
Follicular Fluid-Derived Extracellular Vesicles Influence on In Vitro Maturation of Equine Oocyte: Impact on Cumulus Cell Viability, Expansion and Transcriptome
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
2.1. ffEV Characterization
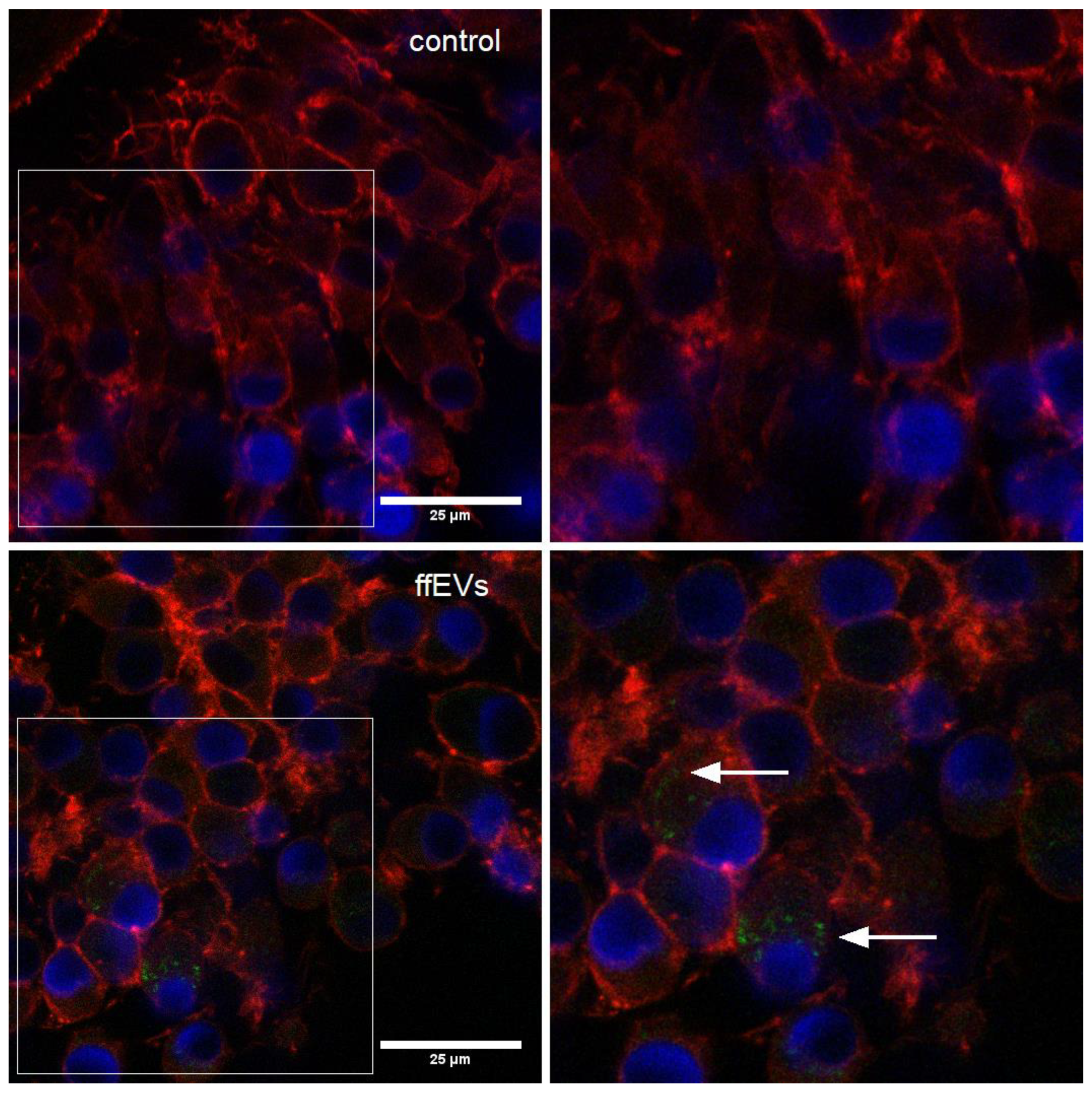
2.2. ffEV Labeling and Uptake by COCs
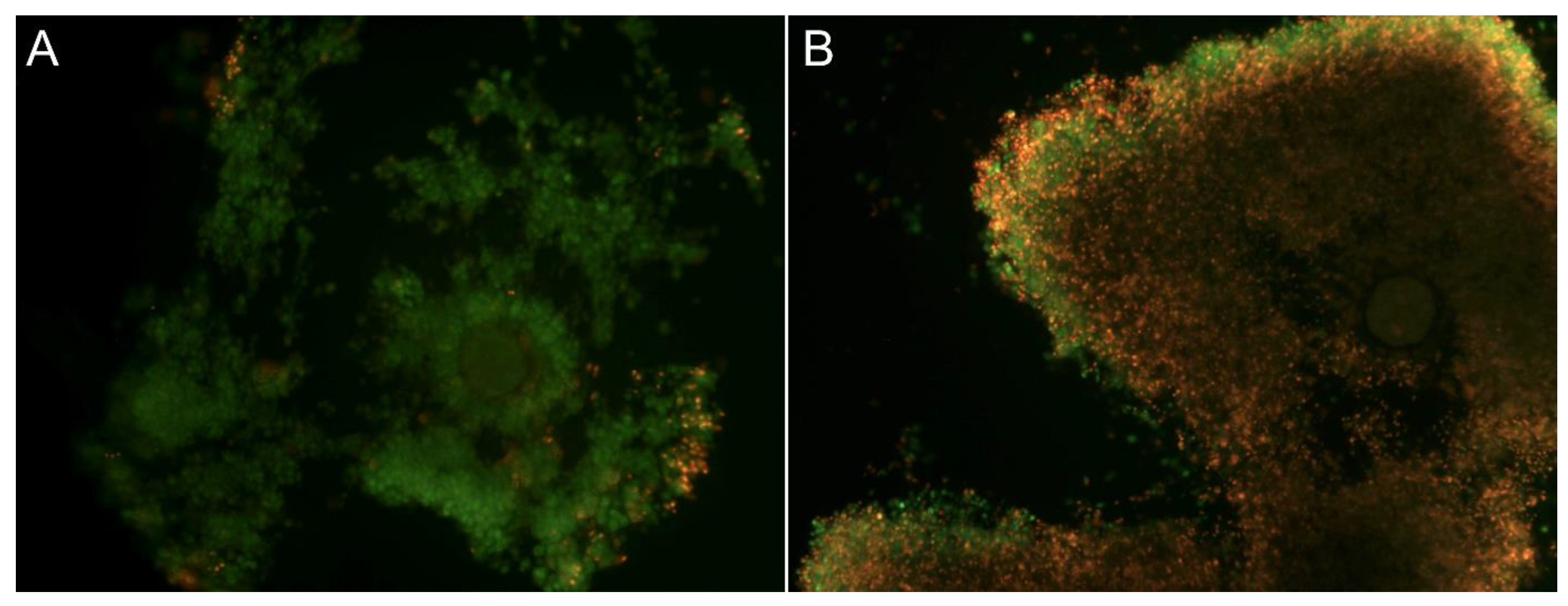
2.3. Cumulus Expansion
2.4. Viability
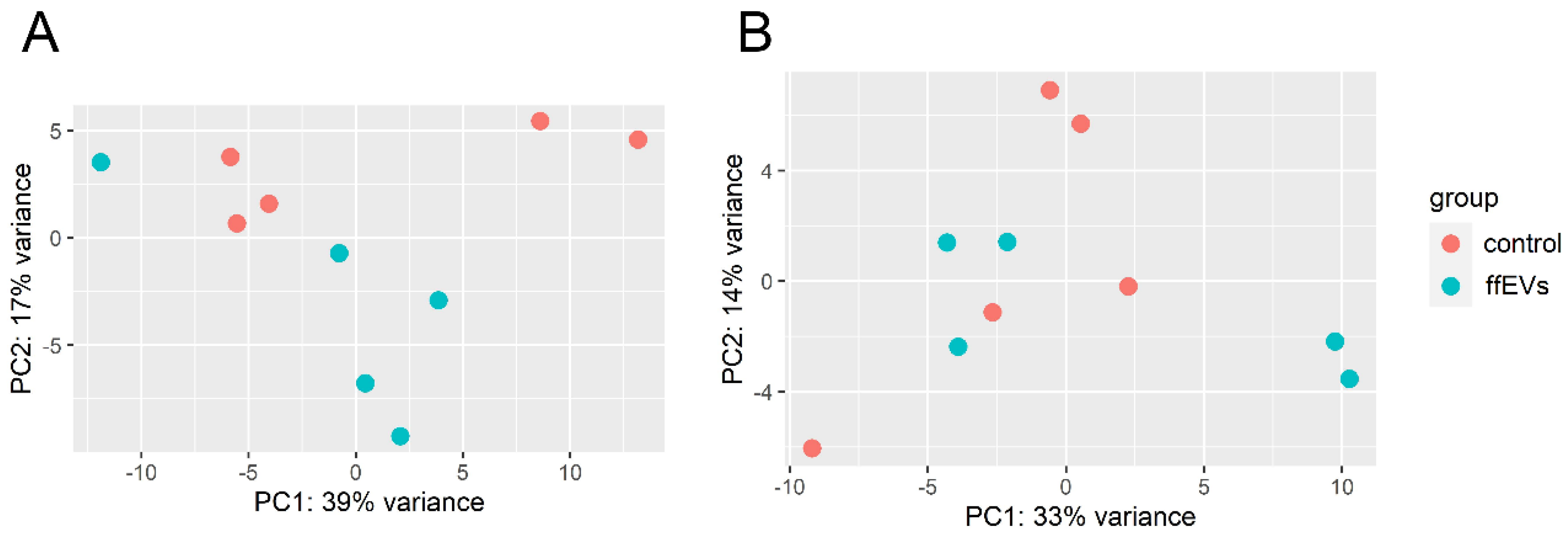
2.5. Mapped Reads, Statistics and Global Expression Profiles
2.6. Transcriptome Alterations in CC Supplemented with ffEVs during IVM
2.7. qPCR Validation of RNA-Seq Results
3. Discussion
4. Materials and Methods
4.1. Experimental Design
- Compacted (Cp) control;
- Cp + ffEVs;
- Expanded (Ex) control;
- Ex + ffEVs.
4.2. Source of Ovaries
4.3. Follicular Fluid and COC Collection
4.4. In Vitro Maturation of Oocyte (IVM)
4.5. Extracellular Vesicle (EV) Extraction
4.6. Transmission Electron Microscopy (TEM)
4.7. Nanoparticle Tracking Analysis (NTA)
4.8. Flow Cytometry (FC)
4.9. ffEV Labeling and Uptake by COCs
4.10. Cumulus Expansion Assessment
4.11. Cumulus Viability Assessment
4.12. RNA Isolation
4.13. 3′ mRNA-Seq Library Preparation and Sequencing
4.14. Transcriptome Analysis and Data Analysis
4.15. RT-PCR and qPCR
4.16. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reader, K.; Stanton, J.-A.; Juengel, J. The Role of Oocyte Organelles in Determining Developmental Competence. Biology 2017, 6, 35. [Google Scholar] [CrossRef]
- Sirard, M.-A.; Richard, F.; Blondin, P.; Robert, C. Contribution of the Oocyte to Embryo Quality. Theriogenology 2006, 65, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Eppig, J.J. FSH Stimulates Hyaluronic Acid Synthesis by Oocyte–Cumulus Cell Complexes from Mouse Preovulatory Follicles. Nature 1979, 281, 483–484. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.-J.; Wu, S.-N.; Shen, J.-P.; Wang, D.-H.; Kong, X.-W.; Lu, A.; Li, Y.-J.; Zhou, H.-X.; Zhao, Y.-F.; Liang, C.-G. The Beneficial Effects of Cumulus Cells and Oocyte-Cumulus Cell Gap Junctions Depends on Oocyte Maturation and Fertilization Methods in Mice. PeerJ 2016, 4, e1761. [Google Scholar] [CrossRef] [PubMed]
- Pangas, S.A.; Matzuk, M.M. The Art and Artifact of GDF9 Activity: Cumulus Expansion and the Cumulus Expansion-Enabling Factor1. Biol. Reprod. 2005, 73, 582–585. [Google Scholar] [CrossRef] [PubMed]
- Fülöp, C.; Salustri, A.; Hascall, V.C. Coding Sequence of a Hyaluronan Synthase Homologue Expressed during Expansion of the Mouse Cumulus–Oocyte Complex. Arch. Biochem. Biophys. 1997, 337, 261–266. [Google Scholar] [CrossRef]
- Davis, B.J.; Lennard, D.E.; Lee, C.A.; Tiano, H.F.; Morham, S.G.; Wetsel, W.C.; Langenbach, R. Anovulation in Cyclooxygenase-2-Deficient Mice Is Restored by Prostaglandin E2 and Interleukin-1β. Endocrinology 1999, 140, 2685–2695. [Google Scholar] [CrossRef]
- Marei, W.F.; Abayasekara, D.R.E.; Wathes, D.C.; Fouladi-Nashta, A.A. Role of PTGS2-Generated PGE2 during Gonadotrophin-Induced Bovine Oocyte Maturation and Cumulus Cell Expansion. Reprod. Biomed. Online 2014, 28, 388–400. [Google Scholar] [CrossRef]
- Chen, L.; Russell, P.T.; Larsen, W.J. Functional Significance of Cumulus Expansion in the Mouse: Roles for the Preovulatory Synthesis of Hyaluronic Acid within the Cumulus Mass. Mol. Reprod. Dev. 1993, 34, 87–93. [Google Scholar] [CrossRef]
