
Genome-Wide Identification of the 14-3-3 Gene Family and Its Involvement in Salt Stress Response through Interaction with NsVP1 in Nitraria sibirica Pall


 and
and
Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification of Ns14-3-3 Gene Family Members and Characterization of Their Proteins
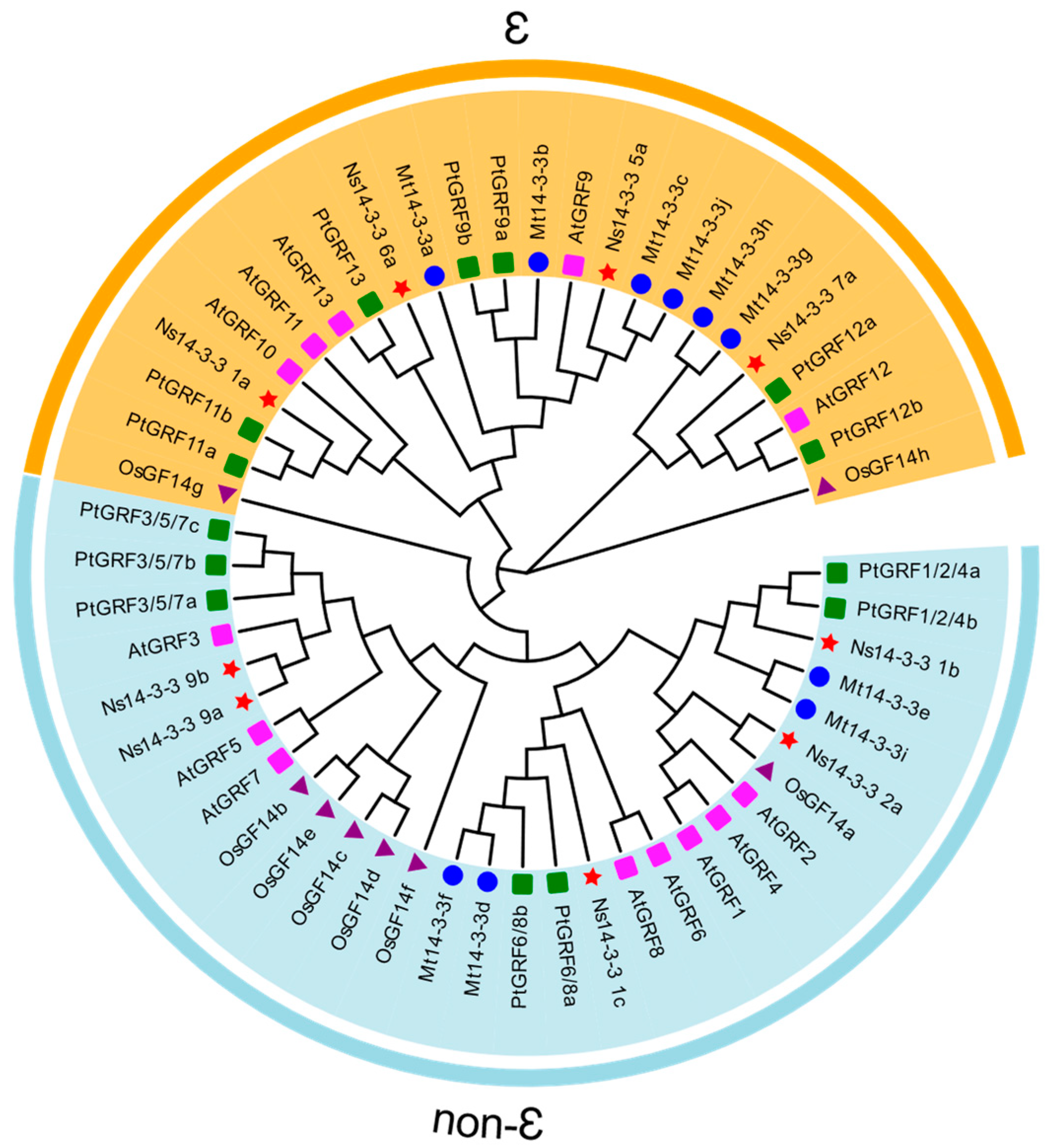
2.2. Phylogenetic Analyses of 14-3-3 Gene Family Members
2.3. The Multiple Sequence Alignment of Ns14-3-3 Proteins
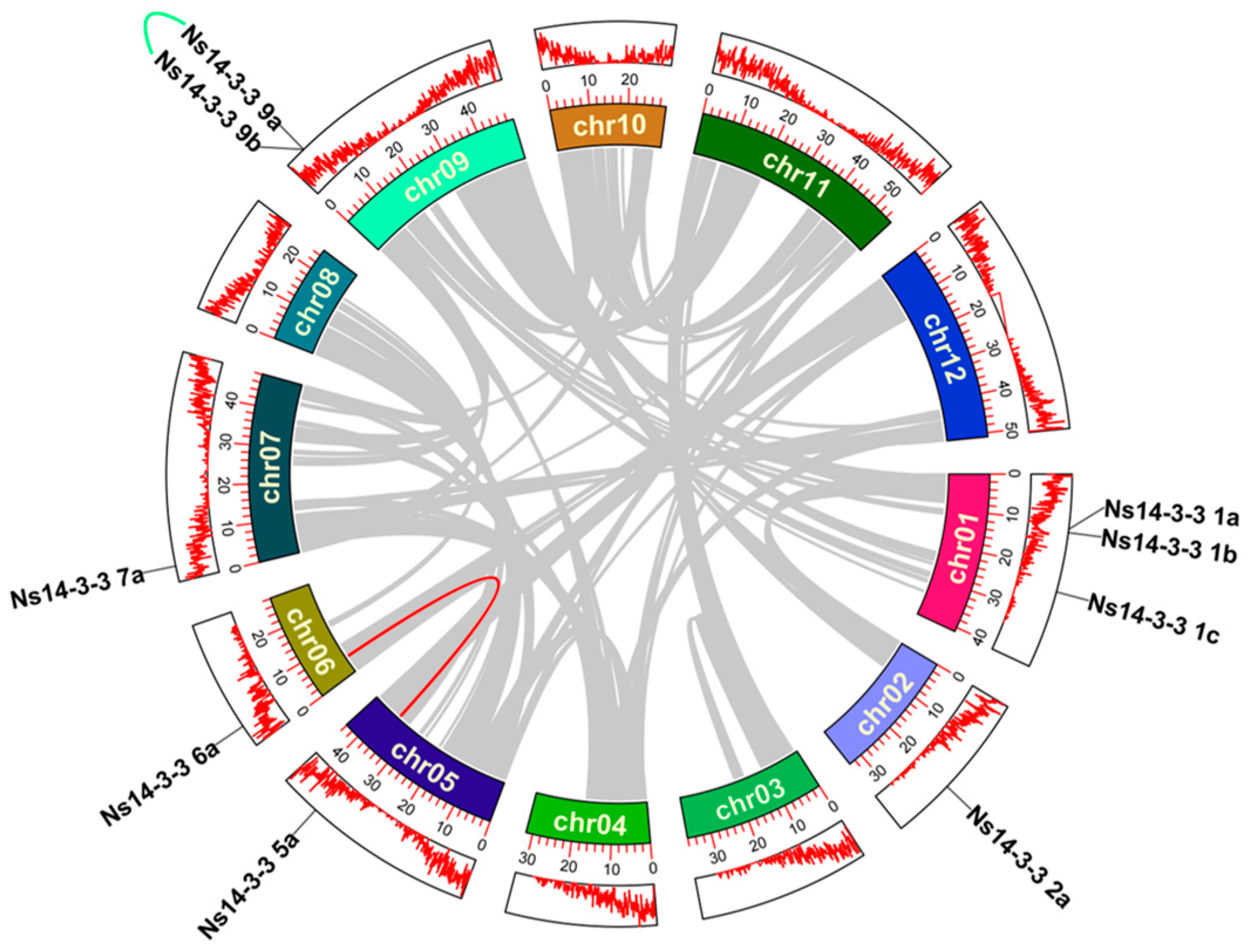
2.4. Chromosome Distribution and Synteny Analysis within Species of 14-3-3 Gene Family Members in N. sibirica
2.5. Gene Structure and Motif Analysis
2.6. Analysis of the Cis-Regulatory Elements of Ns14-3-3 Gene Family Member Promoters
2.7. Transcription Profiling of Ns14-3-3 Genes
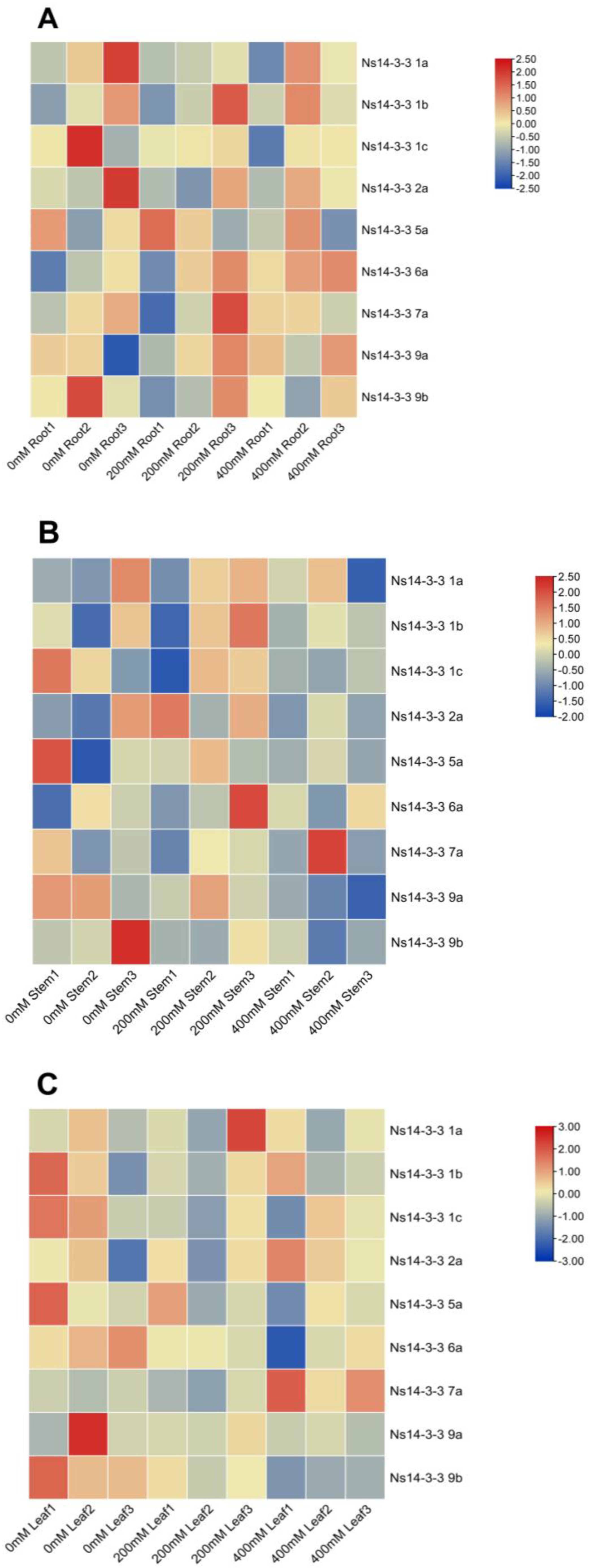
2.8. Expression Profiles of Ns14-3-3 Genes in Various Tissues of N. sibirica
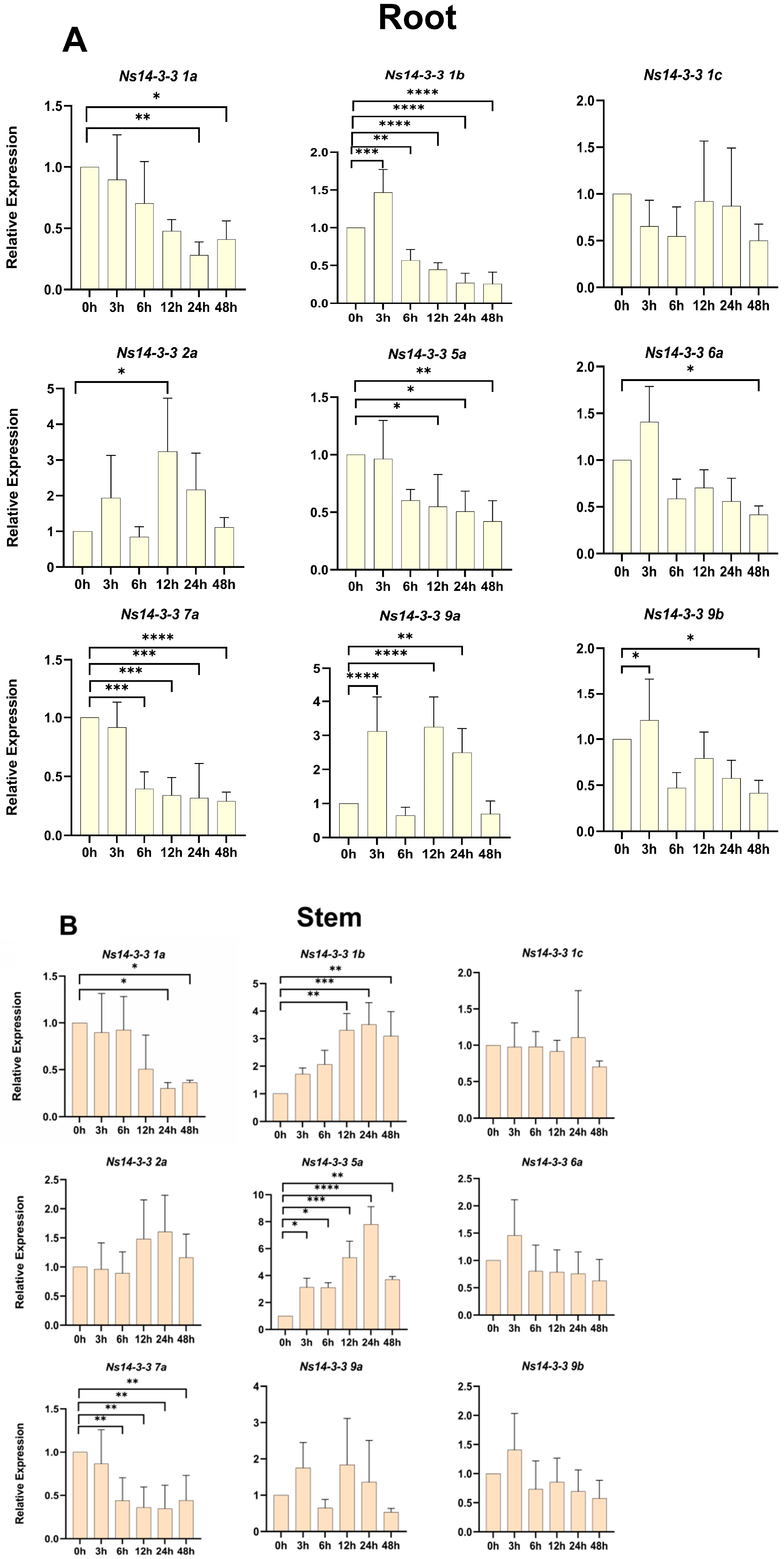
2.9. Expression Patterns of Ns14-3-3s under Salt Stress
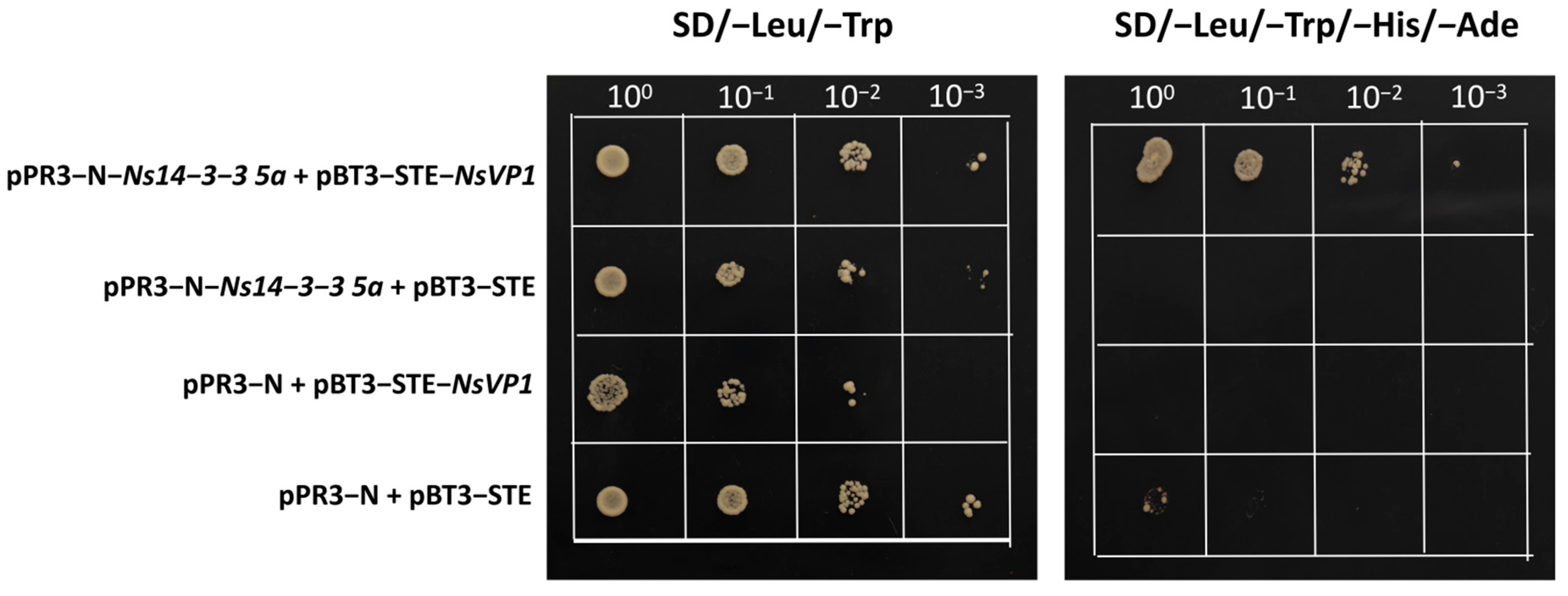
2.10. Validation of Interactions between Ns14-3-3s and NsVP1
3. Discussion
4. Materials and Methods
4.1. Identification and Sequence Analysis of 14-3-3 Gene Family Members in N. sibirica
4.2. Chromosomal Location and Gene Structure Analysis
4.3. Sequence Alignment, Phylogenetic Analysis, and Synteny Analysis of Ns14-3-3s
4.4. Analysis of Cis-Acting Elements in the Promoter Region of Ns14-3-3s
4.5. Transcriptional Profiling of Ns14-3-3s
4.6. Plant Material and Treatments
4.7. Total RNA Extraction, Quantitative Real-Time PCR (qRT-PCR), and Statistical Analysis
4.8. Yeast Two-Hybrid Experiment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, Q.; Zhang, S.; Liu, B. 14-3-3 Proteins: Macro-Regulators with Great Potential for Improving Abiotic Stress Tolerance in Plants. Biochem. Biophys. Res. Commun. 2016, 477, 9–13. [Google Scholar] [CrossRef]
- Zhao, X.; Li, F.; Li, K. The 14-3-3 Proteins: Regulators of Plant Metabolism and Stress Responses. Plant Biol. 2021, 23, 531–539. [Google Scholar] [CrossRef]
