
Domestication Gene Mlx and Its Partner Mondo Are Involved in Controlling the Larval Body Size and Cocoon Shell Weight of Bombyx mori

Abstract
:1. Introduction
2. Results
2.1. Domestication Selection Analysis of a TE Deletion in the Upstream Regulatory Region of Mlx Gene in Domesticated Silkworm
2.2. Retrotransposon Bm1 Is Involved in Modulating the Expression of Mlx
2.3. Mondo-Mlx Regulates the Expression of Genes Related to Trehalose Transport and Glycolysis
2.4. Direct Regulation of Lipogenic Genes by Mondo-Mlx
2.5. Mondo-Mlx Regulates Directly the Expression of Glutamine Synthase 2
2.6. Mondo-Mlx Controls Cabut, Cycle and FAMeT5 Expression
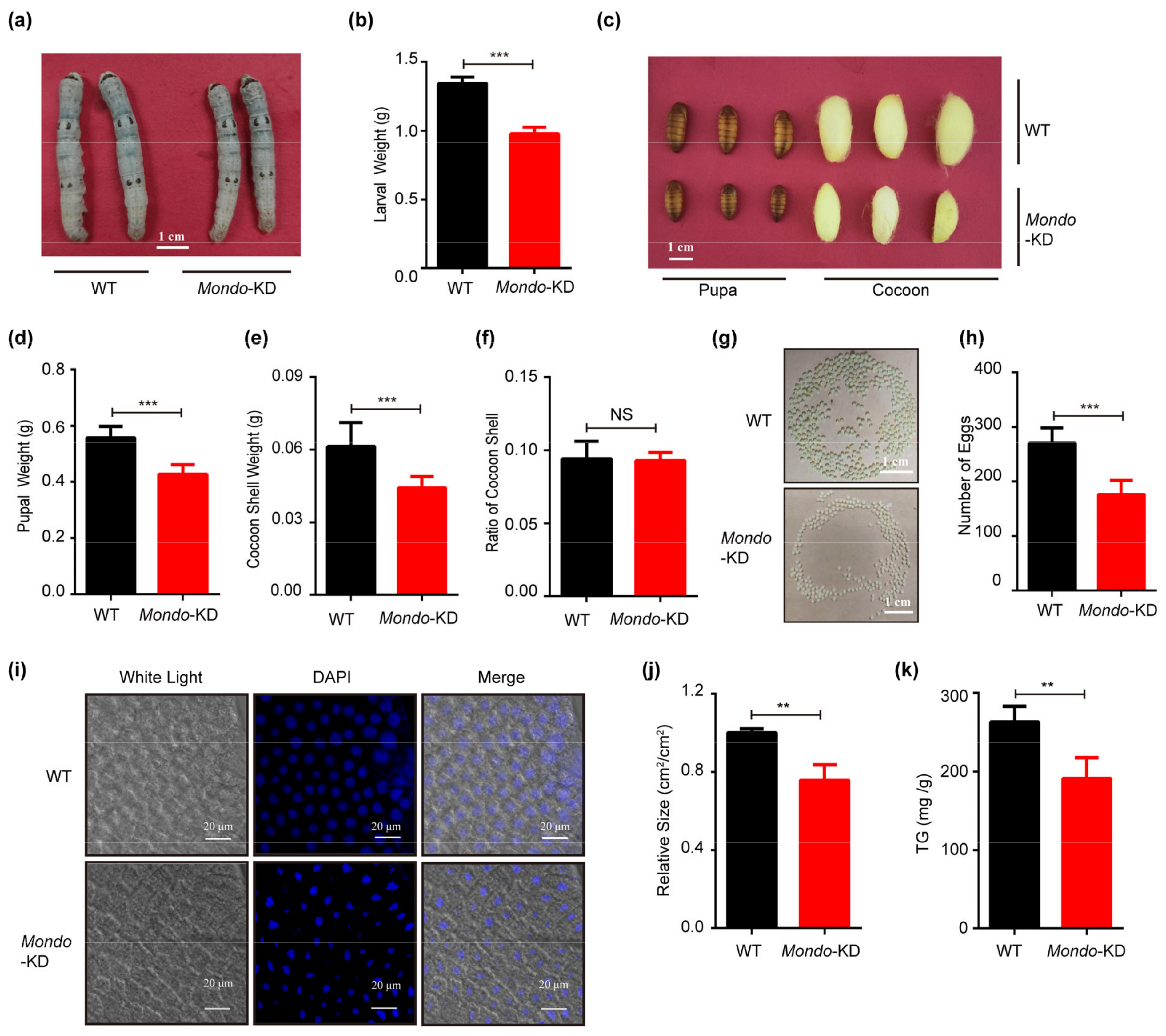
2.7. Manipulating the Expression of Mondo-Mlx Alters Body Size and Cocoon Weight
3. Discussion
4. Materials and Methods
4.1. Statistics of Sequence Polymorphism and Neutrality Tests
4.2. Cells Culture and Transfection
4.3. Total RNA Extraction and Quantitative Real-Time PCR (qPCR)
4.4. Dual Luciferase Reporter Assay
4.5. Electrophoretic Mobility Shift Assay (EMSA)
4.6. ChIP-PCR
4.7. RNA-Seq
4.8. Transgenic Strains Generated
4.9. Insect Rearing and Statistics of Economic Characteristics
4.10. Measure of Triglyceride Content
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hubbell, S. Shrinking the Cat: Genetic Engineering Before We Knew About Genes; Houghton Mifflin: Boston, MA, USA, 2001; p. 175. [Google Scholar]
- Goldsmith, M.R.; Shimada, T.; Abe, H. The genetics and genomics of the silkworm, Bombyx mori. Annu. Rev. Entomol. 2005, 50, 71–100. [Google Scholar] [CrossRef]
- Underhill, A.P. Current issues in Chinese neolithic archaeology. J. World Prehistory 1997, 11, 103–160. [Google Scholar] [CrossRef]
- Xiang, Z.H.; Huang, J.T.; Xia, J.G.; Lu, C. Biology of Sericulture; China Forestry Publishing House: Beijing, China, 2005. [Google Scholar]
- Xiang, H.; Liu, X.; Li, M.; Zhu, Y.; Wang, L.; Cui, Y.; Liu, L.; Fang, G.; Qian, H.; Xu, A.; et al. The evolutionary road from wild moth to domestic silkworm. Nat. Ecol. Evol. 2018, 2, 1268–1279. [Google Scholar] [CrossRef] [PubMed]
- Mc, C.B. Maize genetics. In Year Book; Carnegie Institution of Washington: Washington, DC, USA, 1946; Volume 45, pp. 176–186. [Google Scholar]
- Mc, C.B. The origin and behavior of mutable loci in maize. Proc. Natl. Acad. Sci. USA 1950, 36, 344–355. [Google Scholar]
- McClintock, B. Intranuclear systems controlling gene action and mutation. Brookhaven Symp. Biol. 1956, 8, 58–74. [Google Scholar]
- Burns, K.H.; Boeke, J.D. Human transposon tectonics. Cell 2012, 149, 740–752. [Google Scholar] [CrossRef] [PubMed]
- Studer, A.; Zhao, Q.; Ross-Ibarra, J.; Doebley, J. Identification of a functional transposon insertion in the maize domestication gene tb1. Nat. Genet. 2011, 43, 1160–1163. [Google Scholar] [CrossRef]
