1. Introduction
Inflammatory bowel diseases (IBDs), such as Crohn’s disease (CD) and ulcerative colitis (UC), involve persistent inflammation in the digestive tract. Notably, more than 35% of IBD patients are affected by extraintestinal manifestations (EIMs), which significantly impact the quality of life and require additional treatments. The EIMs include arthropathies, mucocutaneous and ophthalmological manifestations, as well as conditions affecting the hepatobiliary system. Among these, primary sclerosing cholangitis (PSC) is the most common extraintestinal manifestation in IBD, affecting 1.4–7.5% of patients. The gastrointestinal tract is anatomically connected to the hepatobiliary system via the mesenteric and portal veins. Thus, the liver and the biliary system are more susceptible to colonic inflammatory response. Moreover, although the pathogenesis is multifactorial, IBD is associated with an increased intestinal permeability, which provides a favorable environment for the translocation of gut flora to the liver. Further, during recurrent episodes of active colitis, the hepatic microcirculation recruits leukocytes to the sinusoids [
1,
2,
3], leading to liver inflammation and hepatocellular damage.
Polyphenols, a large group of phytochemicals, have been shown to be effective in the prevention and treatment of several diseases [
4], mainly due to their antioxidant, anti-inflammatory and antihyperlipidemic activities. The anti-inflammatory effect of polyphenols is mainly due to the modulation of the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kB) and the reduction of pro-inflammatory cytokines, which represent key targets for the immune therapy in IBD patients [
5]. In the last decade, several studies have reported the contribution of polyphenols, such as chlorogenic acid, isoflavones [
6], proanthocyanidin [
7,
8,
9] and catechins [
10], in maintaining intestinal barrier functions through their antioxidant effects [
11,
12,
13] and in alleviating mucosal inflammation by inhibiting the canonical NF-kB pathway. Moreover, the polyphenols curcumin, quercetin, epigallocathechin-3-gallate and resveratrol have been reported to inhibit the wingless-related integration site (WNT)-activated pathways [
13]. In the intestinal epithelium, the Wnt-signaling pathway promotes proliferation and/or differentiation of stem cells in the intestinal crypts, the epithelial mesenchymal transition (EMT) and the aberrant expansion of cancer stem cells (CSC).
Malus domestica cv. ‘Limoncella’ is a juicy and aromatic apple cultivar [
13] rich in nutritional polyphenols, such as phenolic acids, anthocyanins, flavonoids, and proanthocyanidins, which have several health-promoting effects, including antioxidant and anti-inflammatory activities [
14]. The Limoncella apple develops a yellow peel with a citric taste that lasts until the end of spring. Recently, Riccio et al. have demonstrated the inhibitory effect of Limoncella apple polyphenol extracts (LAPE) on the Wnt/β-catenin pathway in tumor colonic epithelial cells [
15].
In this study, we have evaluated the therapeutic potential of LAPE in a model of colitis associated with liver damage. Hence, the pharmacological effect of LAPE was investigated in a mouse model of colitis induced by 2,4,6-dinitrobenzenesulfonic acid (DNBS) [
16]. Colonic inflammation was studied by monitoring macroscopic parameters, histological damage, cytokine production and intestinal permeability. Therefore, the potential effect of LAPE on colitis-induced liver damage was evaluated by assessing biochemical parameters, inflammatory markers and lipid accumulation. Here, we propose LAPE for the treatment of intestinal inflammation and prevention of colitis-induced liver injury.
3. Discussion
IBDs, such as ulcerative colitis and Crohn’s disease, are chronic relapsing and remitting inflammatory conditions of the gastrointestinal tract. During IBD, pro-inflammatory mediators (such as cytokines) activate the NF-kB and induce an increase in reactive oxygen species (ROS) [
18,
19]. Therefore, the control of inflammation and oxidative stress plays an important role in the treatment of IBD. Notably, chronic intestinal inflammation, as well as the adverse reactions of corticosteroid therapy, are often associated with liver damage [
20] and can lead to chronic liver disease [
21]. Therefore, it is crucial to find new compounds without side effects, which, by reducing intestinal inflammation, could also decrease or prevent liver damage.