- Hess, K.A.; Chen, L.; Larsen, W.J. Inter-α-Inhibitor Binding to Hyaluronan in the Cumulus Extracellular Matrix Is Required for Optimal Ovulation and Development of Mouse Oocytes1. Biol. Reprod. 1999, 61, 436–443. [Google Scholar] [CrossRef]
- Rodgers, R.J.; Irving-Rodgers, H.F. Formation of the Ovarian Follicular Antrum and Follicular Fluid. Biol. Reprod. 2010, 82, 1021–1029. [Google Scholar] [CrossRef]
- Nandi, S.; Kumar, V.G.; Manjunatha, B.M.; Gupta, P.S.P. Biochemical Composition of Ovine Follicular Fluid in Relation to Follicle Size. Dev. Growth Differ. 2007, 49, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Gosden, R.G.; Hunter, R.H.F.; Telfer, E.; Torrance, C.; Brown, N. Physiological Factors Underlying the Formation of Ovarian Follicular Fluid. Reproduction 1988, 82, 813–825. [Google Scholar] [CrossRef]
- McNatty, K.P. Follicular Fluid. In The Vertebrate Ovary; Jones, R.E., Ed.; Plenum Press: New York, NY, USA, 1978. [Google Scholar]
- Espey, L.L.; Lipner, H. Ovulation. In The Physiology of Reproduction; Knobil, E., Neill, J.D., Eds.; Raven Press: New York, NY, USA, 1994. [Google Scholar]
- Ambekar, A.S.; Nirujogi, R.S.; Srikanth, S.M.; Chavan, S.; Kelkar, D.S.; Hinduja, I.; Zaveri, K.; Prasad, T.S.K.; Harsha, H.C.; Pandey, A.; et al. Proteomic Analysis of Human Follicular Fluid: A New Perspective towards Understanding Folliculogenesis. J. Proteom. 2013, 87, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, R.; Musumeci, P.; Ragusa, M.; Barbagallo, D.; Scalia, M.; Zimbone, M.; Lo Faro, J.M.; Borzì, P.; Scollo, P.; Purrello, M.; et al. Ovarian Aging Increases Small Extracellular Vesicle CD81+ Release in Human Follicular Fluid and Influences MiRNA Profiles. Aging 2020, 12, 12324–12341. [Google Scholar] [CrossRef] [PubMed]
- da Silveira, J.C.; Andrade, G.M.; Simas, R.C.; Martins-Júnior, H.A.; Eberlin, M.N.; Smith, L.C.; Perecin, F.; Meirelles, F.V. Lipid Profile of Extracellular Vesicles and Their Relationship with Bovine Oocyte Developmental Competence: New Players in Intra Follicular Cell Communication. Theriogenology 2021, 174, 1–8. [Google Scholar] [CrossRef]
- da Silveira, J.C.; Veeramachaneni, D.N.R.; Winger, Q.A.; Carnevale, E.M.; Bouma, G.J. Cell-Secreted Vesicles in Equine Ovarian Follicular Fluid Contain MiRNAs and Proteins: A Possible New Form of Cell Communication Within the Ovarian Follicle. Biol. Reprod. 2012, 86, 71. [Google Scholar] [CrossRef]
- Mobarak, H.; Heidarpour, M.; Lolicato, F.; Nouri, M.; Rahbarghazi, R.; Mahdipour, M. Physiological Impact of Extracellular Vesicles on Female Reproductive System; Highlights to Possible Restorative Effects on Female Age-related Fertility. BioFactors 2019, 45, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Javadi, M.; Soleimani Rad, J.; Pashaiasl, M.; Farashah, M.S.G.; Roshangar, L. The Effects of Plasma-Derived Extracellular Vesicles on Cumulus Expansion and Oocyte Maturation in Mice. Reprod. Biol. 2022, 22, 100593. [Google Scholar] [CrossRef]
- Chen, Y.; Li, G.; Liu, M.-L. Microvesicles as Emerging Biomarkers and Therapeutic Targets in Cardiometabolic Diseases. Genom. Proteom. Bioinform. 2018, 16, 50–62. [Google Scholar] [CrossRef]