- Butelli, E.; Licciardello, C.; Zhang, Y.; Liu, J.; Mackay, S.; Bailey, P.; Reforgiato-Recupero, G.; Martin, C. Retrotransposons control fruit-specific, cold-dependent accumulation of anthocyanins in blood oranges. Plant Cell 2012, 24, 1242–1255. [Google Scholar] [CrossRef]
- Engels, W.R. The P family of transposable elements in Drosophila. Annu. Rev. Genet. 1983, 17, 315–344. [Google Scholar] [CrossRef]
- Lewis, A.P.; Brookfield, J.F.Y. Movement of Drosophila melanogaster transposable elements other than P elements in a P-M hybrid dysgenic cross. Mol. Gen. Genet. 1987, 208, 506–510. [Google Scholar] [CrossRef]
- Klutsch, C.F.; de Caprona, M.D. The IGF1 small dog haplotype is derived from Middle Eastern grey wolves: A closer look at statistics, sampling, and the alleged Middle Eastern origin of small dogs. BMC Biol. 2010, 8, 119. [Google Scholar] [CrossRef] [PubMed]
- Clark, L.A.; Wahl, J.M.; Rees, C.A.; Murphy, K.E. Retrotransposon insertion in SILV is responsible for merle patterning of the domestic dog. Proc. Natl. Acad. Sci. USA 2006, 103, 1376–1381. [Google Scholar] [CrossRef] [PubMed]
- Dreger, D.L.; Schmutz, S.M. A SINE insertion causes the black-and-tan and saddle tan phenotypes in domestic dogs. J. Hered. 2011, 102 (Suppl. S1), S11–S18. [Google Scholar] [CrossRef] [PubMed]
- David, V.A.; Menotti-Raymond, M.; Wallace, A.C.; Roelke, M.; Kehler, J.; Leighty, R.; Eizirik, E.; Hannah, S.S.; Nelson, G.; Schaffer, A.A.; et al. Endogenous retrovirus insertion in the KIT oncogene determines white and white spotting in domestic cats. G3 2014, 4, 1881–1891. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Shen, Y.H.; Han, M.J.; Cao, Y.F.; Zhang, Z. An adaptive transposable element insertion in the regulatory region of the EO gene in the domesticated silkworm, Bombyx mori. Mol. Biol. Evol. 2014, 31, 3302–3313. [Google Scholar] [CrossRef] [PubMed]
- Foufelle, F.; Gouhot, B.; Pegorier, J.P.; Perdereau, D.; Girard, J.; Ferre, P. Glucose Stimulation of Lipogenic Enzyme Gene-Expression in Cultured White Adipose-Tissue-a Role for Glucose-6-Phosphate. J. Biol. Chem. 1992, 267, 20543–20546. [Google Scholar] [CrossRef] [PubMed]
- Dentin, R.; Tomas-Cobos, L.; Foufelle, F.; Leopold, J.; Girard, J.; Postic, C.; Ferre, P. Glucose 6-phosphate, rather than xylulose 5-phosphate, is required for the activation of ChREBP in response to glucose in the liver. J. Hepatol. 2012, 56, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Li, M.V.; Chen, W.; Harmancey, R.N.; Nuotio-Antar, A.M.; Imamura, M.; Saha, P.; Taegtmeyer, H.; Chan, L. Glucose-6-phosphate mediates activation of the carbohydrate responsive binding protein (ChREBP). Biochem. Biophys. Res. Commun. 2010, 395, 395–400. [Google Scholar] [CrossRef]