Several in vitro and in vivo studies have revealed the preventive and therapeutic effects of polyphenols on intestinal inflammation [
22,
23]. Recently, it has been shown that LAPE inhibits the canonical Wnt pathway in the colonic epithelial cells [
15]. Notably, Wnt/β-catenin signaling is a key regulator of intestinal epithelial homeostasis [
24].
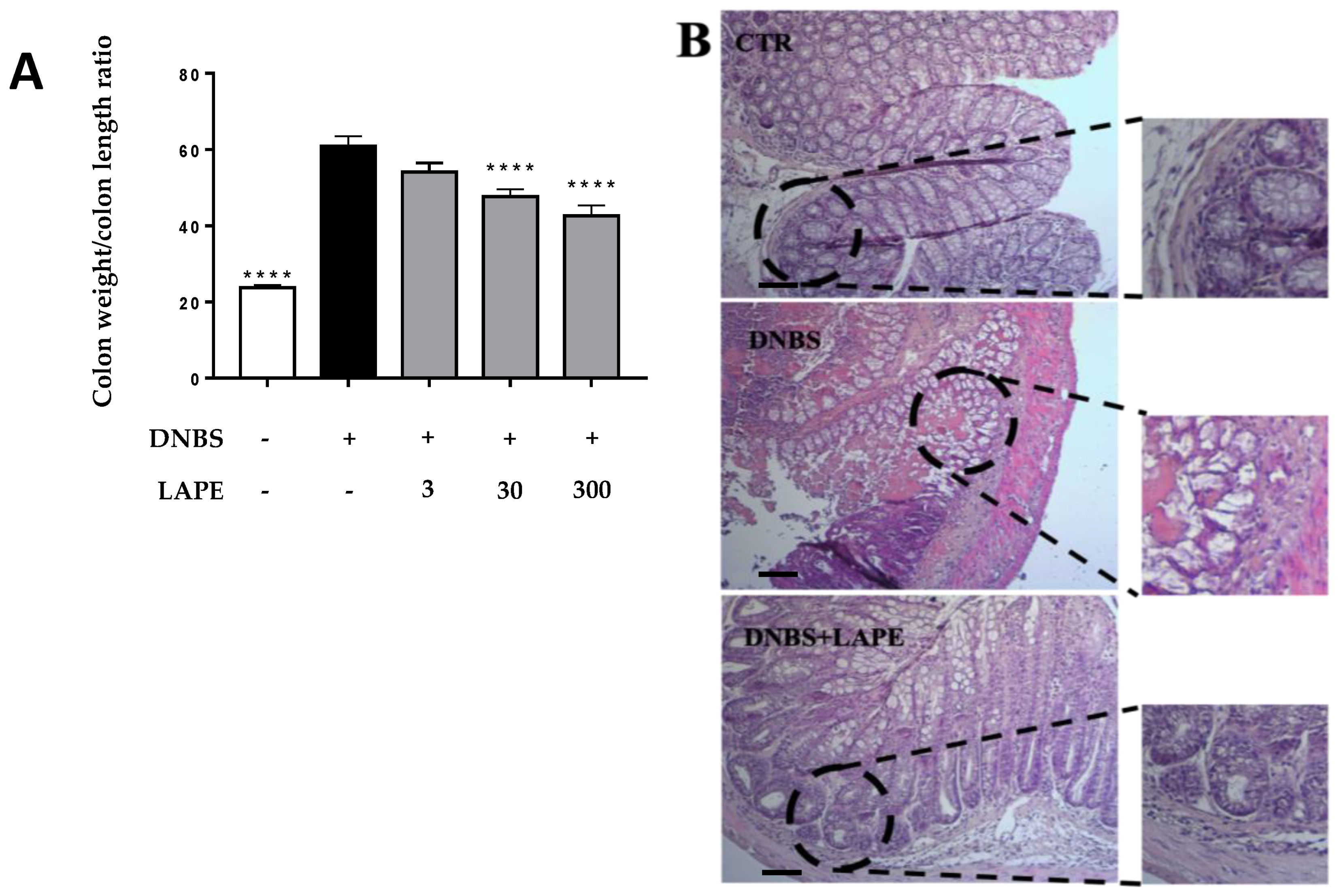
In this study, we demonstrated that a polyphenol extract of Limoncella apple with a high content of chlorogenic acid, catechin and epicatechin ameliorates the DNBS-induced intestinal and liver inflammation. In mice treated with DNBS, we have shown that oral administration of LAPE reduced the histological damage in the inflamed colon, as well as the colon weight/colon length ratio, a macroscopic marker of colitis severity [
16]. The intestinal anti-inflammatory effect of LAPE was further confirmed by its ability to reduce the production of the pro-inflammatory cytokines IL-1β and IL-6 in colon tissues. It is well-established that pro-inflammatory mediators induce neutrophil infiltration, leading to the amplification of the inflammatory response and intestinal injury. Accordingly, LAPE was also able to reduce MPO activity, a known marker of neutrophil infiltration in colitis [
16].
