsis Response to Inhibitor of Cytokinin Degradation INCYDE: Modulations of Cytokinin Signaling and Plant Proteome. Plants 2020, 9, 1563. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zheng, X.; Reiter, R.J.; Feng, S.; Wang, Y.; Liu, S.; Jin, L.; Li, Z.; Datla, R.; Ren, M. Melatonin Attenuates Potato Late Blight by Disrupting Cell Growth, Stress Tolerance, Fungicide Susceptibility and Homeostasis of Gene Expression in Phytophthora infestans. Front. Plant Sci. 2017, 8, 1993. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Han, Y.; Yuan, X.; Zhang, M.; Li, P.; Ding, A.; Wang, J.; Cheng, T.; Zhang, Q. Transcriptome Analysis Reveals that Exogenous Melatonin Confers Lilium Disease Resistance to Botrytis elliptica. Front. Genet. 2022, 13, 892674. [Google Scholar] [CrossRef]
- Simons, S.S., Jr. How much is enough? Modulation of dose-response curve for steroid receptor-regulated gene expression by changing concentrations of transcription factor. Curr. Top. Med. Chem. 2006, 6, 271–285. [Google Scholar] [CrossRef]
- Haga, K.; Moritoshi, l. Auxin-Growth Relationships in Maize Coleoptiles and Pea Internodes and Control by Auxin of the Tissue Sensitivity to Auxin. Plant Physiol. 1998, 117, 1473–1486. [Google Scholar] [CrossRef]
- Chandler, P.M.; Robertson, M. Gibberellin dose-response curves and the characterization of dwarf mutants of barley. Plant Physiol. 1999, 120, 623–632. [Google Scholar] [CrossRef]
- Cleland, R. The dosage-response curve for auxin-induced cell elongation: A reevaluation. Planta 1972, 104, 1–9. [Google Scholar] [CrossRef]
- Karcz, W.; Lüthen, H.; Böttger, M. Effect of IAA and 4-Cl-IAA on growth rate in maize coleoptile segments. Acta Physiol. Plant. 1999, 21, 133–139. [Google Scholar] [CrossRef]
- Hernandez-Ruiz, J.; Cano, A.; Arnao, M.B. Melatonin acts as a growth-stimulating compound in some monocot species. J. Pineal Res. 2005, 39, 137–142. [Google Scholar] [CrossRef]
- Agathokleous, E.; Kitao, M.; Calabrese, E.J. New insights into the role of melatonin in plants and animals. Chem. Biol. Interact. 2019, 299, 163–167. [Google Scholar] [CrossRef]
- Park, W.J.; Hertel, R.; Kang, B.G. Enhancement of auxin sensitivity in Ranunculus sceleratus by ethylene: A mechanism to escape from hypoxia under temporary submergence. Environ. Exp. Bot. 2011, 72, 266–271. [Google Scholar] [CrossRef]
- Romanov, G.A.; Getman, I.A.; Schmülling, T. Investigation of early cytokinin effects in a rapid Amaranthus seedling test. Plant Growth Regul. 2000, 32, 337–344. [Google Scholar] [CrossRef]
- Blanco-Ania, D.; Mateman, J.J.; Hýlová, A.; Spíchal, L.; Debie, L.M.; Zwanenburg, B. Hybrid-type strigolactone analogues derived from auxins. Pest. Manag. Sci. 2019, 75, 3113–3121. [Google Scholar] [CrossRef] [PubMed]
- Fendrych, M.; Akhmanova, M.; Merrin, J.; Glanc, M.; Hagihara, S.; Takahashi, K.; Uchida, N.; Torii, K.U.; Friml, J. Rapid and reversible root growth inhibition by TIR1 auxin signalling. Nat. Plants 2018, 4, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Beaudoin, N.; Serizet, C.; Gosti, F.; Giraudat, J. Interactions between Abscisic Acid and Ethylene Signaling Cascades. Plant Cell 2000, 12, 1103–1115. [Google Scholar] [CrossRef] [PubMed]
- Jiroutová, P.; Mikulík, J.; Novák, O.; Strnad, M.; Oklestkova, J. Brassinosteroids Induce Strong, Dose-Dependent Inhibition of Etiolated Pea Seedling Growth Correlated with Ethylene Production. Biomolecules 2019, 9, 849. [Google Scholar] [CrossRef]
