

Damage to the Locus Coeruleus Alters the Expression of Key Proteins in Limbic Neurodegeneration
 , ,
, ,
Abstract
:1. Introduction
2. Results
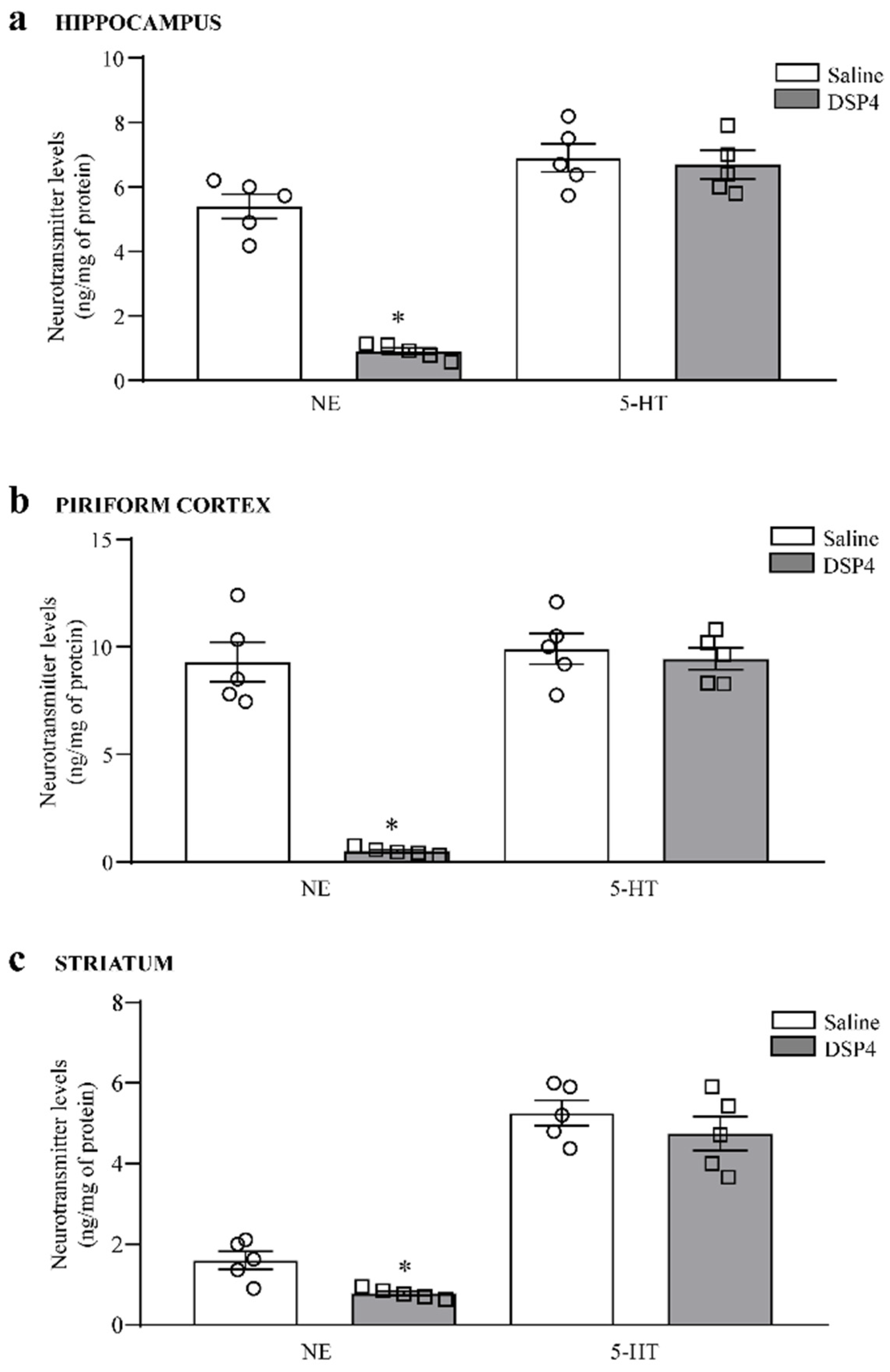
2.1. DSP4 Induces Specific and Severe Loss of NE Terminals within Limbic Regions, Leaving Unmodified 5-HT Levels
2.2. DSP4 Induces a Dramatic Loss of HSP70-Stained Cells within Limbic Regions
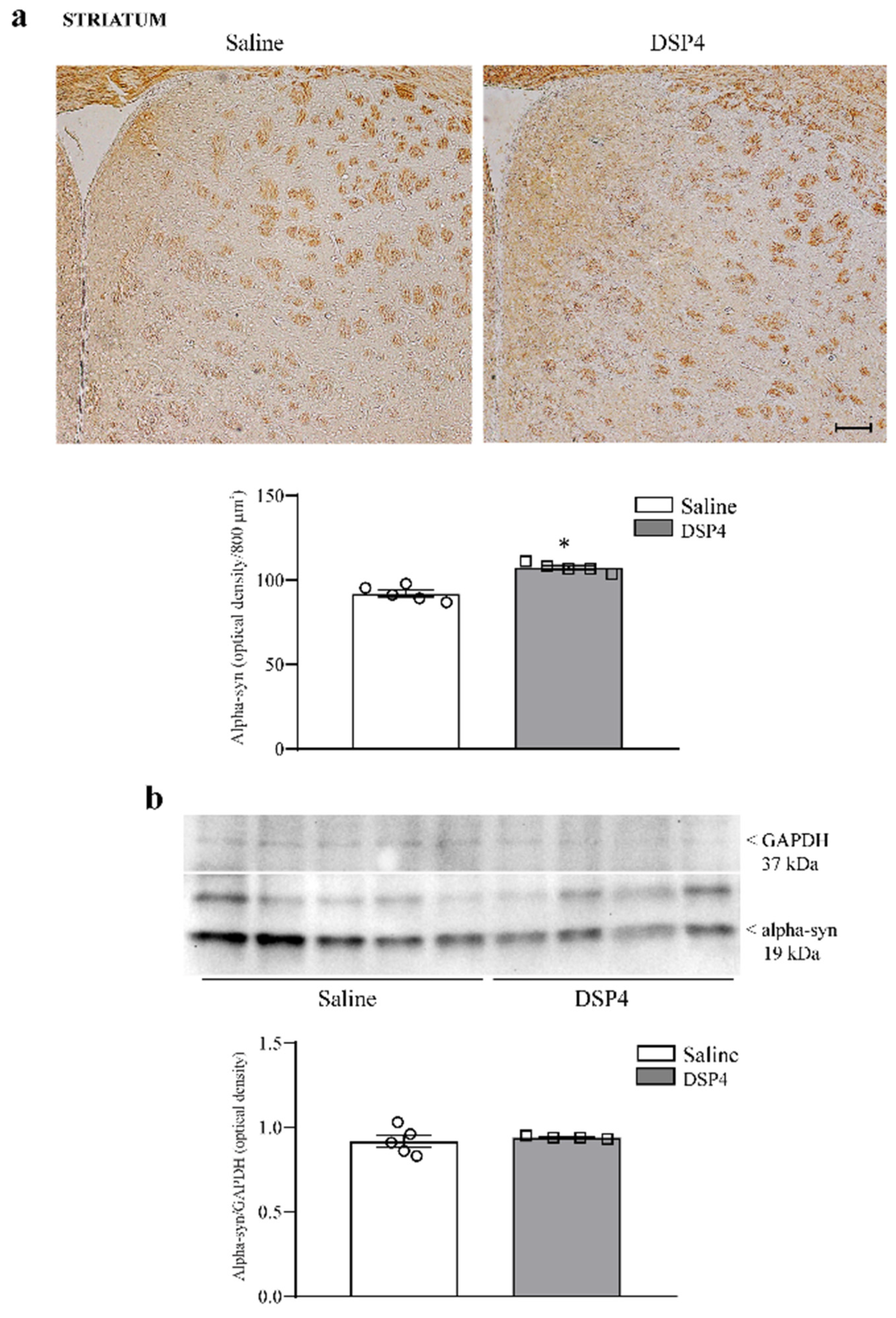
2.3. DSP4 Elevates Alpha-Syn within Limbic Regions
2.4. DSP4 Elevates p-Tau within Limbic Regions
2.5. DSP4 Induces a Loss of p62 within Limbic Regions and the Dorsal Striatum
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Design
4.3. Tissue Sampling
4.4. Histochemistry
4.5. Immunohistochemistry
4.6. SDS Page Immunoblotting
4.7. HPLC Assay
4.8. Open-Field Test
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bucci, D.; Busceti, C.L.; Calierno, M.T.; Di Pietro, P.; Madonna, M.; Biagioni, F.; Ryskalin, L.; Limanaqi, F.; Nicoletti, F.; Fornai, F. Systematic Morphometry of Catecholamine Nuclei in the Brainstem. Front. Neuroanat. 2017, 11, 98. [Google Scholar] [CrossRef]
- Cirelli, C.; Pompeiano, M.; Tononi, G. Neuronal gene expression in the waking state: A role for the locus coeruleus. Science 1996, 274, 1211–1215. [Google Scholar] [CrossRef]
- Bang, D.; Luo, Y.; Barbosa, L.S.; Batten, S.R.; Hadj-Amar, B.; Twomey, T.; Melville, N.; White, J.P.; Torres, A.; Celaya, X.; et al. Noradrenaline tracks emotional modulation of attention in human amygdala. Curr. Biol. 2023, 33, 5003–5010. [Google Scholar] [CrossRef]
- Le, L.; Feidler, A.M.; Li, H.; Kara-Pabani, K.; Lamantia, C.; O’Banion, M.K.; Majewska, K.A. Noradrenergic signaling controls Alzheimer’s disease pathology via activation of microglial β2 adrenergic receptors. bioRxiv 2023. [Google Scholar] [CrossRef]
- Grella, S.L.; Donaldson, T.N. Contextual memory engrams, and the neuromodulatory influence of the locus coeruleus. Front. Mol. Neurosci. 2024, 17, 1342622. [Google Scholar] [CrossRef]
- Vanneste, S. Let’s Shape Learning Into Lasting Memories. Neurosci. Insights 2024, 19, 26331055241227220. [Google Scholar] [CrossRef] [PubMed]
- Markussen, N.B.; Knopper, R.W.; Hasselholt, S.; Skoven, C.S.; Nyengaard, J.R.; Østergaard, L.; Hansen, B. Locus coeruleus ablation in mice: Protocol optimization, stereology and behavioral impact. Front. Cell Neurosci. 2023, 17, 1138624. [Google Scholar] [CrossRef] [PubMed]
- Pace, S.A.; Myers, B. Hindbrain Adrenergic/Noradrenergic Control of Integrated Endocrine and Autonomic Stress Responses. Endocrinology 2023, 165, bqad178. [Google Scholar] [CrossRef] [PubMed]
- Fortin, S.M.; Chen, J.C.; Petticord, M.C.; Ragozzino, F.J.; Peters, J.H.; Hayes, M.R. The locus coeruleus contributes to the anorectic, nausea, and autonomic physiological effects of glucagon-like peptide-1. Sci. Adv. 2023, 9, eadh0980. [Google Scholar] [CrossRef]
- Lui, S.; Brink, A.K.; Corbit, L.H. Optogenetic stimulation of the locus coeruleus enhances appetitive extinction in rats. elife 2024, 12, RP89267. [Google Scholar] [CrossRef] [PubMed]
- Mann, D.M. The locus coeruleus and its possible role in ageing and degenerative disease of the human central nervous system. Mech. Ageing Dev. 1983, 23, 73–94. [Google Scholar] [CrossRef]
- German, D.C.; Manaye, K.F.; White, C.L., 3rd; Woodward, D.J.; McIntire, D.D.; Smith, W.K.; Kalaria, R.N.; Mann, D.M. Disease-specific patterns of locus coeruleus cell loss. Ann. Neurol. 1992, 32, 667–676. [Google Scholar] [CrossRef]