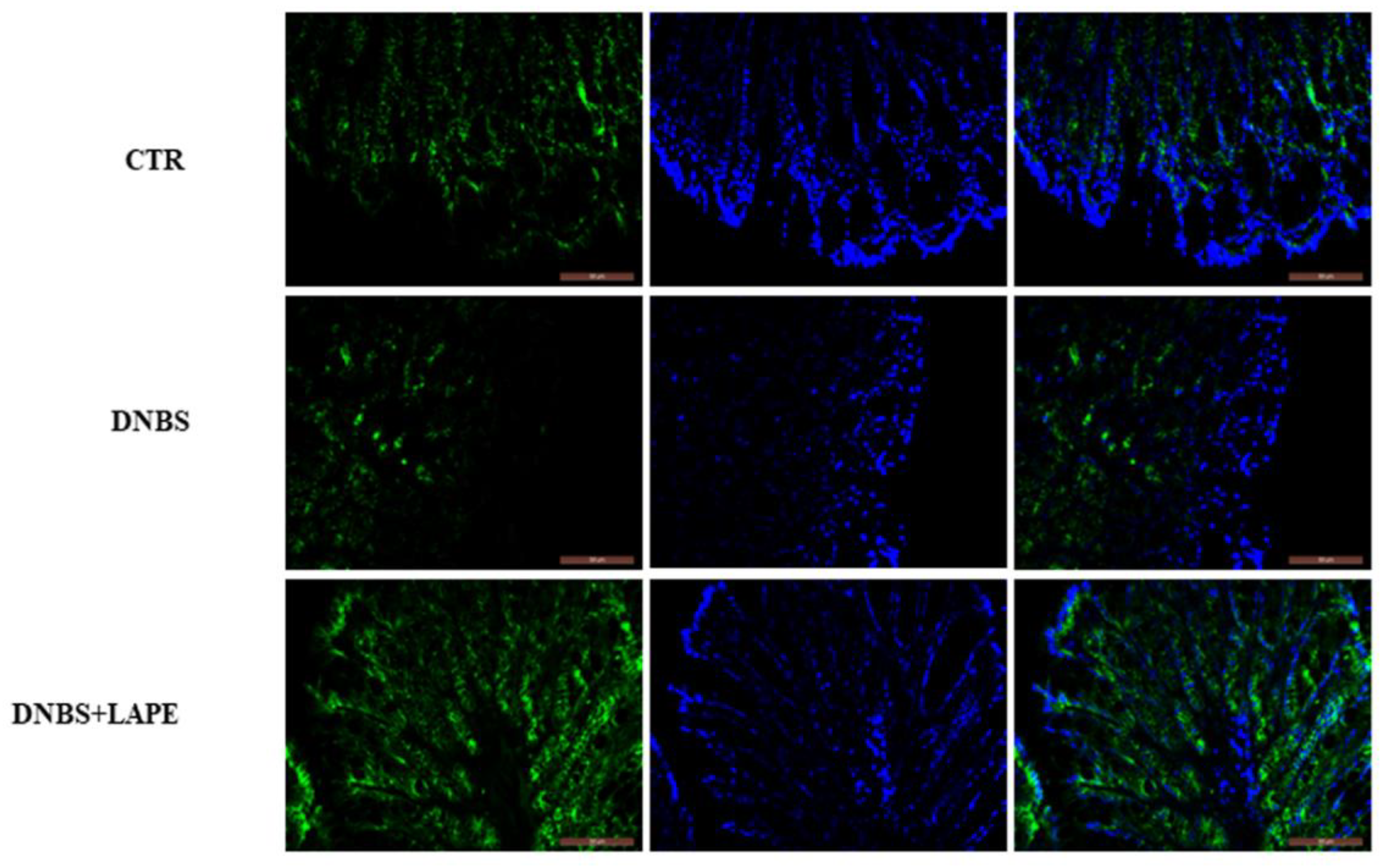
As the dysfunction of the intestinal barrier is a crucial factor in the development of IBD, we assessed the impact of LAPE on the integrity of the intestinal epithelium. In the present study, we demonstrated that oral administration of LAPE restored the DNBS-increased intestinal permeability, as well as increased the β-catenin localization at the colon membrane, maintaining cell–cell adhesion and preventing barrier disruption. β-catenin is a key regulator of intestinal epithelial renewal and the main nuclear effector of canonical Wnt signaling [
13,
24,
25]. Moreover, the inhibition of β-catenin translocation to nucleus after LAPE treatment suggests inactivation of the Wnt pathway, confirming the results previously reported in colonic epithelial cells [
15].