- Mashouri, L.; Yousefi, H.; Aref, A.R.; Ahadi, A.m.; Molaei, F.; Alahari, S.K. Exosomes: Composition, Biogenesis, and Mechanisms in Cancer Metastasis and Drug Resistance. Mol. Cancer 2019, 18, 75. [Google Scholar] [CrossRef]
- Kakarla, R.; Hur, J.; Kim, Y.J.; Kim, J.; Chwae, Y.-J. Apoptotic Cell-Derived Exosomes: Messages from Dying Cells. Exp. Mol. Med. 2020, 52, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.P.; Lau, N.C.; Garrett-Engele, P.; Grimson, A.; Schelter, J.M.; Castle, J.; Bartel, D.P.; Linsley, P.S.; Johnson, J.M. Microarray Analysis Shows That Some MicroRNAs Downregulate Large Numbers of Target MRNAs. Nature 2005, 433, 769–773. [Google Scholar] [CrossRef] [PubMed]
- Sætrom, P.; Snøve, O.; Rossi, J.J. Epigenetics and MicroRNAs. Pediatr. Res. 2007, 61, 17R–23R. [Google Scholar] [CrossRef]
- Lei, L.; Jin, S.; Gonzalez, G.; Behringer, R.R.; Woodruff, T.K. The Regulatory Role of Dicer in Folliculogenesis in Mice. Mol. Cell Endocrinol. 2010, 315, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Santonocito, M.; Vento, M.; Guglielmino, M.R.; Battaglia, R.; Wahlgren, J.; Ragusa, M.; Barbagallo, D.; Borzì, P.; Rizzari, S.; Maugeri, M.; et al. Molecular Characterization of Exosomes and Their MicroRNA Cargo in Human Follicular Fluid: Bioinformatic Analysis Reveals That Exosomal MicroRNAs Control Pathways Involved in Follicular Maturation. Fertil. Steril. 2014, 102, 1751–1761.e1. [Google Scholar] [CrossRef]
- da Silveira, J.C.; Winger, Q.A.; Bouma, G.J.; Carnevale, E.M. Effects of Age on Follicular Fluid Exosomal MicroRNAs and Granulosa Cell Transforming Growth Factor-β Signalling during Follicle Development in the Mare. Reprod. Fertil. Dev. 2015, 27, 897. [Google Scholar] [CrossRef] [PubMed]
- da Silveira, J.C.; de Andrade, G.M.; Nogueira, M.F.G.; Meirelles, F.V.; Perecin, F. Involvement of MiRNAs and Cell-Secreted Vesicles in Mammalian Ovarian Antral Follicle Development. Reprod. Sci. 2015, 22, 1474–1483. [Google Scholar] [CrossRef]
- Sohel, M.d.M.H.; Hoelker, M.; Noferesti, S.S.; Salilew-Wondim, D.; Tholen, E.; Looft, C.; Rings, F.; Uddin, M.J.; Spencer, T.E.; Schellander, K.; et al. Exosomal and Non-Exosomal Transport of Extra-Cellular MicroRNAs in Follicular Fluid: Implications for Bovine Oocyte Developmental Competence. PLoS ONE 2013, 8, e78505. [Google Scholar] [CrossRef]
- de Ávila, A.C.F.C.M.; da Silveira, J.C. Role of Extracellular Vesicles during Oocyte Maturation and Early Embryo Development. Reprod. Fertil. Dev. 2020, 32, 56. [Google Scholar] [CrossRef]
- Czernek, L.; Düchler, M. Exosomes as Messengers between Mother and Fetus in Pregnancy. Int. J. Mol. Sci. 2020, 21, 4264. [Google Scholar] [CrossRef]
- Hasan, M.M.; Viil, J.; Lättekivi, F.; Ord, J.; Reshi, Q.U.A.; Jääger, K.; Velthut-Meikas, A.; Andronowska, A.; Jaakma, Ü.; Salumets, A.; et al. Bovine Follicular Fluid and Extracellular Vesicles Derived from Follicular Fluid Alter the Bovine Oviductal Epithelial Cells Transcriptome. Int. J. Mol. Sci. 2020, 21, 5365. [Google Scholar] [CrossRef]
- de Alcântara-Neto, A.S.; Cuello, C.; Uzbekov, R.; Bauersachs, S.; Mermillod, P.; Almiñana, C. Oviductal Extracellular Vesicles Enhance Porcine In Vitro Embryo Development by Modulating the Embryonic Transcriptome. Biomolecules 2022, 12, 1300. [Google Scholar] [CrossRef]
- Bauersachs, S.; Mermillod, P.; Almiñana, C. The Oviductal Extracellular Vesicles’ RNA Cargo Regulates the Bovine Embryonic Transcriptome. Int. J. Mol. Sci. 2020, 21, 1303. [Google Scholar] [CrossRef] [PubMed]