- Gesi, M.; Soldani, P.; Giorgi, F.S.; Santinami, A.; Bonaccorsi, I.; Fornai, F. The role of the locus coeruleus in the development of Parkinson’s disease. Neurosci. Biobehav. Rev. 2000, 24, 655–668. [Google Scholar] [CrossRef]
- Marien, M.R.; Colpaert, F.C.; Rosenquist, A.C. Noradrenergic mechanisms in neurodegenerative diseases: A theory. Brain Res. Brain Res. Rev. 2004, 45, 38–78. [Google Scholar] [CrossRef] [PubMed]
- Schuerger, R.J.; Balaban, C.D. N-(2-chloroethyl)-N-ethyl-2-bromobenzylamine (DSP-4) has differential efficacy for causing central noradrenergic lesions in two different rat strains: Comparison between Long-Evans and Sprague-Dawley rats. J. Neurosci. Methods 1995, 58, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Rodriguez, E.; Vegas-Suarez, S.; Morera-Herreras, T.; De Deurwaerdere, P.; Miguelez, C. The Noradrenergic System in Parkinson’s Disease. Front. Pharmacol. 2020, 11, 435. [Google Scholar] [CrossRef] [PubMed]
- Tamano, H.; Ishikawa, Y.; Shioya, A.; Itoh, R.; Oneta, N.; Shimaya, R.; Egawa, M.; Adlard, P.A.; Bush, A.I.; Takeda, A. Adrenergic β receptor activation reduces amyloid β1-42-mediated intracellular Zn2+ toxicity in dentate granule cells followed by rescuing impair-ment of dentate gyrus LTP. Neurotoxicology 2020, 79, 177–183. [Google Scholar] [CrossRef]
- Jacobs, H.I.L.; Becker, J.A.; Kwong, K.; Engels-Domínguez, N.; Prokopiou, P.C.; Papp, K.V.; Properzi, M.; Hampton, O.L.; d’Oleire Uquillas, F.; Sanchez, J.S.; et al. In vivo and neuropa-thology data support locus coeruleus integrity as indicator of Alzheimer’s disease pathology and cognitive decline. Sci. Transl. Med. 2021, 13, eabj2511. [Google Scholar] [CrossRef] [PubMed]
- Kjelvik, G.; Evensmoen, H.R.; Hummel, T.; Engedal, K.; Selbæk, G.; Saltvedt, I.; Håberg, A.K. The Human Brain Representation of Odor Identification in Amnestic Mild Cognitive Impairment and Alzheimer’s Dementia of Mild Degree. Front. Neurol. 2021, 11, 607566. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.E. The mechanistic link between selective vulnerability of the locus coeruleus and neurodegeneration in Alzheimer’s disease. Acta Neuropathol. 2021, 141, 631–650. [Google Scholar] [CrossRef]
- Cassidy, C.M.; Therriault, J.; Pascoal, T.A.; Cheung, V.; Savard, M.; Tuominen, L.; Chamoun, M.; McCall, A.; Celebi, S.; Lussier, F.; et al. Association of locus coeruleus integrity with Braak stage and neuropsychiatric symptom severity in Alzheimer’s disease. Neuropsychopharmacology 2022, 47, 1128–1136. [Google Scholar] [CrossRef]
- Gilvesy, A.; Husen, E.; Magloczky, Z.; Mihaly, O.; Hortobágyi, T.; Kanatani, S.; Heinsen, H.; Renier, N.; Hökfelt, T.; Mulder, J.; et al. Spatiotemporal characterization of cellular tau pathology in the human locus coeruleus-pericoerulear complex by three-dimensional imaging. Acta Neuropathol. 2022, 144, 651–676. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, I.L.; Dello Russo, C.; Novellino, F.; Caso, J.R.; García-Bueno, B.; Leza, J.C.; Madrigal, J.L.M. Noradrenaline in Alzheimer’s Disease: A New Potential Therapeutic Target. Int. J. Mol. Sci. 2022, 23, 6143. [Google Scholar] [CrossRef] [PubMed]
- Malatt, C.; Tagliati, M. The role of the locus coeruleus/norepinephrine system in the pathogenesis of neurodegenerative disorders: An update. Curr. Opin. Neurol. 2022, 35, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Sibahi, A.; Gandhi, R.; Al-Haddad, R.; Therriault, J.; Pascoal, T.; Chamoun, M.; Boutin-Miller, K.; Tardif, C.; Rosa-Neto, P.; Cassidy, C.M. Characterization of an automated method to segment the human locus coeruleus. Hum. Brain Mapp. 2023, 44, 3913–3925. [Google Scholar] [CrossRef]