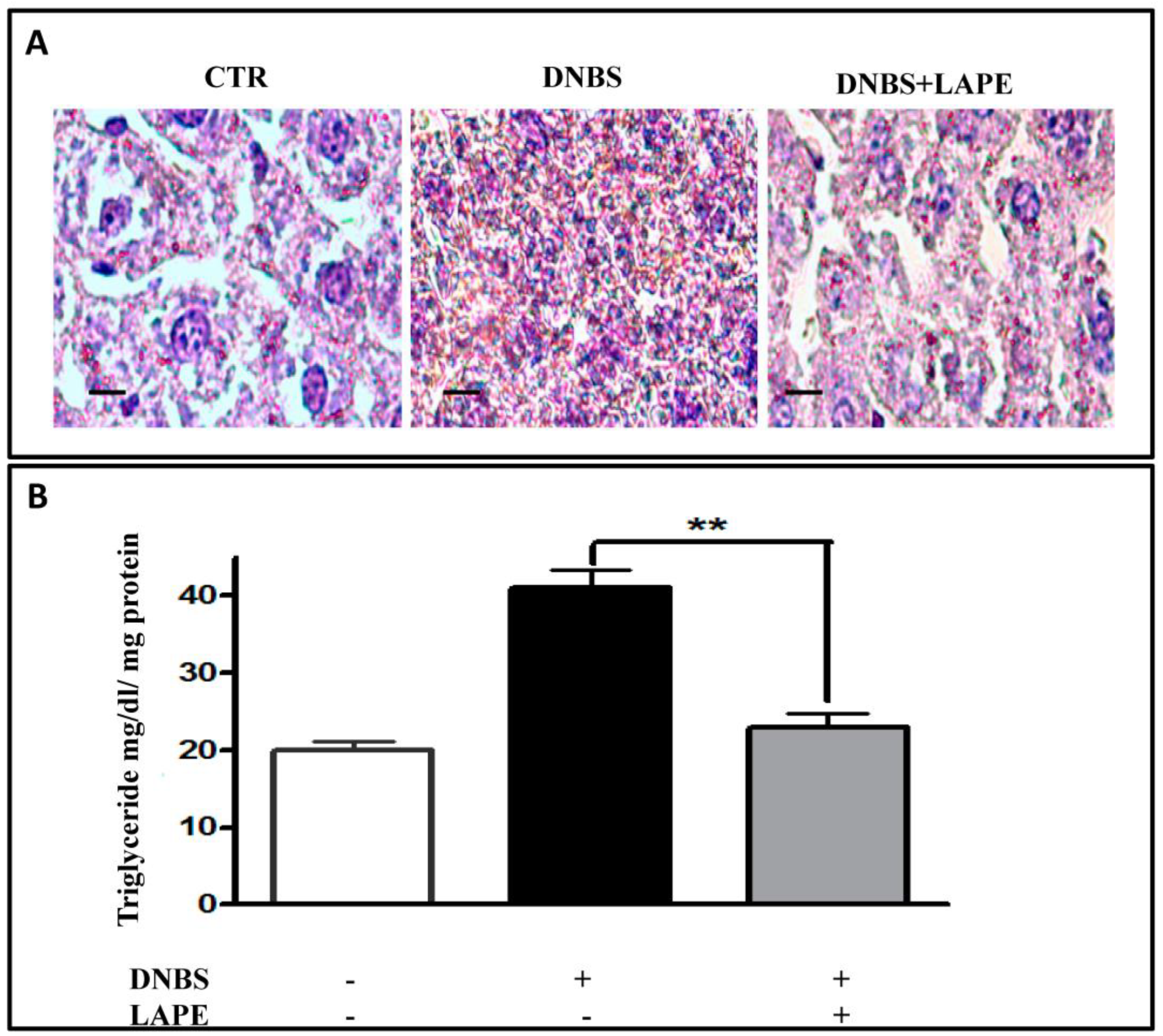
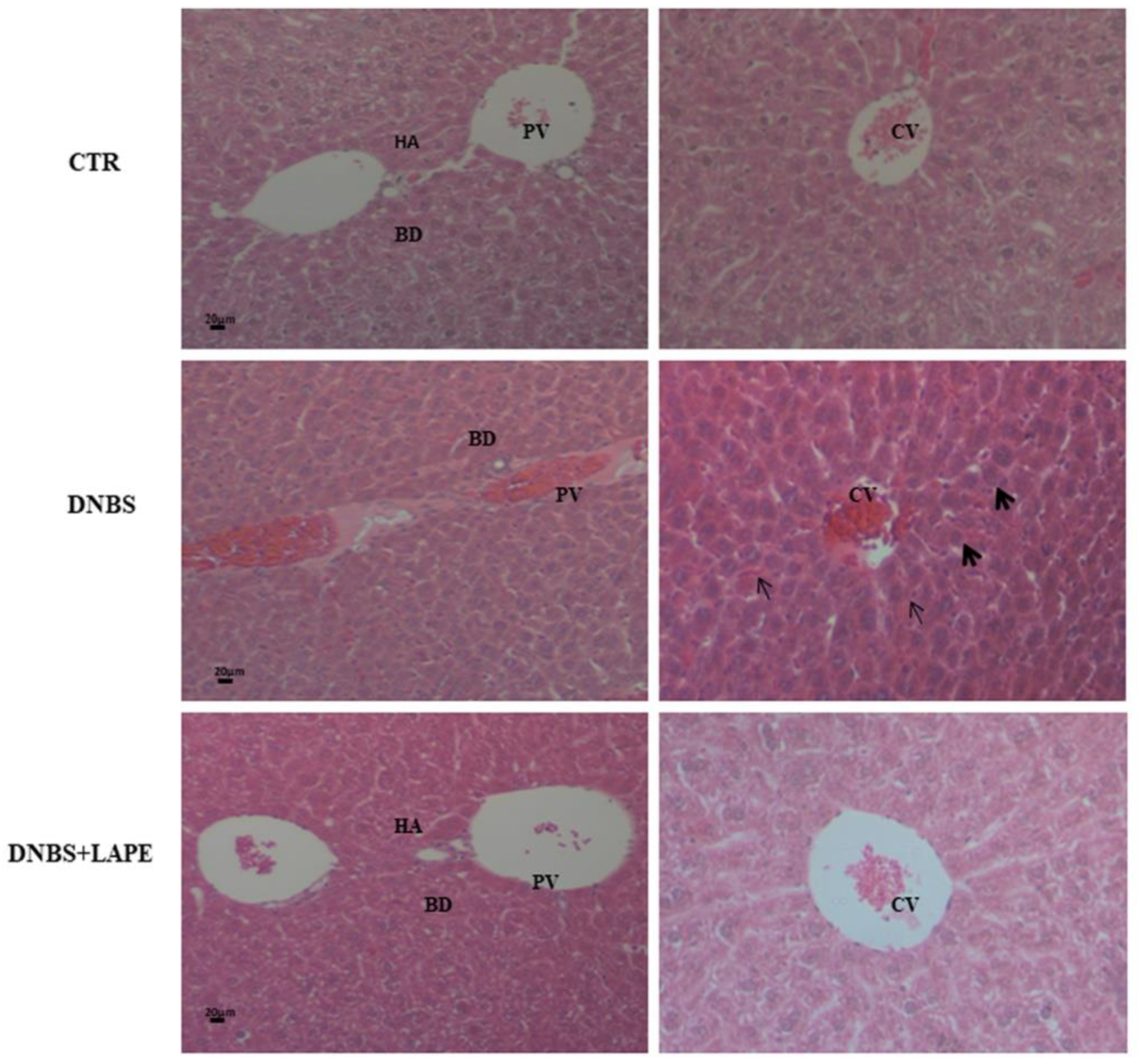
It is well established that hepatobiliary pathologies are the most common extraintestinal manifestations in IBD patients [
26]. Relevant to this point, LAPE administration reduced the histological damage to the liver caused by colitis as well as the accumulation of the lipid in the liver tissue. Furthermore, LAPE ameliorated the hepatic functionality in mice with colitis, as revealed by the reduction in the levels of ALT and AST. Since the Wnt/β-catenin signaling pathway modulates inflammatory and immune responses via NF-κB [
26], we evaluated the effect of LAPE on the expression of this nuclear factor in liver tissues. The NF-κB pathway requires a delicate balance, as too little or too much NF-κB activation may have negative effects on the liver, either by exacerbating liver inflammation or failing to adequately protect it from cell death [
27]. Our study demonstrates that LAPE exerts an anti-inflammatory effect on liver injury by inhibiting NF-κB activation at the concentration of 300 mg/kg. Specifically, this treatment dose for animals is equivalent to a human daily dose of 40.50 g of Limoncella apple (fresh weight). Since this peculiar apple variety produces small and elongated fruits with a reported average weight of 100 g, the theoretically calculated daily dose for humans could be equated to consuming approximately half of a single Limoncella apple per day.