- Stevers, L.M.; Sijbesma, E.; Botta, M.; MacKintosh, C.; Obsil, T.; Landrieu, I.; Cau, Y.; Wilson, A.J.; Karawajczyk, A.; Eickhoff, J.; et al. Modulators of 14-3-3 Protein-Protein Interactions. J. Med. Chem. 2018, 61, 3755–3778. [Google Scholar] [CrossRef]
- Aitken, A. 14-3-3 Proteins: A Historic Overview. Semin. Cancer Biol. 2006, 16, 162–172. [Google Scholar] [CrossRef]
- Chevalier, D.; Morris, E.R.; Walker, J.C. 14-3-3 and FHA Domains Mediate Phosphoprotein Interactions. Annu. Rev. Plant Biol. 2009, 60, 67–91. [Google Scholar] [CrossRef]
- Camoni, L.; Visconti, S.; Aducci, P.; Marra, M. 14-3-3 Proteins in Plant Hormone Signaling: Doing Several Things at Once. Front. Plant Sci. 2018, 9, 297. [Google Scholar] [CrossRef]
- Oecking, C.; Jaspert, N. Plant 14-3-3 Proteins Catch up with Their Mammalian Orthologs. Curr. Opin. Plant Biol. 2009, 12, 760–765. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, W.; Yu, H.; Peng, J.; Hu, Z.; Chen, L. The Role of 14-3-3 Proteins in Plant Growth and Response to Abiotic Stress. Plant Cell Rep. 2022, 41, 833–852. [Google Scholar] [CrossRef]
- Yao, Y.; Du, Y.; Jiang, L.; Liu, J.-Y. Molecular Analysis and Expression Patterns of the 14-3-3 Gene Family from Oryza Sativa. J. Biochem. Mol. Biol. 2007, 40, 349–357. [Google Scholar] [CrossRef]
- Wei, X.; Zhang, Z.; Li, Y.; Wang, X.; Shao, S.; Chen, L.; Li, X. Expression Analysis of Two Novel Cotton 14-3-3 Genes in Root Development and in Response to Salt Stress. Prog. Nat. Sci. 2009, 19, 173–178. [Google Scholar] [CrossRef]
- Xu, W.F.; Shi, W.M. Mechanisms of Salt Tolerance in Transgenic Arabidopsis Thaliana Constitutively Overexpressing the Tomato 14-3-3 Protein TFT7. Plant Soil 2007, 301, 17–28. [Google Scholar] [CrossRef]
- He, Y.; Zhang, Y.; Chen, L.; Wu, C.; Luo, Q.; Zhang, F.; Wei, Q.; Li, K.; Chang, J.; Yang, G.; et al. A Member of the 14-3-3 Gene Family in Brachypodium Distachyon, BdGF14d, Confers Salt Tolerance in Transgenic Tobacco Plants. Front. Plant Sci. 2017, 8, 340. [Google Scholar] [CrossRef]