- Szot, P.; Weinshenker, D.; White, S.S.; Robbins, C.A.; Rust, N.C.; Schwartzkroin, P.A.; Palmiter, R.D. Norepinephrine-deficient mice have increased susceptibility to seizure-inducing stimuli. J. Neurosci. 1999, 19, 10985–10992. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, F.S.; Ferrucci, M.; Lazzeri, G.; Pizzanelli, C.; Lenzi, P.; Alessandrì, M.G.; Murri, L.; Fornai, F. A damage to locus coeruleus neurons converts sporadic seizures into self-sustaining limbic status epilepticus. Eur. J. Neurosci. 2003, 17, 2593–2601. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, F.S.; Pizzanelli, C.; Biagioni, F.; Murri, L.; Fornai, F. The role of norepinephrine in epilepsy: From the bench to the bedside. Neurosci. Biobehav. Rev. 2004, 28, 507–524. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, F.S.; Mauceli, G.; Blandini, F.; Ruggieri, S.; Paparelli, A.; Murri, L.; Fornai, F. Locus coeruleus and neuronal plasticity in a model of focal limbic epilepsy. Epilepsia 2006, 47, 21–25. [Google Scholar] [CrossRef]
- Pizzanelli, C.; Lazzeri, G.; Fulceri, F.; Giorgi, F.S.; Pasquali, L.; Cifelli, G.; Murri, L.; Fornai, F. Lack of alpha 1b-adrenergic receptor protects against epileptic seizures. Epilepsia 2009, 50, S59–S64. [Google Scholar] [CrossRef]
- Fornai, F.; Ruffoli, R.; Giorgi, F.S.; Paparelli, A. The role of locus coeruleus in the antiepileptic activity induced by vagus nerve stimulation. Eur. J. Neurosci. 2011, 33, 2169–2178. [Google Scholar] [CrossRef] [PubMed]
- Biagioni, F.; Celli, R.; Puglisi-Allegra, S.; Nicoletti, F.; Giorgi, F.S.; Fornai, F. Noradrenaline and Seizures: A Perspective on the Role of Adrenergic Receptors in Limbic Seizures. Curr. Neuropharmacol. 2023, 21, 2233–2236. [Google Scholar] [CrossRef] [PubMed]
- Larsen, L.E.; Caestecker, S.; Stevens, L.; van Mierlo, P.; Carrette, E.; Boon, P.; Vonck, K.; Raedt, R. Hippocampal seizures differentially modulate locus coeruleus activity and result in consistent time-locked release of noradrenaline in rat hippocampus. Neurobiol. Dis. 2023, 189, 106355. [Google Scholar] [CrossRef] [PubMed]
- Nishino, K.; Lin, C.S.; Morse, J.K.; Davis, J.N. DSP4 treatment worsens hippocampal pyramidal cell damage after transient ischemia. Neuroscience 1991, 43, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Hu, D.X.; He, F.; Li, C.Y.; Qi, G.J.; Cai, H.W.; Li, T.X.; Ming, J.; Zhang, P.; Chen, X.Q.; et al. Locus coeruleus-CA1 projections are involved in chronic depressive stress-induced hippocampal vulnerability to transient global ischaemia. Nat. Commun. 2019, 10, 2942. [Google Scholar] [CrossRef] [PubMed]
- Sternberg, Z.; Schaller, B. Central Noradrenergic Agonists in the Treatment of Ischemic Stroke-an Overview. Transl. Stroke Res. 2020, 11, 165–184. [Google Scholar] [CrossRef] [PubMed]
- Marien, M.; Briley, M.; Colpaert, F. Noradrenaline depletion exacerbates MPTP-induced striatal dopamine loss in mice. Eur. J. Pharmacol. 1993, 236, 487–489. [Google Scholar] [CrossRef]
- Fornai, F.; Alessandrì, M.G.; Fascetti, F.; Vaglini, F.; Corsini, G.U. Clonidine suppresses 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced reductions of striatal dopamine and tyrosine hydroxylase activity in mice. J. Neurochem. 1995, 65, 704–709. [Google Scholar] [CrossRef]