4. Materials and Methods
4.1. Drugs and Reagents
2,4,6-dinitrobenzenesulfonic acid (DNBS), myeloperoxidase (MPO), fluorescein isothiocyanate (FITC)-conjugated dextran (molecular mass 3–5 kDa), 4′,6-Diamidino-2-phenylindole (DAPI) and bovine serum albumin (BSA) were acquired from Sigma (Milan, Italy). Phosphate-buffered saline (PBS) was purchased from Lonza (Milan, Italy). All buffers and solutions were prepared with ultra-high-quality water. All reagents were of the purest commercial grade. All chemicals and reagents employed in this study were of analytical grade.
4.2. Limoncella Apple Polyphenol Extract (LAPE) Preparation
Apples of Malus domestica B. cv. ‘Limoncella’ were gathered in October, after harvesting, in Castelvetere del Calore (Avellino, Italy, Coordinates: 40°55′47″ N 14°59′13″ E). To prepare Limoncella apple extract, lyophilized peels and flesh (10 g) from Limoncella apple samples underwent treatment with 60 mL of 80% methanol (0.5% formic acid). The mixture was homogenized for 5 min using an ultra-turrax (T25-digital, IKA, Staufen im Breisgau, Berlin, Germany) and agitated on an orbital shaker (Sko-DXL, Argolab, Carpy, Italy) at 300 rpm for 15 min. Subsequently, the samples were subjected to an ultrasonic bath treatment for an additional 10 min before being centrifuged at 4427× g for 10 min. The resulting supernatants were collected and stored in darkness at 4 °C. The obtained pellets underwent a second extraction process