- Stoltzman, C.A.; Kaadige, M.R.; Peterson, C.W.; Ayer, D.E. MondoA senses non-glucose sugars: Regulation of thioredoxin-interacting protein (TXNIP) and the hexose transport curb. J. Biol. Chem. 2011, 286, 38027–38034. [Google Scholar] [CrossRef]
- Billin, A.N.; Eilers, A.L.; Queva, C.; Ayer, D.E. Mlx, a novel Max-like BHLHZip protein that interacts with the Max network of transcription factors. J. Biol. Chem. 1999, 274, 36344–36350. [Google Scholar] [CrossRef]
- Billin, A.N.; Eilers, A.L.; Coulter, K.L.; Logan, J.S.; Ayer, D.E. MondoA, a novel basic helix-loop-helix-leucine zipper transcriptional activator that constitutes a positive branch of a max-like network. Mol. Cell. Biol. 2000, 20, 8845–8854. [Google Scholar] [CrossRef] [PubMed]
- Cairo, S.; Merla, G.; Urbinati, F.; Ballabio, A.; Reymond, A. WBSCR14, a gene mapping to the Williams--Beuren syndrome deleted region, is a new member of the Mlx transcription factor network. Hum. Mol. Genet. 2001, 10, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, H.; Takenoshita, M.; Sakurai, M.; Bruick, R.K.; Henzel, W.J.; Shillinglaw, W.; Arnot, D.; Uyeda, K. A glucose-responsive transcription factor that regulates carbohydrate metabolism in the liver. Proc. Natl. Acad. Sci. USA 2001, 98, 9116–9121. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wollheim, C.B. ChREBP rather than USF2 regulates glucose stimulation of endogenous L-pyruvate kinase expression in insulin-secreting cells. J. Biol. Chem. 2002, 277, 32746–32752. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Jiang, T.; Wang, Z.; Levi, M.; Li, J. Modulation of carbohydrate response element-binding protein gene expression in 3T3-L1 adipocytes and rat adipose tissue. American journal of physiology. Endocrinol. Metab. 2004, 287, E424–E430. [Google Scholar]
- Ma, L.; Tsatsos, N.G.; Towle, H.C. Direct role of ChREBP.Mlx in regulating hepatic glucose-responsive genes. J. Biol. Chem. 2005, 280, 12019–12027. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Robinson, L.N.; Towle, H.C. ChREBP*Mlx is the principal mediator of glucose-induced gene expression in the liver. J. Biol. Chem. 2006, 281, 28721–28730. [Google Scholar] [CrossRef]
- Havula, E.; Teesalu, M.; Hyotylainen, T.; Seppala, H.; Hasygar, K.; Auvinen, P.; Oresic, M.; Sandmann, T.; Hietakangas, V. Mondo/ChREBP-Mlx-regulated transcriptional network is essential for dietary sugar tolerance in Drosophila. PLoS Genet. 2013, 9, e1003438. [Google Scholar] [CrossRef]
- Ishii, S.; Iizuka, K.; Miller, B.C.; Uyeda, K. Carbohydrate response element binding protein directly promotes lipogenic enzyme gene transcription. Proc. Natl. Acad. Sci. USA 2004, 101, 15597–15602. [Google Scholar] [CrossRef]
- Mattila, J.; Havula, E.; Suominen, E.; Teesalu, M.; Surakka, I.; Hynynen, R.; Kilpinen, H.; Vaananen, J.; Hovatta, I.; Kakela, R.; et al. Mondo-Mlx Mediates Organismal Sugar Sensing through the Gli-Similar Transcription Factor Sugarbabe. Cell Rep. 2015, 13, 350–364. [Google Scholar] [CrossRef]