- Gao, G.; Li, Y.; Sun, W.; He, X.; Li, R.; Jin, D.; Qi, X.; Liu, Z.; Bian, S. Functional Roles of Two 14-3-3s in Response to Salt Stress in Common Bean. Acta Physiol. Plant. 2018, 40, 209. [Google Scholar] [CrossRef]
- Zhou, H.; Lin, H.; Chen, S.; Becker, K.; Yang, Y.; Zhao, J.; Kudla, J.; Schumaker, K.S.; Guo, Y. Inhibition of the Arabidopsis Salt Overly Sensitive Pathway by 14-3-3 Proteins. Plant Cell 2014, 26, 1166–1182. [Google Scholar] [CrossRef] [PubMed]
- Latz, A.; Becker, D.; Hekman, M.; Müller, T.; Beyhl, D.; Marten, I.; Eing, C.; Fischer, A.; Dunkel, M.; Bertl, A.; et al. TPK1, a Ca2+-Regulated Arabidopsis Vacuole Two-Pore K+ Channel Is Activated by 14-3-3 Proteins. Plant J. Cell Mol. Biol. 2007, 52, 449–459. [Google Scholar] [CrossRef]
- Latz, A.; Mehlmer, N.; Zapf, S.; Mueller, T.D.; Wurzinger, B.; Pfister, B.; Csaszar, E.; Hedrich, R.; Teige, M.; Becker, D. Salt Stress Triggers Phosphorylation of the Arabidopsis Vacuolar K+ Channel TPK1 by Calcium-Dependent Protein Kinases (CDPKs). Mol. Plant 2013, 6, 1274–1289. [Google Scholar] [CrossRef]
- Fuglsang, A.T.; Guo, Y.; Cuin, T.A.; Qiu, Q.; Song, C.; Kristiansen, K.A.; Bych, K.; Schulz, A.; Shabala, S.; Schumaker, K.S.; et al. Arabidopsis Protein Kinase PKS5 Inhibits the Plasma Membrane H+ -ATPase by Preventing Interaction with 14-3-3 Protein. Plant Cell 2007, 19, 1617–1634. [Google Scholar] [CrossRef]
- Gupta, A.; Shaw, B.P.; Sahu, B.B. Post-Translational Regulation of the Membrane Transporters Contributing to Salt Tolerance in Plants. Funct. Plant Biol. FPB 2021, 48, 1199–1212. [Google Scholar] [CrossRef]
- Rosenquist, M.; Alsterfjord, M.; Larsson, C.; Sommarin, M. Data Mining the Arabidopsis Genome Reveals Fifteen 14-3-3 Genes. Expression Is Demonstrated for Two out of Five Novel Genes. Plant Physiol. 2001, 127, 142–149. [Google Scholar] [CrossRef]