following the same procedure with an additional 40 mL of the solvent mixture. Finally, the extracts were vacuum filtered, the methanol fraction was removed through evaporation, and the water fraction was lyophilized. The extracts were stored at −20 °C until analysis.
4.3. HPLC/DAD Analysis
Analyses of Limoncella apple peel extract (LAPE) were conducted using a Jasco ExtremaLC-4000 system (Jasco Inc., Easton, MD, USA) equipped with a photodiode array detector (DAD). The chosen column was a Kinetex® C18 column (250 mm × 4.6 mm, 5 μm; Phenomenex, Torrance, CA, USA). The analyses were carried out at a flow rate of 1 mL/min, using solvent A (2% formic acid) and solvent B (0.5% formic acid in acetonitrile and water 50:50, v/v). After a 5 min hold at 10% solvent B, elution proceeded with the following conditions: a gradient from 10% (B) to 55% (B) over 50 min, followed by an increase to 95% (B) in 10 min, with a 5 min maintenance period. Flavanols, procyanidins, dihydrochalcones and hydroxycinnamic acids were acquired at 280 nm, while flavan-3-ols were recorded at 360 nm. For quantitative determination, standard curves were established for each polyphenol standard across a concentration range of 0.1–1.0 μg/μL, with six different concentration levels and duplicate injections at each level. The identity of polyphenols was confirmed by the comparison of the retention time with analytical standards from Sigma-Aldrich Chemical Co. (St. Louis, MO, USA).