- Fornai, F.; Alessandrì, M.G.; Torracca, M.T.; Bassi, L.; Corsini, G.U. Effects of noradrenergic lesions on MPTP/MPP+ kinetics and MPTP-induced nigrostriatal dopamine depletions. J. Pharmacol. Exp. Ther. 1997, 283, 100–107. [Google Scholar]
- Mavridis, M.; Degryse, A.D.; Lategan, A.J.; Marien, M.R.; Colpaert, F.C. Effects of locus coeruleus lesions on parkinsonian signs, striatal dopamine and substantia nigra cell loss after 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine in monkeys: A possible role for the locus coeruleus in the progression of Parkinson’s disease. Neuroscience 1991, 41, 507–523. [Google Scholar] [CrossRef]
- Chan-Palay, V.; Asan, E. Alterations in catecholamine neurons of the locus coeruleus in senile dementia of the Alzheimer type and in Parkinson’s disease with and without dementia and depression. J. Comp. Neurol. 1989, 287, 373–392. [Google Scholar] [CrossRef] [PubMed]
- Rajput, A.H.; Uitti, R.J.; Sudhakar, S.; Rozdilsky, B. Parkinsonism and neurofibrillary tangle pathology in pigmented nuclei. Ann. Neurol. 1989, 25, 602–606. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Hornykiewicz, O.; Kish, S.J. Inverse relationship between brain noradrenaline level and dopamine loss in Parkinson disease: A possible neuroprotective role for noradrenaline. Arch. Neurol. 2006, 63, 1724–1728. [Google Scholar] [CrossRef] [PubMed]
- Terry, R.D. Morphological changes in Alzheimer’s disease-senile dementia: Ultrastructural changes and quantitative studies. Res. Publ. Assoc. Res. Nerv. Ment. Dis. 1979, 57, 99–105. [Google Scholar] [PubMed]
- Mann, D.M.; Lincoln, J.; Yates, P.O.; Stamp, J.E.; Toper, S. Changes in the monoamine containing neurones of the human CNS in senile dementia. Br. J. Psychiatry 1980, 136, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Bondareff, W.; Mountjoy, C.Q.; Roth, M. Selective loss of neurones of origin of adrenergic projection to cerebral cortex (nucleus locus coeruleus) in senile dementia. Lancet 1981, 1, 783–784. [Google Scholar] [CrossRef] [PubMed]
- Forno, L.S.; Norville, R.L. Synaptic morphology in the human locus ceruleus. Acta Neuropathol. 1981, 53, 7–14. [Google Scholar] [CrossRef]
- Tomlinson, B.E.; Irving, D.; Blessed, G. Cell loss in the locus coeruleus in senile dementia of Alzheimer type. J. Neurol. Sci. 1981, 49, 419–428. [Google Scholar] [CrossRef]
- van Dongen, P.A. The human locus coeruleus in neurology and psychiatry. (Parkinson’s, Lewy body, Hallervorden-Spatz, Alzheimer’s and Korsakoff’s disease, (pre)senile dementia, schizophrenia, affective disorders, psychosis). Prog. Neurobiol. 1981, 17, 97–139. [Google Scholar] [CrossRef]
- Chan-Palay, V. Alterations in the locus coeruleus in dementias of Alzheimer’s and Parkinson’s disease. Prog. Brain Res. 1991, 88, 625–630. [Google Scholar] [CrossRef]
- Mann, D.M.; Yates, P.O.; Hawkes, J. The noradrenergic system in Alzheimer and multi-infarct dementias. J. Neurol. Neurosurg. Psychiatry 1982, 45, 113–119. [Google Scholar] [CrossRef]
- Marcyniuk, B.; Mann, D.M.; Yates, P.O. Loss of nerve cells from locus coeruleus in Alzheimer’s disease is topographically arranged. Neurosci. Lett. 1986, 64, 247–252. [Google Scholar] [CrossRef]
- Banwinkler, M.; Theis, H.; Prange, S.; van Eimeren, T. Imaging the Limbic System in Parkinson’s Disease-A Review of Limbic Pathology and Clinical Symptoms. Brain Sci. 2022, 12, 1248. [Google Scholar] [CrossRef]