- Yashvardhini, N.; Bhattacharya, S.; Chaudhuri, S.; Sengupta, D.N. Molecular Characterization of the 14-3-3 Gene Family in Rice and Its Expression Studies under Abiotic Stress. Planta 2018, 247, 229–253. [Google Scholar] [CrossRef]
- Jia, C.; Guo, B.; Wang, B.; Li, X.; Yang, T.; Li, N.; Wang, J.; Yu, Q. Genome-Wide Identification and Expression Analysis of the 14-3-3 (TFT) Gene Family in Tomato, and the Role of SlTFT4 in Salt Stress. Plants 2022, 11, 3491. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; He, X.; Huang, X.; Yu, H.; Lu, T.; Xie, X.; Zeng, X.; Zhu, J.; Luo, C. Genome-Wide Identification and Expression Analysis of the 14-3-3 Gene Family in Mango (Mangifera indica L.). Int. J. Mol. Sci. 2022, 23, 1593. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhang, F.; Wu, Z.; Cahaeraduqin, S.; Liu, W.; Yan, Y. Analysis on the Salt Tolerance of Nitraria Sibirica Pall. Based on Pacbio Full-Length Transcriptome Sequencing. Plant Cell Rep. 2023, 42, 1665–1686. [Google Scholar] [CrossRef]
- Li, H.; Tang, X.; Yang, X.; Zhang, H. Comprehensive Transcriptome and Metabolome Profiling Reveal Metabolic Mechanisms of Nitraria Sibirica Pall. to Salt Stress. Sci. Rep. 2021, 11, 12878. [Google Scholar] [CrossRef]
- Tang, X.; Zhang, H.; Shabala, S.; Li, H.; Yang, X.; Zhang, H. Tissue Tolerance Mechanisms Conferring Salinity Tolerance in a Halophytic Perennial Species Nitraria Sibirica Pall. Tree Physiol. 2021, 41, 1264–1277. [Google Scholar] [CrossRef]
- Hsu, Y.-D.; Huang, Y.-F.; Pan, Y.-J.; Huang, L.-K.; Liao, Y.-Y.; Lin, W.-H.; Liu, T.-Y.; Lee, C.-H.; Pan, R.-L. Regulation of H+-Pyrophosphatase by 14-3-3 Proteins from Arabidopsis Thaliana. J. Membr. Biol. 2018, 251, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Martinoia, E.; Maeshima, M.; Neuhaus, H.E. Vacuolar Transporters and Their Essential Role in Plant Metabolism. J. Exp. Bot. 2007, 58, 83–102. [Google Scholar] [CrossRef]
- Mansour, M.M.F. Role of Vacuolar Membrane Transport Systems in Plant Salinity Tolerance. J. Plant Growth Regul. 2023, 42, 1364–1401. [Google Scholar] [CrossRef]