4.4. Animals
Male CD1 mice (25–30 g, 6 to 8 weeks old) were supplied from Charles River Laboratories (Calco, Lecco, Italy) and housed at the animal house of the Department of Pharmacy (University of Naples Federico II) in polycarbonate cages under controlled temperature (23 ± 2 °C), constant humidity (60%) and with a 12 h light and 12 h dark cycle. Mice were acclimatized for 1 week under standard environment conditions, with free access to tap water, and fed ad libitum on a standard rodent diet. All mice were fasted overnight before the intracolonic injection of DNBS and for 2 h before the oral gavage of LAPE. All experimental procedures and protocols were approved by the Institutional Animal Ethics Committee and the Italian Ministry of Health (protocol number 444/2018) and were carried out in accordance with the Italian D.L. no. 116 of 27 January 1992 and associated guidelines in the European Communities Council (86/609/ECC and 2010/63/UE). All studies involving mice are reported in compliance with the ARRIVE guidelines for the use of experimental animals [
16].
4.5. DNBS-Induced Experimental Colitis and Pharmacological Treatments
Colitis was induced by intrarectal administration of DNBS (150 mg/kg) by a polyethylene catheter (1 mm in diameter) inserted approximately 4.5 cm proximal to the anus. DNBS was dissolved in 50% (v/v) ethanol/water solution (150 μL/mouse). The DNBS dose was selected based on our preliminary experiments, showing remarkable colonic damage associated with high reproducibility and low mortality. All mice were euthanized by asphyxiation with CO2 3 days after the administration of DNBS, when intestinal inflammation can be easily assessed. Then, the mice’s abdomens were opened by a midline incision, and the colons were removed, isolated from surrounding tissues, their length measured, rinsed, weighed, and processed as required. Mice were randomly allocated to 5 experimental groups as follows: control (vehicle), DNBS, DNBS+LAPE 3 mg/kg, DNBS+LAPE 30 mg/kg and DNBS+LAPE 300 mg/kg (10–12 mice for each experimental group). Outcome assessments were performed in a single-blind manner. The vehicle (water) or LAPE were given by oral gavage (3–300 mg/kg) once a day for 3 consecutive days starting 24 h after the colitis induction. The last administration of vehicle or LAPE was given 2 h before euthanasia. The colon weight/colon length ratio (mg/cm) was used as a macroscopic marker of intestinal inflammation. For biochemistry analysis, tissues were snap-frozen and kept at −80 °C until use, while for histopathological analysis, samples were fixed in 10% formaldehyde.
4.6. In Vivo Intestinal Permeability
Intestinal permeability was evaluated using the FITC-labeled dextran method in mice, involving blood collection through cardiac puncture [
19]. In brief, two days after administering DNBS, mice received 600 mg/kg body weight of fluorescein isothiocyanate (FITC)-conjugated dextran (molecular mass 3–5 kDa) via oral gavage. One day later, mice were euthanized using CO
2 asphyxiation, blood was collected through cardiac puncture and the serum was analyzed for FITC-derived fluorescence using a fluorescent microplate reader with excitation–emission wavelengths of 485–520 nm (LS55 Luminescence Spectrometer, PerkinElmer Instruments, Shanghai, China). A standard curve was generated using serial dilutions of FITC-dextran in tap water, and then the fluorescein fluorescence in samples and standards was measured. Intestinal permeability was quantified as FITC (μM) detected in the mice serum.
4.7. Measurements of Myeloperoxidase (MPO) Activity
MPO activity, a peroxidase enzyme used to measure the neutrophil infiltration in inflamed colons, was assessed as previously described. Briefly, full-thickness colons were mechanically homogenized in a lysis buffer (0.5% hexadecyltrimethylammonium bromide in 3-(N morpholino) propane sulfonic acid 10 mM) in the ratio of 50 mg tissue/mL buffer. The homogenates were centrifuged for 20 min at 15,000× g at 4 °C and the supernatant was incubated with a sodium phosphate buffer (NaPP pH 5.5) and tetra-methylbenzidine 16 mM. After 5 min, 1 M hydrogen peroxide was added, then the reaction was stopped with 2 M acetic acid. The rate of MPO activity was measured by using a spectrophotometer cuvette (650 nm). Different dilutions of human MPO enzyme of known concentrations were used to obtain a standard curve. MPO activity was calculated using standard curve interpolation and expressed as U/mg of colon tissue.