- Jellinger, K.A. Pathobiology of Cognitive Impairment in Parkinson Disease: Challenges and Outlooks. Int. J. Mol. Sci. 2023, 25, 498. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Wang, L.L.; Jiang, X.D.; Chen, D.J.; Huang, T.A.; Ding, W.B. Hippocampal volume and resting-state functional connectivity on magnetic resonance imaging in patients with Parkinson and depression. Quant. Imaging Med. Surg. 2024, 14, 824–836. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, P.; Dong, Q.; Zhao, Z.; Zheng, W.; Zhang, J.; Hu, X.; Yao, Z.; Hu, B. Fine-grained features characterize hippocampal and amygdaloid change pattern in Parkinson’s disease and discriminate cognitive-deficit subtype. CNS Neurosci. Ther. 2024, 30, e14480. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.V.; Gelinas, J.N. Noradrenergic gating of long-lasting synaptic potentiation in the hippocampus: From neurobiology to translational biomedicine. J. Neurogenet. 2018, 32, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Bortolotto, V.; Bondi, H.; Cuccurazzu, B.; Rinaldi, M.; Canonico, P.L.; Grilli, M. Salmeterol, a β2 Adrenergic Agonist, Promotes Adult Hippocampal Neurogenesis in a Region-Specific Manner. Front. Pharmacol. 2019, 10, 1000. [Google Scholar] [CrossRef] [PubMed]
- James, T.; Kula, B.; Choi, S.; Khan, S.S.; Bekar, L.K.; Smith, N.A. Locus coeruleus in memory formation and Alzheimer’s disease. Eur. J. Neurosci. 2021, 54, 6948–6959. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Fisher, D.W.; Rodriguez, G.; Yu, T.; Dong, H. Comparisons of neuroinflammation, microglial activation, and degeneration of the locus coeruleus-norepinephrine system in APP/PS1 and aging mice. J. Neuroinflamm. 2021, 18, 10. [Google Scholar] [CrossRef] [PubMed]
- Goodman, A.M.; Langner, B.M.; Jackson, N.; Alex, C.; McMahon, L.L. Heightened Hippocampal β-Adrenergic Receptor Function Drives Synaptic Potentiation and Supports Learning and Memory in the TgF344-AD Rat Model during Prodromal Alzheimer’s Disease. J. Neurosci. 2021, 41, 5747–5761. [Google Scholar] [CrossRef]
- Titulaer, J.; Björkholm, C.; Feltmann, K.; Malmlöf, T.; Mishra, D.; Bengtsson Gonzales, C.; Schilström, B.; Konradsson-Geuken, Å. The Importance of Ventral Hippocampal Dopamine and Norepinephrine in Recognition Memory. Front. Behav. Neurosci. 2021, 15, 667244. [Google Scholar] [CrossRef]
- Khan, K.M.; Balasubramanian, N.; Gaudencio, G.; Wang, R.; Selvakumar, G.P.; Kolling, L.; Pierson, S.; Tadinada, S.M.; Abel, T.; Hefti, M.; et al. Human tau-overexpressing mice recapitulate brainstem involvement and neuropsychiatric features of early Alzheimer’s disease. Acta Neuropathol. Commun. 2023, 11, 57. [Google Scholar] [CrossRef]
- Terrier, C.; Greco-Vuilloud, J.; Cavelius, M.; Thevenet, M.; Mandairon, N.; Didier, A.; Richard, M. Long-term olfactory enrichment promotes non-olfactory cognition, noradrenergic plasticity and remodeling of brain functional connectivity in older mice. Neurobiol. Aging 2024, 136, 133–156. [Google Scholar] [CrossRef]
- Fallon, J.H.; Moore, R.Y. Catecholamine innervation of the basal forebrain. III. Olfactory bulb, anterior olfactory nuclei, olfactory tubercle and piriform cortex. J. Comp. Neurol. 1978, 180, 533–544. [Google Scholar] [CrossRef]