- da Silveira, J.C.; Carnevale, E.M.; Winger, Q.A.; Bouma, G.J. Regulation of ACVR1 and ID2 by Cell-Secreted Exosomes during Follicle Maturation in the Mare. Reprod. Biol. Endocrinol. 2014, 12, 44. [Google Scholar] [CrossRef]
- Hung, W.-T.; Hong, X.; Christenson, L.K.; McGinnis, L.K. Extracellular Vesicles from Bovine Follicular Fluid Support Cumulus Expansion. Biol. Reprod. 2015, 93, 117. [Google Scholar] [CrossRef] [PubMed]
- Hung, W.-T.; Navakanitworakul, R.; Khan, T.; Zhang, P.; Davis, J.S.; McGinnis, L.K.; Christenson, L.K. Stage-Specific Follicular Extracellular Vesicle Uptake and Regulation of Bovine Granulosa Cell Proliferation. Biol. Reprod. 2017, 97, 644–655. [Google Scholar] [CrossRef] [PubMed]
- Gabryś, J.; Kij-Mitka, B.; Sawicki, S.; Kochan, J.; Nowak, A.; Łojko, J.; Karnas, E.; Bugno-Poniewierska, M. Extracellular Vesicles from Follicular Fluid May Improve the Nuclear Maturation Rate of In Vitro Matured Mare Oocytes. Theriogenology 2022, 188, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Lange-Consiglio, A.; Perrini, C.; Albini, G.; Modina, S.; Lodde, V.; Orsini, E.; Esposti, P.; Cremonesi, F. Oviductal Microvesicles and Their Effect on In Vitro Maturation of Canine Oocytes. Reproduction 2017, 154, 167–180. [Google Scholar] [CrossRef]
- Matsuno, Y.; Onuma, A.; Fujioka, Y.A.; Yasuhara, K.; Fujii, W.; Naito, K.; Sugiura, K. Effects of Exosome-like Vesicles on Cumulus Expansion in Pigs In Vitro. J. Reprod. Dev. 2017, 63, 51–58. [Google Scholar] [CrossRef]
- Mateo-Otero, Y.; Yeste, M.; Roca, J.; Llavanera, M.; Bucci, D.; Galeati, G.; Spinaci, M.; Barranco, I. Seminal Extracellular Vesicles Subsets Modulate Gene Expression in Cumulus Cells of Porcine In Vitro Matured Oocytes. Sci. Rep. 2022, 12, 19096. [Google Scholar] [CrossRef]
- Diaz, F.J.; O’Brien, M.J.; Wigglesworth, K.; Eppig, J.J. The Preantral Granulosa Cell to Cumulus Cell Transition in the Mouse Ovary: Development of Competence to Undergo Expansion. Dev. Biol. 2006, 299, 91–104. [Google Scholar] [CrossRef]
- Liu, Z.; Fan, H.-Y.; Wang, Y.; Richards, J.S. Targeted Disruption of Mapk14 (P38MAPKα) in Granulosa Cells and Cumulus Cells Causes Cell-Specific Changes in Gene Expression Profiles That Rescue COC Expansion and Maintain Fertility. Mol. Endocrinol. 2010, 24, 1794–1804. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.-Y.; Sun, Q.-Y. Involvement of Mitogen-Activated Protein Kinase Cascade During Oocyte Maturation and Fertilization in Mammals1. Biol. Reprod. 2004, 70, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.-Q.; Denegre, J.M.; Wigglesworth, K.; Pendola, F.L.; O’Brien, M.J.; Eppig, J.J. Oocyte-Dependent Activation of Mitogen-Activated Protein Kinase (ERK1/2) in Cumulus Cells Is Required for the Maturation of the Mouse Oocyte–Cumulus Cell Complex. Dev. Biol. 2003, 263, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Cibelli, J.B.; Iager, A.E.; Otu, H.H. Genes Differentially Expressed by Cumulus Cells and Assays Using Same to Dentify Pregnancy Competent Oocytes. Faculty Publications from the Department of Electrical and Computer Engineering. 2009. Available online: http://digitalcommons.unl.edu/electricalengineeringfacpub/444 (accessed on 12 December 2023).