- Tian, F.; Wang, T.; Xie, Y.; Zhang, J.; Hu, J. Genome-Wide Identification, Classification, and Expression Analysis of 14-3-3 Gene Family in Populus. PLoS ONE 2015, 10, e0123225. [Google Scholar] [CrossRef]
- Wang, Y.; Ling, L.; Jiang, Z.; Tan, W.; Liu, Z.; Wu, L.; Zhao, Y.; Xia, S.; Ma, J.; Wang, G.; et al. Genome-Wide Identification and Expression Analysis of the 14-3-3 Gene Family in Soybean (Glycine Max). PeerJ 2019, 7, e7950. [Google Scholar] [CrossRef]
- Qin, C.; Cheng, L.; Shen, J.; Zhang, Y.; Cao, H.; Lu, D.; Shen, C. Genome-Wide Identification and Expression Analysis of the 14-3-3 Family Genes in Medicago truncatula. Front. Plant Sci. 2016, 7, 320. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.; Landherr, L.L.; Frohlich, M.W.; Leebens-Mack, J.; Ma, H.; dePamphilis, C.W. Patterns of Gene Duplication in the Plant SKP1 Gene Family in Angiosperms: Evidence for Multiple Mechanisms of Rapid Gene Birth. Plant J. Cell Mol. Biol. 2007, 50, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, R.; Sethi, S.; Zutshi, I.; Bhalothia, P.; Mehrotra, S. Patterns and Evolution of ACGT Repeat Cis-Element Landscape across Four Plant Genomes. BMC Genom. 2013, 14, 203. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jiang, C.; Kang, L.; Zhang, H.; Song, Y.; Zou, Z.; Zheng, W. Over-Expression of a 14-3-3 Protein From Foxtail Millet Improves Plant Tolerance to Salinity Stress in Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 449. [Google Scholar] [CrossRef]
- Schoonheim, P.J.; Sinnige, M.P.; Casaretto, J.A.; Veiga, H.; Bunney, T.D.; Quatrano, R.S.; de Boer, A.H. 14-3-3 Adaptor Proteins Are Intermediates in ABA Signal Transduction during Barley Seed Germination. Plant J. Cell Mol. Biol. 2007, 49, 289–301. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, H.; Zhou, S.; He, Y.; Luo, Q.; Zhang, F.; Qiu, D.; Feng, J.; Wei, Q.; Chen, L.; et al. Expression of TaGF14b, a 14-3-3 Adaptor Protein Gene from Wheat, Enhances Drought and Salt Tolerance in Transgenic Tobacco. Planta 2018, 248, 117–137. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of Salinity Tolerance in Plants: Physiological, Biochemical, and Molecular Characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef]