- Sans, C.L.; Satterwhite, D.J.; Stoltzman, C.A.; Breen, K.T.; Ayer, D.E. MondoA-Mlx heterodimers are candidate sensors of cellular energy status: Mitochondrial localization and direct regulation of glycolysis. Mol. Cell. Biol. 2006, 26, 4863–4871. [Google Scholar] [CrossRef] [PubMed]
- Stoltzman, C.A.; Peterson, C.W.; Breen, K.T.; Muoio, D.M.; Billin, A.N.; Ayer, D.E. Glucose sensing by MondoA:Mlx complexes: A role for hexokinases and direct regulation of thioredoxin-interacting protein expression. Proc. Natl. Acad. Sci. USA 2008, 105, 6912–6917. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.S.; Kim, D.; Lee, Y.S.; Kim, H.J.; Han, J.Y.; Im, S.S.; Chong, H.K.; Kwon, J.K.; Cho, Y.H.; Kim, W.K.; et al. Integrated expression profiling and genome-wide analysis of ChREBP targets reveals the dual role for ChREBP in glucose-regulated gene expression. PLoS ONE 2011, 6, e22544. [Google Scholar] [CrossRef] [PubMed]
- Musselman, L.P.; Fink, J.L.; Ramachandran, P.V.; Patterson, B.W.; Okunade, A.L.; Maier, E.; Brent, M.R.; Turk, J.; Baranski, T.J. Role of fat body lipogenesis in protection against the effects of caloric overload in Drosophila. J. Biol. Chem. 2013, 288, 8028–8042. [Google Scholar] [CrossRef] [PubMed]
- Bartok, O.; Teesalu, M.; Ashwall-Fluss, R.; Pandey, V.; Hanan, M.; Rovenko, B.M.; Poukkula, M.; Havula, E.; Moussaieff, A.; Vodala, S.; et al. The transcription factor Cabut coordinates energy metabolism and the circadian clock in response to sugar sensing. EMBO J. 2015, 34, 1538–1553. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.W.; Llop, J.R.; Farrell, S.F.; Yuan, J.; Stolzenburg, L.R.; Samuelson, A.V. The Caenorhabditis elegans Myc-Mondo/Mad complexes integrate diverse longevity signals. PLoS Genet. 2014, 10, e1004278. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, K.; Bruick, R.K.; Liang, G.; Horton, J.D.; Uyeda, K. Deficiency of carbohydrate response element-binding protein (ChREBP) reduces lipogenesis as well as glycolysis. Proc. Natl. Acad. Sci. USA 2004, 101, 7281–7286. [Google Scholar] [CrossRef]
- Herman, M.A.; Peroni, O.D.; Villoria, J.; Schon, M.R.; Abumrad, N.A.; Bluher, M.; Klein, S.; Kahn, B.B. A novel ChREBP isoform in adipose tissue regulates systemic glucose metabolism. Nature 2012, 484, 333–338. [Google Scholar] [CrossRef]
- Kathiresan, S.; Melander, O.; Guiducci, C.; Surti, A.; Burtt, N.P.; Rieder, M.J.; Cooper, G.M.; Roos, C.; Voight, B.F.; Havulinna, A.S.; et al. Six new loci associated with blood low-density lipoprotein cholesterol, high-density lipoprotein cholesterol or triglycerides in humans. Nat. Genet. 2008, 40, 189–197. [Google Scholar] [CrossRef]