- Salhab, M.; Tosca, L.; Cabau, C.; Papillier, P.; Perreau, C.; Dupont, J.; Mermillod, P.; Uzbekova, S. Kinetics of Gene Expression and Signaling in Bovine Cumulus Cells throughout IVM in Different Mediums in Relation to Oocyte Developmental Competence, Cumulus Apoptosis and Progesterone Secretion. Theriogenology 2011, 75, 90–104. [Google Scholar] [CrossRef]
- Hu, C.-L.; Cowan, R.G.; Harman, R.M.; Quirk, S.M. Cell Cycle Progression and Activation of Akt Kinase Are Required for Insulin-Like Growth Factor I-Mediated Suppression of Apoptosis in Granulosa Cells. Mol. Endocrinol. 2004, 18, 326–338. [Google Scholar] [CrossRef]
- Yuan, Y.Q.; Van Soom, A.; Leroy, J.L.M.R.; Dewulf, J.; Van Zeveren, A.; de Kruif, A.; Peelman, L.J. Apoptosis in Cumulus Cells, but Not in Oocytes, May Influence Bovine Embryonic Developmental Competence. Theriogenology 2005, 63, 2147–2163. [Google Scholar] [CrossRef]
- Lee, K.S.; Joo, B.S.; Na, Y.J.; Yoon, M.S.; Choi, O.H.; Kim, W.W. Cumulus Cells Apoptosis as an Indicator to Predict the Quality of Oocytes and the Outcome of IVF-ET. J. Assist. Reprod. Genet. 2001, 18, 490–498. [Google Scholar] [CrossRef]
- Abate, C.; Marshak, D.R.; Curran, T. Fos Is Phosphorylated by P34cdc2, CAMP-Dependent Protein Kinase and Protein Kinase C at Multiple Sites Clustered within Regulatory Regions. Oncogene 1991, 6, 2179–2185. [Google Scholar] [PubMed]
- Tanos, T.; Marinissen, M.J.; Leskow, F.C.; Hochbaum, D.; Martinetto, H.; Gutkind, J.S.; Coso, O.A. Phosphorylation of C-Fos by Members of the P38 MAPK Family. J. Biol. Chem. 2005, 280, 18842–18852. [Google Scholar] [CrossRef] [PubMed]
- Angel, P.; Karin, M. The Role of Jun, Fos and the AP-1 Complex in Cell-Proliferation and Transformation. Biochim. Biophys. Acta-Rev. Cancer 1991, 1072, 129–157. [Google Scholar] [CrossRef]
- Ohki, M.; Ohki, Y.; Ishihara, M.; Nishida, C.; Tashiro, Y.; Akiyama, H.; Komiyama, H.; Lund, L.R.; Nitta, A.; Yamada, K.; et al. Tissue Type Plasminogen Activator Regulates Myeloid-Cell Dependent Neoangiogenesis during Tissue Regeneration. Blood 2010, 115, 4302–4312. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhou, J.; Wei, X.; Chen, R.; Geng, J.; Zheng, R.; Chai, J.; Li, F.; Jiang, S. MiR-144 and Targets, c-Fos and Cyclooxygenase-2 (COX2), Modulate Synthesis of PGE2 in the Amnion during Pregnancy and Labor. Sci. Rep. 2016, 6, 27914. [Google Scholar] [CrossRef]
- Stocco, C.; Telleria, C.; Gibori, G. The Molecular Control of Corpus Luteum Formation, Function, and Regression. Endocr. Rev. 2007, 28, 117–149. [Google Scholar] [CrossRef] [PubMed]
- Perugini, M.; Kok, C.H.; Brown, A.L.; Wilkinson, C.R.; Salerno, D.G.; Young, S.M.; Diakiw, S.M.; Lewis, I.D.; Gonda, T.J.; D’Andrea, R.J. Repression of Gadd45α by Activated FLT3 and GM-CSF Receptor Mutants Contributes to Growth, Survival and Blocked Differentiation. Leukemia 2009, 23, 729–738. [Google Scholar] [CrossRef]
- Chang, B.; Liu, G.; Yang, G.; Mercado-Uribe, I.; Huang, M.; Liu, J. REDD1 Is Required for RAS-Mediated Transformation of Human Ovarian Epithelial Cells. Cell Cycle 2009, 8, 780–786. [Google Scholar] [CrossRef]
- Goudet, G.; Belin, F.; Bezard, J.; Gerard, N. Maturation-Promoting Factor (MPF) and Mitogen Activated Protein Kinase (MAPK) Expression in Relation to Oocyte Competence for in-Vitro Maturation in the Mare. Mol. Hum. Reprod. 1998, 4, 563–570. [Google Scholar] [CrossRef]
- Hinrichs, K. The Equine Oocyte: Factors Affecting Meiotic and Developmental Competence. Mol. Reprod. Dev. 2010, 77, 651–661. [Google Scholar] [CrossRef]
- Andrews, S.C.; Wood, M.D.; Tunster, S.J.; Barton, S.C.; Surani, M.A.; John, R.M. Cdkn1c (P57 Kip2) Is the Major Regulator of Embryonic Growth within Its Imprinted Domain on Mouse Distal Chromosome 7. BMC Dev. Biol. 2007, 7, 53. [Google Scholar] [CrossRef]
- Spate, L.D.; Brown, A.N.; Redel, B.K.; Whitworth, K.M.; Murphy, C.N.; Prather, R.S. Dickkopf-Related Protein 1 Inhibits the WNT Signaling Pathway and Improves Pig Oocyte Maturation. PLoS ONE 2014, 9, e95114. [Google Scholar] [CrossRef] [PubMed]
- Harwood, B.N.; Cross, S.K.; Radford, E.E.; Haac, B.E.; De Vries, W.N. Members of the WNT Signaling Pathways Are Widely Expressed in Mouse Ovaries, Oocytes, and Cleavage Stage Embryos. Dev. Dyn. 2008, 237, 1099–1111. [Google Scholar] [CrossRef]