- Leng, B.; Wang, X.; Yuan, F.; Zhang, H.; Lu, C.; Chen, M.; Wang, B. Heterologous Expression of the Limonium Bicolor MYB Transcription Factor LbTRY in Arabidopsis thaliana Increases Salt Sensitivity by Modifying Root Hair Development and Osmotic Homeostasis. Plant Sci. Int. J. Exp. Plant Biol. 2021, 302, 110704. [Google Scholar] [CrossRef]
- Wu, W.; Zhang, Q.; Ervin, E.H.; Yang, Z.; Zhang, X. Physiological Mechanism of Enhancing Salt Stress Tolerance of Perennial Ryegrass by 24-Epibrassinolide. Front. Plant Sci. 2017, 8, 1017. [Google Scholar] [CrossRef]
- Qiu, Z.; Guo, J.; Zhu, A.; Zhang, L.; Zhang, M. Exogenous Jasmonic Acid Can Enhance Tolerance of Wheat Seedlings to Salt Stress. Ecotoxicol. Environ. Saf. 2014, 104, 202–208. [Google Scholar] [CrossRef]
- Shi, H.; Zhang, Y. Pear 14-3-3a Gene (Pp14-3-3a) Is Regulated during Fruit Ripening and Senescense, and Involved in Response to Salicylic Acid and Ethylene Signalling. J. Genet. 2014, 93, 747–753. [Google Scholar] [CrossRef]
- Garg, N.; Bharti, A. Salicylic Acid Improves Arbuscular Mycorrhizal Symbiosis, and Chickpea Growth and Yield by Modulating Carbohydrate Metabolism under Salt Stress. Mycorrhiza 2018, 28, 727–746. [Google Scholar] [CrossRef] [PubMed]
- Ishida, S.; Fukazawa, J.; Yuasa, T.; Takahashi, Y. Involvement of 14-3-3 Signaling Protein Binding in the Functional Regulation of the Transcriptional Activator Repression of Shoot Growth by Gibberellins. Plant Cell 2004, 16, 2641–2651. [Google Scholar] [CrossRef]
- Maggio, A.; Barbieri, G.; Raimondi, G.; De Pascale, S. Contrasting Effects of GA3 Treatments on Tomato Plants Exposed to Increasing Salinity. J. Plant Growth Regul. 2010, 29, 63–72. [Google Scholar] [CrossRef]