- Kooner, J.S.; Chambers, J.C.; Aguilar-Salinas, C.A.; Hinds, D.A.; Hyde, C.L.; Warnes, G.R.; Gomez Perez, F.J.; Frazer, K.A.; Elliott, P.; Scott, J.; et al. Genome-wide scan identifies variation in MLXIPL associated with plasma triglycerides. Nat. Genet. 2008, 40, 149–151. [Google Scholar] [CrossRef]
- Shi, J.H.; Lu, J.Y.; Chen, H.Y.; Wei, C.C.; Xu, X.; Li, H.; Bai, Q.; Xia, F.Z.; Lam, S.M.; Zhang, H.; et al. Liver ChREBP Protects Against Fructose-Induced Glycogenic Hepatotoxicity by Regulating L-Type Pyruvate Kinase. Diabetes 2020, 69, 591–602. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, S.; Ospina, E.; Shabandri, O.; Lank, D.; Akakpo, J.Y.; Zhao, Z.; Yang, M.; Wu, J.; Jaeschke, H.; et al. Fructose Protects Against Acetaminophen-Induced Hepatotoxicity Mainly by Activating the Carbohydrate-Response Element-Binding Protein alpha-Fibroblast Growth Factor 21 Axis in Mice. Hepatol. Commun. 2021, 5, 992–1008. [Google Scholar] [CrossRef] [PubMed]
- Carroll, P.A.; Diolaiti, D.; McFerrin, L.; Gu, H.; Djukovic, D.; Du, J.; Cheng, P.F.; Anderson, S.; Ulrich, M.; Hurley, J.B.; et al. Deregulated Myc requires MondoA/Mlx for metabolic reprogramming and tumorigenesis. Cancer Cell 2015, 27, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Zhao, F.; Mancuso, A.; Gruber, J.J.; Thompson, C.B. The glucose-responsive transcription factor ChREBP contributes to glucose-dependent anabolic synthesis and cell proliferation. Proc. Natl. Acad. Sci. USA 2009, 106, 21660–21665. [Google Scholar] [CrossRef] [PubMed]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, J.Y.; Yan, X.D.; Su, K.; Cao, D.M.; Zhang, W.P. Fructose intolerance induced by intestinal ChREBP deficiency in mice. Chin. J. Pathophysiol. 2021, 37, 586–592. [Google Scholar]
- Xia, Q.; Guo, Y.; Zhang, Z.; Li, D.; Xuan, Z.; Li, Z.; Dai, F.; Li, Y.; Cheng, D.; Li, R.; et al. Complete resequencing of 40 genomes reveals domestication events and genes in silkworm (Bombyx). Science 2009, 326, 433–436. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Han, M.J.; Lu, K.; Tai, S.; Liang, S.; Liu, Y.; Hu, H.; Shen, J.; Long, A.; Zhan, C.; et al. High-resolution silkworm pan-genome provides genetic insights into artificial selection and ecological adaptation. Nat. Commun. 2022, 13, 5619. [Google Scholar] [CrossRef]
- McConnell, B.B.; Yang, V.W. Mammalian Kruppel-like factors in health and diseases. Physiol. Rev. 2010, 90, 1337–1381. [Google Scholar] [CrossRef]
- Pearson, R.; Fleetwood, J.; Eaton, S.; Crossley, M.; Bao, S. Kruppel-like transcription factors: A functional family. Int. J. Biochem. Cell Biol. 2008, 40, 1996–2001. [Google Scholar] [CrossRef]
- Suske, G.; Bruford, E.; Philipsen, S. Mammalian SP/KLF transcription factors: Bring in the family. Genomics 2005, 85, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Tetreault, M.P.; Yang, Y.; Katz, J.P. Kruppel-like factors in cancer. Nature reviews. Cancer 2013, 13, 701–713. [Google Scholar] [PubMed]