- Yamagami, M.; Haga, K.; Napier, R.M.; Iino, M. Two distinct signaling pathways participate in auxin-induced swelling of pea epidermal protoplasts. Plant Physiol. 2004, 134, 735–747. [Google Scholar] [CrossRef]
- Polak, M.; Karcz, W. Some New Methodological and Conceptual Aspects of the “Acid Growth Theory” for the Auxin Action in Maize (Zea mays L.) Coleoptile Segments: Do Acid- and Auxin-Induced Rapid Growth Differ in Their Mechanisms? Int. J. Mol. Sci. 2021, 22, 2317. [Google Scholar] [CrossRef] [PubMed]
- Wigchert, S.C.M.; Kuiper, E.; Boelhouwer, G.J.; Nefkens, G.H.L.; Verkleij, J.A.C.; Zwanenburg, B. Dose−Response of Seeds of the Parasitic Weeds Striga and Orobanche toward the Synthetic Germination Stimulants GR 24 and Nijmegen 1. J. Agric. Food Chem. 1999, 47, 1705–1710. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Fan, B.; Chen, Z. Induction of PR-1 proteins and potentiation of pathogen signals by salicylic acid exhibit the same dose response and structural specificity in plant cell cultures. Mol. Plant-Microbe Interact. 1998, 11, 568–571. [Google Scholar] [CrossRef]
- Li, H.; He, J.; Yang, X.; Li, X.; Luo, D.; Wei, C.; Ma, J.; Zhang, Y.; Yang, J.; Zhang, X. Glutathione-dependent induction of local and systemic defense against oxidative stress by exogenous melatonin in cucumber (Cucumis sativus L.). J. Pineal Res. 2016, 60, 206–216. [Google Scholar] [CrossRef]
- Firn, R.D. Growth substance sensitivity: The need for clearer ideas, precise terms and purposeful experiments. Physiol. Plant. 1986, 67, 267–272. [Google Scholar] [CrossRef]
- Vesper, M.J.; Evans, M.L. Time-dependent Changes in the Auxin Sensitivity of Coleoptile Segments: Apparent Sensory Adaptation 1. Plant Physiol. 1978, 61, 204–208. [Google Scholar] [CrossRef]
- Rai, M.I.; Wang, X.; Thibault, D.M.; Kim, H.J.; Bombyk, M.M.; Binder, B.M.; Shakeel, S.N.; Schaller, G.E. The ARGOS gene family functions in a negative feedback loop to desensitize plants to ethylene. BMC Plant Biol. 2015, 15, 157. [Google Scholar] [CrossRef]
- Gookin, T.E.; Kim, J.; Assmann, S.M. Whole proteome identification of plant candidate G-protein coupled receptors in Arabidopsis, rice, and poplar: Computational prediction and in-vivo protein coupling. Genome Biol. 2008, 9, R120. [Google Scholar] [CrossRef]
- Yang, Q.; Peng, Z.; Ma, W.; Zhang, S.; Hou, S.; Wei, J.; Dong, S.; Yu, X.; Song, Y.; Gao, W.; et al. Melatonin functions in priming of stomatal immunity in Panax notoginseng and Arabidopsis thaliana. Plant Physiol. 2021, 187, 2837–2851. [Google Scholar] [CrossRef] [PubMed]
- Kong, M.; Sheng, T.; Liang, J.; Ali, Q.; Gu, Q.; Wu, H.; Chen, J.; Liu, J.; Gao, X. Melatonin and Its Homologs Induce Immune Responses via Receptors trP47363-trP13076 in Nicotiana benthamiana. Front. Plant Sci. 2021, 12, 691835. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Zuo, T.; Diao, P.; Fu, J.; Fan, Y.; Wang, Y.; Zhao, Q.; Ma, X.; Lu, W.; Li, A.; et al. Melatonin Enhances Seed Germination and Seedling Growth of Medicago sativa Under Salinity via a Putative Melatonin Receptor MsPMTR1. Front. Plant Sci. 2021, 12, 702875. [Google Scholar] [CrossRef]
- Wang, L.F.; Lu, K.K.; Li, T.T.; Zhang, Y.; Guo, J.X.; Song, R.F.; Liu, W.C. Maize PHYTOMELATONIN RECEPTOR1 functions in plant tolerance to osmotic and drought stress. J. Exp. Bot. 2022, 73, 5961–5973. [Google Scholar] [CrossRef]
- Zhang, Y.; Dai, M.; Wu, Z.; Wang, S.; Fan, Y.; Ni, K.; Lu, X.; Liu, X.; Liu, M.; Chen, W.; et al. Melatonin receptor, GhCAND2-D5 motivated responding to NaCl signaling in cotton. Plant Physiol. Biochem. 2023, 203, 108001. [Google Scholar] [CrossRef]