4.8. Measurement of Colonic Cytokine Production
Interleukin (IL)-1β, IL-6 and IL-10 levels were measured in the colon homogenate supernatants obtained from all experimental groups by using commercial enzyme-linked immunosorbent assay (ELISA) kits (Invitrogen, Waltham, MA, USA) according to the manufacturer’s instructions. Briefly, full-thickness colons were homogenized in a lysis buffer containing 0.5 M b-glycerophosphate, 20 mM MgCl2, 10 mM ethylene glycol tetraacetic acid and supplemented with 100 mM dithiothreitol and protease/phosphatase inhibitors (100 mM dimethylsulphonyl fluoride, 2 mg/mL apronitin, 2 mM leupeptin and 10 mM Na3VO4). Homogenates were centrifuged at 600× g for 10 min at 4 °C, then the supernatants were collected, centrifuged at 15,000× g for 10 min at 4 °C and used for cytokine detection. The absorbance was measured at 490 nm and the results were expressed as pg/mL of protein extract.
4.9. Histological Analysis
Paraffin-embedded colon and liver tissues from mice were cut into 5 mm sections. The sections were stained with hematoxylin-eosin (H&E), according to the protocol previously described [
21]. After dehydration, the sections were examined under a light microscope (Leica DM 2500, Leica Microsystems, Wetzlar, Germany). Photographs were taken using a Leica DFC320 R2 digital camera.
4.10. Serum ALT and AST and Lipid Peroxidation Analysis
To measure the biochemical markers of hepatic injury, the serum levels of alanine transaminase (ALT) and aspartate transaminase (AST) were analyzed by colorimetric methods. All measurements were performed using commercially available diagnostic kits for AST/ALT activity (cat. N. MAK055/MAK052, Sigma-Aldrich, St. Louis, MO, USA). Further, the serum lipid peroxidation was evaluated by the thiobarbituric acid reactive substance (TBARS) assay. Briefly, sera samples were incubated with 0.5 mL of aqueous solution of thiobarbituric acid and acetic acid at pH 3.5. After heating at 95 °C for 45 min, the samples were centrifuged at 4000 rpm for 5 min. In the supernatant fractions, TBARS were quantified by spectrophotometry at 532 nm. Results were expressed as TBARS μM/μg of serum protein. Each data point is the average of triplicate measurements, with each individual experiment performed in duplicate.
4.11. Immuno-Fluorescence Analysis
The fixed samples (colon and liver) were dewaxed, re-hydrated and processed. Briefly, antigen retrieval was carried out by pressure-cooking slides for 3 min in a 0.01 M citrate buffer (pH 6.0). To avoid nonspecific interactions of antibodies, the slides were treated for 3 h in 5% BSA in PBS. Immunostaining of colon tissue was performed by overnight incubation at 4 °C, with an anti-β-catenin antibody (1:500, Alexa Fluor
®, BD Pharmingen
TM, Franklin Lakes, NJ, USA). Immunostaining of liver tissue was performed by overnight incubation at 4 °C with an anti-p50 NF-kB antibody (1:100, Santa Cruz Biotechnology, Dallas, TX, USA). After washing in PBS, the slides were incubated for 1 h with a secondary 633-conjugated antibody (1:500 Alexa Fluor, Invitrogen). The slides were mounted on microscope slides using Mowiol + DAPI for nuclear staining and then observed under the optical microscope (Leica DM 5000 B + CTR 5000) [
17].
4.12. Statistical Analysis
All results were expressed as means ± standard error of the mean (SEM) of at least three independent experiments. Statistical analysis was performed using GraphPad Prism version 8. All data were compared using a one-way analysis of variance (ANOVA), followed by Dunnett’s multiple comparison test. The symbol “n” refers to the number of samples for each set of experiments. Statistically significant differences were accepted when p was <0.05.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}