- Hirota, T.; Kon, N.; Itagaki, T.; Hoshina, N.; Okano, T.; Fukada, Y. Transcriptional repressor TIEG1 regulates Bmal1 gene through GC box and controls circadian clockwork. Genes Cells Devoted Mol. Cell. Mech. 2010, 15, 111–121. [Google Scholar] [CrossRef]
- Lomberk, G.; Urrutia, R. The family feud: Turning off Sp1 by Sp1-like KLF proteins. Biochem. J. 2005, 392 Pt 1, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Shields, J.M.; Yang, V.W. Identification of the DNA sequence that interacts with the gut-enriched Kruppel-like factor. Nucleic Acids Res. 1998, 26, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Spittau, B.; Wang, Z.; Boinska, D.; Krieglstein, K. Functional domains of the TGF-beta-inducible transcription factor Tieg3 and detection of two putative nuclear localization signals within the zinc finger DNA-binding domain. J. Cell. Biochem. 2007, 101, 712–722. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.L.; Grau, D.J.; DeVido, S.K.; Kassis, J.A. An Sp1/KLF binding site is important for the activity of a Polycomb group response element from the Drosophila engrailed gene. Nucleic Acids Res. 2005, 33, 5181–5189. [Google Scholar] [CrossRef] [PubMed]
- Belacortu, Y.; Weiss, R.; Kadener, S.; Paricio, N. Transcriptional activity and nuclear localization of Cabut, the Drosophila ortholog of vertebrate TGF-beta-inducible early-response gene (TIEG) proteins. PLoS ONE 2012, 7, e32004. [Google Scholar] [CrossRef]
- Grunstein, M. Histone acetylation in chromatin structure and transcription. Nature 1997, 389, 349–352. [Google Scholar] [CrossRef]
- Eberharter, A.; Becker, P.B. Histone acetylation: A switch between repressive and permissive chromatin. Second in review series on chromatin dynamics. EMBO Rep. 2002, 3, 224–229. [Google Scholar] [CrossRef]
- Vidal, M.; Gaber, R.F. RPD3 encodes a second factor required to achieve maximum positive and negative transcriptional states in Saccharomyces cerevisiae. Mol. Cell. Biol. 1991, 11, 6317–6327. [Google Scholar] [PubMed]
- Yang, W.M.; Inouye, C.; Zeng, Y.; Bearss, D.; Seto, E. Transcriptional repression by YY1 is mediated by interaction with a mammalian homolog of the yeast global regulator RPD3. Proc. Natl. Acad. Sci. USA 1996, 93, 12845–12850. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.M.; Yao, Y.L.; Sun, J.M.; Davie, J.R.; Seto, E. Isolation and characterization of cDNAs corresponding to an additional member of the human histone deacetylase gene family. J. Biol. Chem. 1997, 272, 28001–28007. [Google Scholar] [CrossRef] [PubMed]