- Schlesinger, A.; Shelton, C.; Maloof, J.; Meneghini, M.; Bowerman, B. Wnt Pathway Components Orient a Mitotic Spindle in the Early Caenorhabditis Elegans Embryo without Requiring Gene Transcription in the Responding Cell. Genes. Dev. 1999, 13, 2028–2038. [Google Scholar] [CrossRef] [PubMed]
- Posor, Y.; Eichhorn-Grünig, M.; Haucke, V. Phosphoinositides in Endocytosis. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2015, 1851, 794–804. [Google Scholar] [CrossRef]
- Nowak, A.; Kochan, J.; Witarski, W.; Okólski, A. In Vitro Maturation of Equine Oocytes Followed by Two Vitrification Protocols and Subjected to Either Intracytoplasmic Sperm Injection (ICSI) or Parthenogenic Activation. Theriogenology 2021, 162, 42–48. [Google Scholar] [CrossRef]
- Takasugi, M. An Improved Fluoroghromatic Cytotoxic Test. Transplantation 1971, 12, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. The Single-Step Method of RNA Isolation by Acid Guanidinium Thiocyanate–Phenol–Chloroform Extraction: Twenty-Something Years On. Nat. Protoc. 2006, 1, 581–585. [Google Scholar] [CrossRef]
- Maisarah, Y.; Hashida, H.N.; Yusmin, M.-Y. The Challenge of Getting a High Quality of RNA from Oocyte for Gene Expression Study. Vet. Res. Forum 2020, 11, 179–184. [Google Scholar] [CrossRef]
- Dodt, M.; Roehr, J.; Ahmed, R.; Dieterich, C. FLEXBAR—Flexible Barcode and Adapter Processing for Next-Generation Sequencing Platforms. Biology 2012, 1, 895–905. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Aken, B.L.; Ayling, S.; Barrell, D.; Clarke, L.; Curwen, V.; Fairley, S.; Fernandez Banet, J.; Billis, K.; García Girón, C.; Hourlier, T.; et al. The Ensembl Gene Annotation System. Database 2016, 2016, baw093. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential Expression Analysis for Sequence Count Data. Nat. Preced. 2010. [Google Scholar] [CrossRef]
- Thomas, P.D.; Ebert, D.; Muruganujan, A.; Mushayahama, T.; Albou, L.; Mi, H. PANTHER: Making Genome-scale Phylogenetics Accessible to All. Protein Sci. 2022, 31, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A Graphical Gene-Set Enrichment Tool for Animals and Plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef] [PubMed]
- Bu, D.; Luo, H.; Huo, P.; Wang, Z.; Zhang, S.; He, Z.; Wu, Y.; Zhao, L.; Liu, J.; Guo, J.; et al. KOBAS-i: Intelligent Prioritization and Exploratory Visualization of Biological Functions for Gene Enrichment Analysis. Nucleic Acids Res. 2021, 49, W317–W325. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| COC | Gene Symbol | Gene | Description | Log2fold | Padj |
|---|---|---|---|---|---|
| Cp | ENSECAG00000013822 | KIF5C | kinesin family member 5C | −1.82667 | 9.44 × 10−5 |
| ENSECAG00000024531 | ADM | adrenomedullin | −1.85264 | 9.44 × 10−5 | |
| ENSECAG00000009958 | DUSP1 | dual specificity phosphatase 1 | −1.24247 | 9.53 × 10−5 | |
| ENSECAG00000020042 | GAS8 | growth arrest specific 8 | 5.207698 | 0.002822 | |
| ENSECAG00000014260 | FOS | Fos proto-onco, AP-1 transcription factor subunit | −1.43215 | 0.006363 | |
| ENSECAG00000016495 | NUSAP1 | nucleolar and spindle-associated protein 1 | −1.36047 | 0.006363 | |
| ENSECAG00000004107 | DDIT4 | DNA damage-inducible transcript 4 | −1.32112 | 0.006983 | |
| ENSECAG00000024055 | JUN | Jun proto-oncogene, AP-1 transcription factor subunit | −1.06773 | 0.011582 | |
| ENSECAG00000003112 | protein coding | uncharacterized | 5.77611 | 0.012615 | |
| ENSECAG00000048140 | lncRNA | uncharacterized | 6.058224 | 0.012615 | |
| ENSECAG00000010132 | HMGB2 | high mobility group box 2 | −1.19162 | 0.01456 | |
| ENSECAG00000049390 | protein coding | uncharacterized | 6.568594 | 0.01456 | |
| ENSECAG00000033482 | MAF | MAF bZIP transcription factor | −2.62299 | 0.015728 | |
| ENSECAG00000034891 | FOXL2 | forkhead box L2 | −0.88989 | 0.019069 | |
| ENSECAG00000046492 | lncRNA | uncharacterized | 5.316257 | 0.019069 | |
| ENSECAG00000040631 | GADD45A | growth arrest and DNA damage-inducible alpha | −1.38945 | 0.025639 | |
| ENSECAG00000009294 | PARD6B | par-6 family cell polarity regulator beta | −1.37246 | 0.028419 | |
| ENSECAG00000057832 | lncRNA | uncharacterized | 5.798079 | 0.039461 | |
| ENSECAG00000019564 | protein coding | uncharacterized | 5.979852 | 0.043859 | |