- Li, D.; Wei, J.; Peng, Z.; Ma, W.; Yang, Q.; Song, Z.; Sun, W.; Yang, W.; Yuan, L.; Xu, X.; et al. Daily rhythms of phytomelatonin signaling modulate diurnal stomatal closure via regulating reactive oxygen species dynamics in Arabidopsis. J. Pineal Res. 2020, 68, e12640. [Google Scholar] [CrossRef]
- Wang, L.F.; Li, T.T.; Zhang, Y.; Guo, J.X.; Lu, K.K.; Liu, W.C. CAND2/PMTR1 Is Required for Melatonin-Conferred Osmotic Stress Tolerance in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 4014. [Google Scholar] [CrossRef]
- Bychkov, I.A.; Kudryakova, N.V.; Shugaev, A.G.; Kuznetsov, V.V.; Kusnetsov, V.V. The Melatonin Receptor CAND2/PMTR1 Is Involved in the Regulation of Mitochondrial Gene Expression under Photooxidative Stress. Dokl. Biochem. Biophys. 2022, 502, 15–20. [Google Scholar] [CrossRef]
- Lee, H.Y.; Back, K. The phytomelatonin receptor (PMRT1) Arabidopsis CAND2 is not a bona fide G-protein-coupled melatonin receptor. Melatonin Res. 2020, 3, 10. [Google Scholar] [CrossRef]
- Bai, Y.; Wei, Y.; Yin, H.; Hu, W.; Cheng, X.; Guo, J.; Dong, Y.; Zheng, L.; Xie, H.; Zeng, H.; et al. PP2C1 fine-tunes melatonin biosynthesis and phytomelatonin receptor PMTR1 binding to melatonin in cassava. J. Pineal Res. 2022, 73, e12804. [Google Scholar] [CrossRef] [PubMed]
- Hertel, R. OPINION: Auxin binding protein 1 is a red herring. J. Exp. Bot. 1995, 46, 461–462. [Google Scholar] [CrossRef]
- Venis, M.A. OPINION: Auxin binding protein 1 is a red herring? Oh no it isn’t! J. Exp. Bot. 1995, 46, 463–465. [Google Scholar] [CrossRef]
- Sauer, M.; Kleine-Vehn, J. AUXIN BINDING PROTEIN1: The outsider. Plant Cell 2011, 23, 2033–2043. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.H.; Yang, Z.B. Is ABP1 an auxin receptor yet? Mol. Plant 2011, 4, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Kim, J.Y. Revisiting Apoplastic Auxin Signaling Mediated by AUXIN BINDING PROTEIN 1. Mol. Cells 2015, 38, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Napier, R. The Story of Auxin-Binding Protein 1 (ABP1). Cold Spring Harb. Perspect. Biol. 2021, 13, a039909. [Google Scholar] [CrossRef]
- Hertel, R.; Thomson, K.S.; Russo, V.E. In-vitro auxin binding to particulate cell fractions from corn coleoptiles. Planta 1972, 107, 325–340. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, S.; Sotobayashi, T.; Futai, M.; Fukui, T. Purification and properties of an auxin-binding protein from maize shoot membranes. J. Biochem. 1986, 99, 1513–1524. [Google Scholar] [CrossRef]
- Inohara, N.; Shimomura, S.; Fukui, T.; Futai, M. Auxin-binding protein located in the endoplasmic reticulum of maize shoots: Molecular cloning and complete primary structure. Proc. Natl. Acad. Sci. USA 1989, 86, 3564–3568. [Google Scholar] [CrossRef]
- Dohrmann, U.; Hertel, R.; Kowalik, H. Properties of auxin binding sites in different subcellular fractions from maize coleoptiles. Planta 1978, 140, 97–106. [Google Scholar] [CrossRef]
Figure 1.
Dose–response curves of a phytohormone reveal the sensitivity change reflecting the ligand affinity of the receptor. Arrows indicate the direction of shift of the original dose-response curve (black).
Figure 1.
Dose–response curves of a phytohormone reveal the sensitivity change reflecting the ligand affinity of the receptor. Arrows indicate the direction of shift of the original dose-response curve (black).