- Mao, J.; Niu, C.; Li, K.; Mobeen Tahir, M.; Khan, A.; Wang, H.; Li, S.; Liang, Y.; Li, G.; Yang, Z.; et al. Exogenous 6-Benzyladenine Application Affects Root Morphology by Altering Hormone Status and Gene Expression of Developing Lateral Roots in Malus hupehensis. Plant Biol. 2020, 22, 1150–1159. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.-K.; Bohnert, H.J. Plant Cellular and Molecular Responses to High Salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef]
- Blumwald, E. Sodium Transport and Salt Tolerance in Plants. Curr. Opin. Cell Biol. 2000, 12, 431–434. [Google Scholar] [CrossRef]
- Shabala, S.; Pottosin, I. Regulation of Potassium Transport in Plants under Hostile Conditions: Implications for Abiotic and Biotic Stress Tolerance. Physiol. Plant. 2014, 151, 257–279. [Google Scholar] [CrossRef]
- Lin, S.-M.; Tsai, J.-Y.; Hsiao, C.-D.; Huang, Y.-T.; Chiu, C.-L.; Liu, M.-H.; Tung, J.-Y.; Liu, T.-H.; Pan, R.-L.; Sun, Y.-J. Crystal Structure of a Membrane-Embedded H+-Translocating Pyrophosphatase. Nature 2012, 484, 399–403. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.-K.; Shabala, S. Mechanisms of Plant Responses and Adaptation to Soil Salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef] [PubMed]
- Poole, R.L. The TAIR Database. Methods Mol. Biol. Clifton NJ 2007, 406, 179–212. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Hu, A.; Yang, X.; Zhu, J.; Wang, X.; Liu, J.; Wang, J.; Wu, H.; Zhang, H.; Zhang, H. Selection and Validation of Appropriate Reference Genes for RT-qPCR Analysis of Nitraria Sibirica under Various Abiotic Stresses. BMC Plant Biol. 2022, 22, 592. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Iyer, K.; Bürkle, L.; Auerbach, D.; Thaminy, S.; Dinkel, M.; Engels, K.; Stagljar, I. Utilizing the Split-Ubiquitin Membrane Yeast Two-Hybrid System to Identify Protein-Protein Interactions of Integral Membrane Proteins. Sci. STKE Signal Transduct. Knowl. Environ. 2005, 2005, pl3. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | ID | Chr | aa | pI | II | MW (Da) | GRAVY | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|