| ENSECAG00000024127 | CENPF | centromere protein F | −1.20947 | 0.043859 | |
| ENSECAG00000049668 | lncRNA | uncharacterized | 6.15095 | 0.050595 | |
| ENSECAG00000019267 | protein coding | uncharacterized | 5.56764 | 0.050842 | |
| ENSECAG00000004339 | MIS18BP1 | MIS18 binding protein 1 | −1.44315 | 0.050887 | |
| ENSECAG00000058688 | lncRNA | uncharacterized | 1.808977 | 0.05315 | |
| ENSECAG00000048591 | lncRNA | uncharacterized | −5.84793 | 0.053781 | |
| ENSECAG00000010938 | CEP57 | centrosomal protein 57 | −0.80552 | 0.056617 | |
| ENSECAG00000020246 | SEPTIN9 | septin 9 | 0.740021 | 0.056617 | |
| ENSECAG00000034339 | MHCX1 | MHC class I heavy chain | −1.121 | 0.056617 | |
| ENSECAG00000020645 | ASPM | assembly factor for spindle microtubules | −1.31423 | 0.061879 | |
| ENSECAG00000005570 | VMP1 | vacuole membrane protein 1 | 0.634005 | 0.082138 | |
| ENSECAG00000006404 | SLC17A3 | solute carrier family 17 member 3 | 5.861373 | 0.082138 | |
| ENSECAG00000010064 | OLFM3 | olfactomedin 3 | 5.769174 | 0.082138 | |
| ENSECAG00000024196 | KIFC1 | kinesin family member C1 | −1.22762 | 0.082138 | |
| ENSECAG00000047902 | lncRNA | uncharacterized | 5.046975 | 0.082138 | |
| ENSECAG00000000658 | GPNMB | glycoprotein nmb | −5.85274 | 0.086203 | |
| ENSECAG00000010303 | CIAO2A | cytosolic iron–sulfur assembly component 2A | 3.922212 | 0.097695 | |
| Ex | ENSECAG00000009081 | KIAA1210 | KIAA1210 | −1.54446 | 0.000302 |
| ENSECAG00000031389 | CDKN1C | cyclin-dependent kinase inhibitor 1C | 1.240626 | 0.0074 | |
| ENSECAG00000024127 | CENPF | centromere protein F | −1.15973 | 0.010131 | |
| ENSECAG00000000658 | GPNMB | glycoprotein nmb | −2.82836 | 0.068233 | |
| ENSECAG00000011581 | protein coding | uncharacterized | 1.20007 | 0.068233 | |
| ENSECAG00000007309 | FZD5 | frizzled class receptor 5 | 0.685669 | 0.079819 | |
| ENSECAG00000057237 | lncRNA | uncharacterized | −2.32694 | 0.090332 | |
| ENSECAG00000029287 | HOPX | HOP homeobox | 1.480728 | 0.094982 | |
| ENSECAG00000007607 | PTPRG | protein tyrosine phosphatase receptor type G | −0.6582 | 0.097187 | |
| ENSECAG00000013053 | TOP2A | DNA topoisomerase II alpha | −1.05374 | 0.097187 | |
| ENSECAG00000013597 | DAB2 | DAB adaptor protein 2 | −1.01644 | 0.097187 | |
| ENSECAG00000014517 | CDCA3 | cell division cycle-associated 3 | −1.59657 | 0.097187 | |
| ENSECAG00000018981 | ITPKA | inositol-trisphosphate 3-kinase A | −0.76345 | 0.097187 | |
| ENSECAG00000019296 | GUF1 | GTP binding elongation factor GUF1 | 1.778967 | 0.097187 | |
| ENSECAG00000023776 | TTK | TTK protein kinase | −3.71007 | 0.097187 | |
| ENSECAG00000044783 | PCDHGA7 | protocadherin gamma subfamily A, 7 | −3.8215 | 0.097187 |
| GO | Description | Term ID | P | Engaged Genes |
|---|---|---|---|---|
| Biological processes | microtubule-based process | GO:0007017 | 1.34 × 10−3 | KIFC1, KIF5C, PARD6B, NUSAP1 |
| cytoskeleton-dependent cytokinesis | GO:0061640 | 1.47 × 10−3 | SEPTIN9, NUSAP1 | |
| cytokinesis | GO:0000910 | 1.54 × 10−3 | SEPTIN9, NUSAP1 | |
| cell division | GO:0051301 | 2.21 × 10−3 | SEPTIN9, NUSAP1 | |
| negative regulation of stress-activated MAPK cascade | GO:0032873 | 5.16 × 10−3 | DUSP1 | |
| regulation of p38MAPK cascade | GO:1900744 | 5.16 × 10−3 | DUSP1 | |
| negative regulation of intracellular signal transduction | GO:1902532 | 5.38 × 10−3 | DUSP1, DDIT4 | |
| establishment of organelle localization | GO:0051656 | 7.51 × 10−3 | KIF5C, NUSAP1 | |
| cell cycle process | GO:0022402 | 7.80 × 10−3 | SEPTIN9, PARD6B, NUSAP1 | |
| endoderm development | GO:0007492 | 9.01 × 10−3 | DUSP1 | |
| Molecular functions | microtubule binding | GO:0008017 | 4.48 × 10−5 | KIFC1, KIF5C, CEP57, NUSAP1 |
| tubulin binding | GO:0015631 | 9.48 × 10−5 | KIFC1, KIF5C, CEP57, NUSAP1 | |
| microtubule motor activity | GO:0003777 | 2.46 × 10−3 | KIFC1, KIF5C | |
| cytoskeletal protein binding | GO:0008092 | 2.77 × 10−3 | KIFC1, KIF5C, CEP57, NUSAP1 | |
| ATP hydrolysis activity | GO:0016887 | 3.48 × 10−3 | KIFC1, KIF5C | |
| ribonucleoside triphosphate phosphatase activity | GO:0017111 | 3.79 × 10−3 | SEPTIN9, KIFC1, KIF5C | |
| efflux transmembrane transporter activity | GO:0015562 | 3.87 × 10−3 | SLC17A3, ABCG2 | |