- Kao, H.Y.; Downes, M.; Ordentlich, P.; Evans, R.M. Isolation of a novel histone deacetylase reveals that class I and class II deacetylases promote SMRT-mediated repression. Genes Dev. 2000, 14, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Kao, H.Y.; Lee, C.H.; Komarov, A.; Han, C.C.; Evans, R.M. Isolation and characterization of mammalian HDAC10, a novel histone deacetylase. J. Biol. Chem. 2002, 277, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Richon, V.M.; Rifkind, R.A.; Marks, P.A. Identification of a transcriptional repressor related to the noncatalytic domain of histone deacetylases 4 and 5. Proc. Natl. Acad. Sci. USA 2000, 97, 1056–1061. [Google Scholar] [CrossRef] [PubMed]
- Seto, E.; Yoshida, M. Erasers of histone acetylation: The histone deacetylase enzymes. Cold Spring Harb Perspect Biol 2014, 6, a018713. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Kijima, M.; Akita, M.; Beppu, T. Potent and specific inhibition of mammalian histone deacetylase both in vivo and in vitro by trichostatin A. J. Biol. Chem. 1990, 265, 17174–17179. [Google Scholar] [CrossRef]
- Graff, J.; Tsai, L.H. The potential of HDAC inhibitors as cognitive enhancers. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 311–330. [Google Scholar] [CrossRef]
- Zhang, G.Z.; Li, M.W.; Lu, C.; Ai, J.W.; Bi, L.H.; Chen, J.; Chen, P.; Chen, T.; Cheng, D.J. Chinese Sericulture; Shanghai Scientific & Technical Publishers: Shanghai, China, 2020. [Google Scholar]
- DeBerardinis, R.J.; Cheng, T. Q’s next: The diverse functions of glutamine in metabolism, cell biology and cancer. Oncogene 2010, 29, 313–324. [Google Scholar] [CrossRef]
- Silva Gunawardene, Y.I.; Chow, B.K.; He, J.G.; Chan, S.M. The shrimp FAMeT cDNA is encoded for a putative enzyme involved in the methylfarnesoate (MF) biosynthetic pathway and is temporally expressed in the eyestalk of different sexes. Insect Biochem. Mol. Biol. 2001, 31, 1115–1124. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ding, Q.; Yagi, K.J.; Tobe, S.S. Terminal stages in juvenile hormone biosynthesis in corpora allata of Diploptera punctata: Developmental changes in enzyme activity and regulation by allatostatins. J. Insect Physiol. 1994, 40, 217–233. [Google Scholar] [CrossRef]
- Williamson, M.; Lenz, C.; Winther, A.M.; Nassel, D.R.; Grimmelikhuijzen, C.J. Molecular cloning, genomic organization, and expression of a B-type (cricket-type) allatostatin preprohormone from Drosophila melanogaster. Biochem. Biophys. Res. Commun. 2001, 281, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T.; Thibert, C.; Royer, C.; Kanda, T.; Abraham, E.; Kamba, M.; Komoto, N.; Thomas, J.L.; Mauchamp, B.; Chavancy, G.; et al. Germline transformation of the silkworm Bombyx mori L. using a piggyBac transposon-derived vector. Nat. Biotechnol. 2000, 18, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.S.; Eickbush, T.H.; Herrera, R.J.; Lizardi, P.M. A highly reiterated family of transcribed oligo(A)-terminated, interspersed DNA elements in the genome of Bombyx mori. J. Mol. Biol. 1986, 187, 465–478. [Google Scholar] [CrossRef]