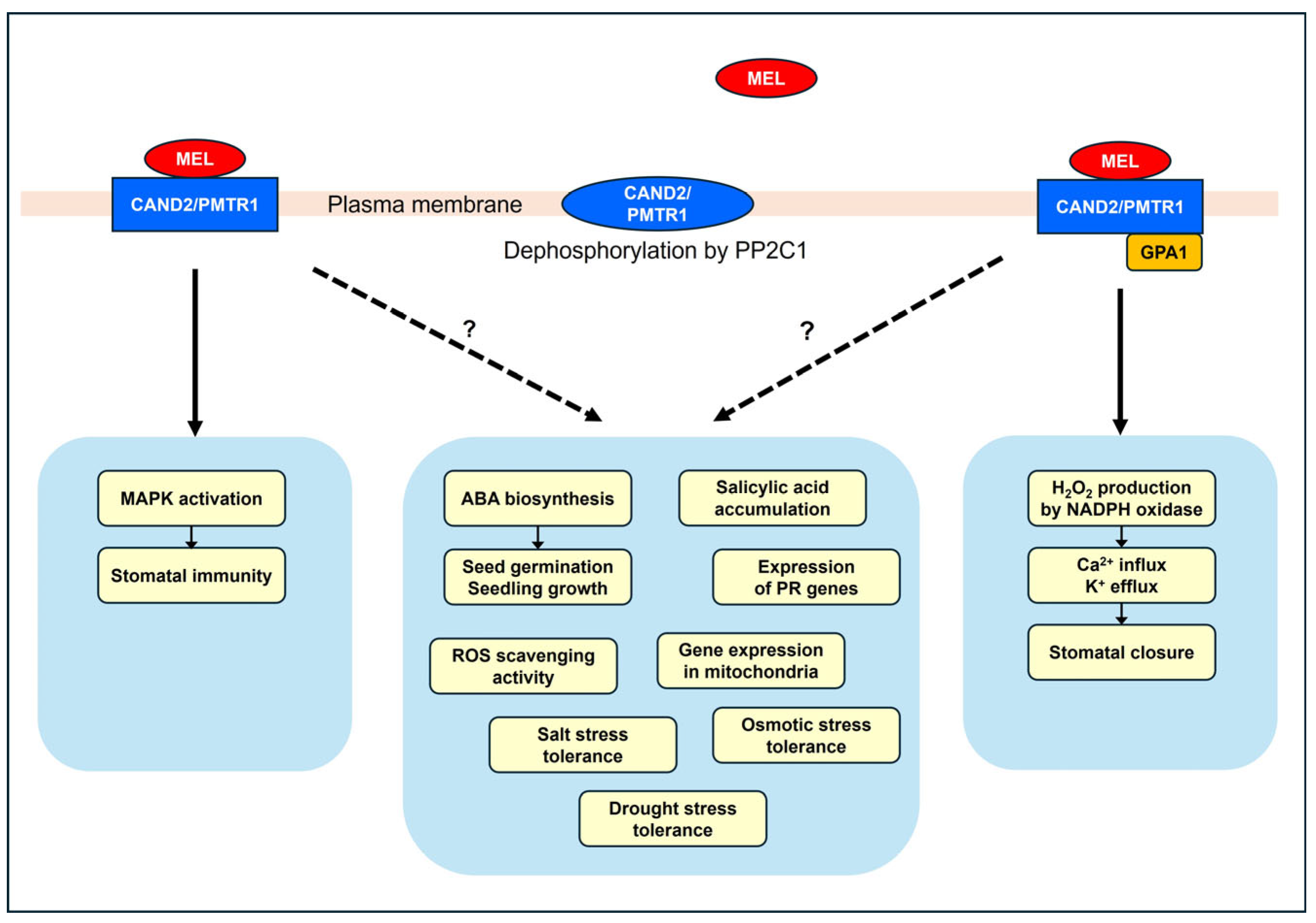
Figure 2.
Melatonin responses regulated by CAND2/PMTR1. MAPK activation occurs independently of GPA1 [119], while stomatal closure is dependent on GPA1 [119]. The mechanisms that regulate ABA biosynthesis [22], salicylic acid accumulation [120], expression of PR genes [120], mitochondrial gene expression [126], ROS scavenging activity [125], and tolerance to salt [123], drought [122], and osmotic [125] stress under the regulation of CAND2/PMTR1 are not yet fully understood. Dephosphorylation of CAND2/PMTR1 abolishes the binding of melatonin to CAND2/PMTR1 [128].
Figure 2.
Melatonin responses regulated by CAND2/PMTR1. MAPK activation occurs independently of GPA1 [119], while stomatal closure is dependent on GPA1 [119]. The mechanisms that regulate ABA biosynthesis [22], salicylic acid accumulation [120], expression of PR genes [120], mitochondrial gene expression [126], ROS scavenging activity [125], and tolerance to salt [123], drought [122], and osmotic [125] stress under the regulation of CAND2/PMTR1 are not yet fully understood. Dephosphorylation of CAND2/PMTR1 abolishes the binding of melatonin to CAND2/PMTR1 [128].

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Park, W.J. Have All of the Phytohormonal Properties of Melatonin Been Verified? Int. J. Mol. Sci. 2024, 25, 3550. https://doi.org/10.3390/ijms25063550
AMA Style
Park WJ. Have All of the Phytohormonal Properties of Melatonin Been Verified? International Journal of Molecular Sciences. 2024; 25(6):3550. https://doi.org/10.3390/ijms25063550
Chicago/Turabian StylePark, Woong June. 2024. "Have All of the Phytohormonal Properties of Melatonin Been Verified?" International Journal of Molecular Sciences 25, no. 6: 3550. https://doi.org/10.3390/ijms25063550
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.