| xenobiotic transmembrane transporter activity | GO:0042910 | 5.16 × 10−3 | ABCC1, SLC17A3, SLC6A6 | |
| pyrophosphatase activity | GO:0016462 | 5.44 × 10−3 | SEPTIN9, KIFC1, KIF5C | |
| hydrolase activity, acting on acid anhydrides, in phosphorus-containing anhydrides | GO:0016818 | 5.49 × 10−3 | SEPTIN9, KIFC1, KIF5C | |
| Cellular components | microtubule cytoskeleton | GO:0015630 | 1.69 × 10−5 | SEPTIN9, KIFC1, KIF5C, CEP57, GAS8, NUSAP1 |
| cytoskeleton | GO:0005856 | 3.80 × 10−4 | SEPTIN9, KIFC1, KIF5C, CEP57, GAS8, NUSAP1 | |
| microtubule | GO:0005874 | 9.53 × 10−4 | KIFC1, KIF5C, GAS8 | |
| intracellular non-membrane-bounded organelle | GO:0043232 | 2.59 × 10−3 | SEPTIN9, KIFC1, MIS18BP1, KIF5C, CEP57, GAS8, NUSAP1 | |
| non-membrane-bounded organelle | GO:0043228 | 2.59 × 10−3 | SEPTIN9, KIFC1, MIS18BP1, KIF5C, CEP57, GAS8, NUSAP1 | |
| polymeric cytoskeletal fiber | GO:0099513 | 3.79 × 10−3 | KIFC1, KIF5C, GAS8 | |
| microtubule associated complex | GO:0005875 | 6.94 × 10−3 | KIFC1, KIF5C | |
| supramolecular fiber | GO:0099512 | 8.14 × 10−3 | KIFC1, KIF5C, GAS8 | |
| supramolecular polymer | GO:0099081 | 8.21 × 10−3 | KIFC1, KIF5C, GAS8 | |
| cell cortex | GO:0005938 | 9.00 × 10−3 | SEPTIN9, PARD6B |
| GO | Description | Term ID | P | Engaged Genes |
|---|---|---|---|---|
| Biological processes | negative regulation of mitotic cell cycle | GO:0045930 | 5.86 × 10−4 | TTK, CDKN1C |
| meiotic cell cycle process | GO:1903046 | 7.47 × 10−4 | TOP2A, TTK | |
| meiotic cell cycle | GO:0051321 | 9.55 × 10−4 | TOP2A, TTK | |
| negative regulation of cell cycle | GO:0045786 | 1.34 × 10−3 | TTK, CDKN1C | |
| regulation of mitotic cell cycle | GO:0007346 | 2.03 × 10−3 | TTK, CDKN1C | |
| chromosome segregation | GO:0007059 | 2.53 × 10−3 | TOP2A, TTK | |
| protein localization to kinetochore | GO:0034501 | 4.02 × 10−3 | TTK | |
| sexual reproduction | GO:0019953 | 5.82 × 10−3 | TOP2A, TTK | |
| positive regulation of endocytosis | GO:0045807 | 6.02 × 10−3 | DAB2 | |
| negative regulation of mitotic sister chromatid segregation | GO:0033048 | 7.35 × 10−3 | TTK | |
| Molecular functions | Wnt-protein binding | GO:0017147 | 1.80 × 10−2 | FZD5 |
| ribosome binding | GO:0043022 | 2.13 × 10−2 | GUF1 | |
| ribonucleoprotein complex binding | GO:0043021 | 3.04 × 10−2 | GUF1 | |
| integrin binding | GO:0005178 | 3.50 × 10−2 | GPNMB | |
| protein tyrosine phosphatase activity | GO:0004725 | 4.66 × 10−2 | PTPRG | |
| phosphotransferase activity, alcohol group as acceptor | GO:0016773 | 4.69 × 10−2 | TTK, ITPKA | |
| protein-containing complex binding | GO:0044877 | 4.82 × 10−2 | GUF1, GPNMB | |
| Cellular components | kinetochore | GO:0000776 | 2.45 × 10−2 | TTK |
| condensed chromosome, centromeric region | GO:0000779 | 2.58 × 10−2 | TTK | |
| chromosome, centromeric region | GO:0000775 | 3.24 × 10−2 | TTK | |
| chromosomal region | GO:0098687 | 3.69 × 10−2 | TTK |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gabryś, J.; Gurgul, A.; Szmatoła, T.; Kij-Mitka, B.; Andronowska, A.; Karnas, E.; Kucharski, M.; Wojciechowska-Puchałka, J.; Kochan, J.; Bugno-Poniewierska, M. Follicular Fluid-Derived Extracellular Vesicles Influence on In Vitro Maturation of Equine Oocyte: Impact on Cumulus Cell Viability, Expansion and Transcriptome. Int. J. Mol. Sci. 2024, 25, 3262. https://doi.org/10.3390/ijms25063262
Gabryś J, Gurgul A, Szmatoła T, Kij-Mitka B, Andronowska A, Karnas E, Kucharski M, Wojciechowska-Puchałka J, Kochan J, Bugno-Poniewierska M. Follicular Fluid-Derived Extracellular Vesicles Influence on In Vitro Maturation of Equine Oocyte: Impact on Cumulus Cell Viability, Expansion and Transcriptome. International Journal of Molecular Sciences. 2024; 25(6):3262. https://doi.org/10.3390/ijms25063262
Chicago/Turabian StyleGabryś, Julia, Artur Gurgul, Tomasz Szmatoła, Barbara Kij-Mitka, Aneta Andronowska, Elżbieta Karnas, Mirosław Kucharski, Joanna Wojciechowska-Puchałka, Joanna Kochan, and Monika Bugno-Poniewierska. 2024. "Follicular Fluid-Derived Extracellular Vesicles Influence on In Vitro Maturation of Equine Oocyte: Impact on Cumulus Cell Viability, Expansion and Transcriptome" International Journal of Molecular Sciences 25, no. 6: 3262. https://doi.org/10.3390/ijms25063262