- Datiche, F.; Cattarelli, M. Catecholamine innervation of the piriform cortex: A tracing and immunohistochemical study in the rat. Brain Res. 1996, 710, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Soto, M.; Casadó-Anguera, V.; Yano, H.; Bender, B.J.; Cai, N.S.; Moreno, E.; Canela, E.I.; Cortés, A.; Meiler, J.; Casadó, V.; et al. α2A- and α2C-Adrenoceptors as Potential Targets for Dopamine and Dopamine Receptor Ligands. Mol. Neurobiol. 2018, 55, 8438–8454. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Tan, H.Y.; Wu, Z.H.; Sun, C.P.; He, J.X.; Li, X.C.; Shao, M. Imaging of olfactory bulb and gray matter volumes in brain areas associated with olfactory function in patients with Parkinson’s disease and multiple system atrophy. Eur. J. Radiol. 2014, 83, 564–570. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.Y.; Eslinger, P.J.; Du, G.; Kong, L.; Lewis, M.M.; Huang, X. Olfactory-related cortical atrophy is associated with olfactory dysfunction in Parkinson’s disease. Mov. Disord. 2014, 29, 1205–1208. [Google Scholar] [CrossRef] [PubMed]
- Sancandi, M.; Schul, E.V.; Economides, G.; Constanti, A.; Mercer, A. Structural Changes Observed in the Piriform Cortex in a Rat Model of Pre-motor Parkinson’s Disease. Front. Cell Neurosci. 2018, 12, 479. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Sun, F.; Sun, W.; Zhang, L.; Wang, Q. Lesion of the Locus Coeruleus Damages Learning and Memory Performance in Paraquat and Maneb-induced Mouse Parkinson’s Disease Model. Neuroscience 2019, 419, 129–140. [Google Scholar] [CrossRef]
- Hummel, T.; Haehner, A.; Thaploo, D.; Georgiopoulos, C.; Falkenburger, B.; Whitcroft, K. Advancement of PD Is Reflected by White Matter Changes in Olfactory Areas: A Pilot Study. Medicina 2021, 57, 1183. [Google Scholar] [CrossRef]
- Roh, H.; Kang, J.; Koh, S.B.; Kim, J.H. Hippocampal volume is related to olfactory impairment in Parkinson’s disease. J. Neuroimaging 2021, 31, 1176–1183. [Google Scholar] [CrossRef]
- Rajani, V.; Yuan, Q. Noradrenergic Modulation of the Piriform Cortex: A Possible Avenue for Understanding Pre-Clinical Alzheimer’s Disease Pathogenesis. Front. Cell Neurosci. 2022, 16, 908758. [Google Scholar] [CrossRef] [PubMed]
- Wylie, K.P.; Kluger, B.M.; Medina, L.D.; Holden, S.K.; Kronberg, E.; Tregellas, J.R.; Buard, I. Hippocampal, basal ganglia and olfactory connectivity contribute to cognitive impairments in Parkinson’s disease. Eur. J. Neurosci. 2023, 57, 511–526. [Google Scholar] [CrossRef] [PubMed]
- Abyadeh, M.; Gupta, V.; Paulo, J.A.; Mahmoudabad, A.G.; Shadfar, S.; Mirshahvaladi, S.; Gupta, V.; Nguyen, C.T.O.; Finkelstein, D.I.; You, Y.; et al. Amyloid-beta and tau protein beyond Alzheimer’s disease. Neural Regen Res. 2024, 19, 1262–1276. [Google Scholar] [CrossRef] [PubMed]
- Citro, S.; Lazzaro, G.D.; Cimmino, A.T.; Giuffrè, G.M.; Marra, C.; Calabresi, P. A multiple hits hypothesis for memory dysfunction in Parkinson disease. Nat. Rev. Neurol. 2024, 20, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Zhang, J.; Zhao, X.; Nie, W.; Xu, X.; Liu, M.; Zhang, X. Olfactory dysfunction and its related molecular mechanisms in Parkinson’s disease. Neural Regen Res. 2024, 19, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Iravani, M.M.; Shoaib, M. Executive dysfunction and cognitive decline, a non-motor symptom of Parkinson’s disease captured in animal models. Int. Rev. Neurobiol. 2024, 174, 231–255. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.P.; Frigerio, I.; Bol, J.G.J.M.; Bouwman, M.M.A.; Wesseling, A.J.; Dahl, M.J.; Rozemuller, A.J.M.; van der Werf, Y.D.; Pouwels, P.J.W.; van de Berg, W.D.J.; et al. Microstructural integrity of the locus coeruleus and its tracts reflect noradrenergic degeneration in Alzheimer’s disease and Parkinson’s disease. Transl. Neurodegener. 2024, 13, 9. [Google Scholar] [CrossRef]