| Ns14−3−3 1a | Nisib01aG0106800.1 | Chr01 | 253 | 4.96 | 51.41 | 28,887.41 | −0.626 | plas |
| Ns14−3−3 1b | Nisib01aG0110100.1 | Chr01 | 265 | 4.73 | 44.81 | 29,929.55 | −0.563 | plas |
| Ns14−3−3 1c | Nisib01aG0151400.1 | Chr01 | 252 | 4.76 | 38.01 | 28,539.45 | −0.287 | chlo |
| Ns14−3−3 2a | Nisib02aG0114500.1 | Chr02 | 264 | 4.72 | 49.62 | 29,789.47 | −0.492 | chlo |
| Ns14−3−3 5a | Nisib05aG0198000.1 | Chr05 | 261 | 4.78 | 38.18 | 29,371.89 | −0.55 | nucl |
| Ns14−3−3 6a | Nisib06aG0046300.1 | Chr06 | 257 | 4.73 | 44.24 | 29,290.77 | −0.615 | nucl |
| Ns14−3−3 7a | Nisib07aG0034600.1 | Chr07 | 262 | 4.90 | 48.29 | 29,860.49 | −0.702 | nucl |
| Ns14−3−3 9a | Nisib09aG0041400.2 | Chr09 | 249 | 4.97 | 45.09 | 28,234.04 | −0.310 | cyto |
| Ns14−3−3 9b | Nisib09aG0041400.1 | Chr09 | 261 | 4.78 | 49.08 | 29,479.14 | −0.47 | nucl |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, X.; Duan, R.; Zhang, H.; Zhu, J.; Wu, H.; Zhang, H.; Yang, X. Genome-Wide Identification of the 14-3-3 Gene Family and Its Involvement in Salt Stress Response through Interaction with NsVP1 in Nitraria sibirica Pall. Int. J. Mol. Sci. 2024, 25, 3432. https://doi.org/10.3390/ijms25063432
Wan X, Duan R, Zhang H, Zhu J, Wu H, Zhang H, Yang X. Genome-Wide Identification of the 14-3-3 Gene Family and Its Involvement in Salt Stress Response through Interaction with NsVP1 in Nitraria sibirica Pall. International Journal of Molecular Sciences. 2024; 25(6):3432. https://doi.org/10.3390/ijms25063432
Chicago/Turabian StyleWan, Xihong, Rongfeng Duan, Huaxin Zhang, Jianfeng Zhu, Haiwen Wu, Huilong Zhang, and Xiuyan Yang. 2024. "Genome-Wide Identification of the 14-3-3 Gene Family and Its Involvement in Salt Stress Response through Interaction with NsVP1 in Nitraria sibirica Pall" International Journal of Molecular Sciences 25, no. 6: 3432. https://doi.org/10.3390/ijms25063432