- Osanai-Futahashi, M.; Suetsugu, Y.; Mita, K.; Fujiwara, H. Genome-wide screening and characterization of transposable elements and their distribution analysis in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2008, 38, 1046–1057. [Google Scholar] [CrossRef] [PubMed]
- Benhamed, F.; Denechaud, P.D.; Lemoine, M.; Robichon, C.; Moldes, M.; Bertrand-Michel, J.; Ratziu, V.; Serfaty, L.; Housset, C.; Capeau, J.; et al. The lipogenic transcription factor ChREBP dissociates hepatic steatosis from insulin resistance in mice and humans. J. Clin. Investig. 2012, 122, 2176–2194. [Google Scholar] [CrossRef] [PubMed]
- Meng, M.; Cheng, D.J.; Peng, J.; Kang, L.X.; Wang, Y.H.; Qian, W.L.; Xia, Q.Y. Identification and Expression Profiling of Farnesoic Acid O-methyltransferase Gene in Silkworm, Bombyx mori. Sci. Seric. 2013, 39, 0680–0688. [Google Scholar]
- Zhang, J.; Zhu, M.; Li, Q.; Tang, T.; Wen, L.; Zhong, J.; Zhang, R.; Yu, X.Q.; Lu, Y. Genome-wide identification and characterization of basic helix-loop-helix transcription factors in Spodoptera litura upon pathogen infection. Insect Sci. 2022, 29, 977–992. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Tanaka, H. Construction of shRNA expression plasmids for silkworm cell lines using single-stranded DNA and Bst DNA polymerase. Methods Mol. Biol. 2013, 942, 347–355. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Set | 5′-Flanking Region (~3 kb) | |||||
|---|---|---|---|---|---|---|
| S | π | θw | Tajima’s D | Fu and Li’s D | Fu and Li’s F | |
| Domestic (n = 18) | 232 | 0.00842 | 0.02202 | −2.62506 | −4.00135 | −4.1804 |
| Wild (n = 8) | 123 | 0.01751 | 0.01622 | 0.34000 | 0.97307 | 0.92106 |
| With TE (n = 4) | 377 | 0.06996 | 0.06066 | 1.5464 | 1.56333 | 1.68282 |
| Without TE (n = 4) | 126 | 0.02301 | 0.02326 | −0.35185 | −0.27742 | −0.31234 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, X.; Jiang, L.; Zhao, P.; Lin, Y.; Zhang, Y.; Xia, Q. Domestication Gene Mlx and Its Partner Mondo Are Involved in Controlling the Larval Body Size and Cocoon Shell Weight of Bombyx mori. Int. J. Mol. Sci. 2024, 25, 3427. https://doi.org/10.3390/ijms25063427
Qin X, Jiang L, Zhao P, Lin Y, Zhang Y, Xia Q. Domestication Gene Mlx and Its Partner Mondo Are Involved in Controlling the Larval Body Size and Cocoon Shell Weight of Bombyx mori. International Journal of Molecular Sciences. 2024; 25(6):3427. https://doi.org/10.3390/ijms25063427
Chicago/Turabian StyleQin, Xiaoxuan, Liang Jiang, Ping Zhao, Ying Lin, Yi Zhang, and Qingyou Xia. 2024. "Domestication Gene Mlx and Its Partner Mondo Are Involved in Controlling the Larval Body Size and Cocoon Shell Weight of Bombyx mori" International Journal of Molecular Sciences 25, no. 6: 3427. https://doi.org/10.3390/ijms25063427