- Huang, M.; Macdonald, J.; Lavenir, I.; Chen, R.; Craxton, M.; Slavik-Smith, E.; Davies, S.W.; Goedert, M. Increase in Tau Pathology in P290S Knock-In Mice Crossed with AppNL-G-F Mice. eNeuro 2022, 9, ENEURO.0247-22.2022. [Google Scholar] [CrossRef]
- Shen, J.; Li, M.; Long, C.; Yang, L.; Jiang, J. Altered Odor-Evoked Electrophysiological Responses in the Anterior Piriform Cortex of Conscious APP/PS1 Mice. J. Alzheimer’s Dis. 2022, 90, 1277–1289. [Google Scholar] [CrossRef]
- Tsui, K.C.; Roy, J.; Chau, S.C.; Wong, K.H.; Shi, L.; Poon, C.H.; Wang, Y.; Strekalova, T.; Aquili, L.; Chang, R.C.; et al. Distribution and inter-regional relationship of amyloid-beta plaque deposition in a 5xFAD mouse model of Alzheimer’s disease. Front Aging Neurosci. 2022, 14, 964336. [Google Scholar] [CrossRef] [PubMed]
- Jobin, B.; Boller, B.; Frasnelli, J.; CIMA-Q group. Smaller grey matter volume in the central olfactory system in mild cognitive impairment. Exp. Gerontol. 2023, 183, 112325. [Google Scholar] [CrossRef]
- Ponnusamy, M.; Wang, S.; Yuksel, M.; Hansen, M.T.; Blazier, D.M.; McMillan, J.D.; Zhang, X.; Dammer, E.B.; Collier, L.; Thinakaran, G. Loss of forebrain BIN1 attenuates hippocampal pathology and neuroinflammation in a tauopathy model. Brain 2023, 146, 1561–1579. [Google Scholar] [CrossRef] [PubMed]
- Steinbart, D.; Yaakub, S.N.; Steinbrenner, M.; Guldin, L.S.; Holtkamp, M.; Keller, S.S.; Weber, B.; Rüber, T.; Heckemann, R.A.; Ilyas-Feldmann, M.; et al. Alzheimer’s Disease Neuroimaging Initiative. Automatic and manual segmentation of the piriform cortex: Method development and validation in patients with temporal lobe epilepsy and Alzheimer’s disease. Hum. Brain Mapp. 2023, 44, 3196–3209. [Google Scholar] [CrossRef]
- Wang, X.; Hu, M.; Xie, Q.; Geng, C.; Jin, C.; Ren, W.; Fan, J.; Ma, T.; Hu, B. Amyloid β oligomers disrupt piriform cortical output via a serotonergic pathway. Neurobiol. Aging 2023, 121, 64–77. [Google Scholar] [CrossRef]
- Silveira-Moriyama, L.; Holton, J.L.; Kingsbury, A.; Ayling, H.; Petrie, A.; Sterlacci, W.; Poewe, W.; Maier, H.; Lees, A.J.; Revesz, T. Regional differences in the severity of Lewy body pathology across the olfactory cortex. Neurosci. Lett. 2009, 453, 77–80. [Google Scholar] [CrossRef]
- Ubeda-Bañon, I.; Saiz-Sanchez, D.; de la Rosa-Prieto, C.; Argandoña-Palacios, L.; Garcia-Muñozguren, S.; Martinez-Marcos, A. alpha-Synucleinopathy in the human olfactory system in Parkinson’s disease: Involvement of calcium-binding protein- and substance P-positive cells. Acta Neuropathol. 2010, 119, 723–735. [Google Scholar] [CrossRef] [PubMed]
- Ubeda-Bañon, I.; Saiz-Sanchez, D.; de la Rosa-Prieto, C.; Martinez-Marcos, A. α-Synuclein in the olfactory system of a mouse model of Parkinson’s disease: Correlation with olfactory projections. Brain Struct. Funct. 2012, 217, 447–458. [Google Scholar] [CrossRef]
- Mason, D.M.; Nouraei, N.; Pant, D.B.; Miner, K.M.; Hutchison, D.F.; Luk, K.C.; Stolz, J.F.; Leak, R.K. Transmission of α-synucleinopathy from olfactory structures deep into the temporal lobe. Mol. Neurodegener. 2016, 11, 49. [Google Scholar] [CrossRef]
- Flores-Cuadrado, A.; Saiz-Sanchez, D.; Mohedano-Moriano, A.; Martinez-Marcos, A.; Ubeda-Bañon, I. Neurodegeneration and contralateral α-synuclein induction after intracerebral α-synuclein injections in the anterior olfactory nucleus of a Parkinson’s disease A53T mouse model. Acta Neuropathol. Commun. 2019, 7, 56. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.E.; Bergkvist, L.; Mercado, G.; Stetzik, L.; Meyerdirk, L.; Wolfrum, E.; Madaj, Z.; Brundin, P.; Wesson, D.W. Deficits in olfactory sensitivity in a mouse model of Parkinson’s disease revealed by plethysmography of odor-evoked sniffing. Sci. Rep. 2020, 10, 9242. [Google Scholar] [CrossRef] [PubMed]
- Kasongo, D.W.; de Leo, G.; Vicario, N.; Leanza, G.; Legname, G. Chronic α-Synuclein Accumulation in Rat Hippocampus Induces Lewy Bodies Formation and Specific Cognitive Impairments. eNeuro 2020, 7, ENEURO.0009-20.2020. [Google Scholar] [CrossRef] [PubMed]
- Bieniek, K.F.; Murray, M.E.; Rutherford, N.J.; Castanedes-Casey, M.; DeJesus-Hernandez, M.; Liesinger, A.M.; Baker, M.C.; Boylan, K.B.; Rademakers, R.; Dickson, D.W. Tau pathology in frontotemporal lobar degeneration with C9ORF72 hexanucleotide repeat expansion. Acta Neuropathol. 2013, 125, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.E.; Bieniek, K.F.; Banks Greenberg, M.; DeJesus-Hernandez, M.; Rutherford, N.J.; van Blitterswijk, M.; Niemantsverdriet, E.; Ash, P.E.; Gendron, T.F.; Kouri, N.; et al. Progressive amnestic dementia, hippocampal sclerosis, and mutation in C9ORF72. Acta Neuropathol. 2013, 126, 545–554. [Google Scholar] [CrossRef]
- Akbar, M.T.; Wells, D.J.; Latchman, D.S.; de Belleroche, J. Heat shock protein 27 shows a distinctive widespread spatial and temporal pattern of induction in CNS glial and neuronal cells compared to heat shock protein 70 and caspase 3 following kainate administration. Brain Res. Mol. Brain Res. 2001, 93, 148–163. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kim, J.Y.; Ko, A.R.; Kang, T.C. Reduction in heat shock protein 90 correlates to neuronal vulnerability in the rat piriform cortex following status epilepticus. Neuroscience 2013, 255, 265–277. [Google Scholar] [CrossRef]
- Posimo, J.M.; Weilnau, J.N.; Gleixner, A.M.; Broeren, M.T.; Weiland, N.L.; Brodsky, J.L.; Wipf, P.; Leak, R.K. Heat shock protein defenses in the neocortex and allocortex of the telencephalon. Neurobiol. Aging 2015, 36, 1924–1937. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Li, T.; Ma, Z.; Zhou, C.; Wang, X.; Li, J. HSPA1A, HSPA2, and HSPA8 Are Potential Molecular Biomarkers for Prognosis among HSP70 Family in Alzheimer’s Disease. Dis. Markers 2022, 2022, 9480398. [Google Scholar] [CrossRef] [PubMed]
- Han, S.M.; Jang, Y.J.; Kim, E.Y.; Park, S.A. The Change in Circadian Rhythms in P301S Transgenic Mice is Linked to Variability in Hsp70-related Tau Disaggregation. Exp. Neurobiol. 2022, 31, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Moll, A.; Ramirez, L.M.; Ninov, M.; Schwarz, J.; Urlaub, H.; Zweckstetter, M. Hsp multichaperone complex buffers pathologically modified Tau. Nat. Commun. 2022, 13, 3668. [Google Scholar] [CrossRef] [PubMed]
- Babazadeh, R.; Schneider, K.L.; Fischbach, A.; Hao, X.; Liu, B.; Nystrom, T. The yeast guanine nucleotide exchange factor Sec7 is a bottleneck in spatial protein quality control and detoxifies neurological disease proteins. Sci. Rep. 2023, 13, 14068. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.L.; Yang, C.C.; Huang, Y.Y.; Chen, Y.A.; Yang, C.W.; Liao, C.Y.; Li, H.; Wu, C.S.; Lin, C.H.; Teng, S.C. The HSP40 family chaperone isoform DNAJB6b prevents neuronal cells from tau aggregation. BMC Biol. 2023, 21, 293. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Xin, J.; Peng, Q.; Li, N.; Sun, S.; Hou, H.; Ma, G.; Wang, N.; Zhang, L.; Tam, K.Y.; et al. UBQLN2 and HSP70 participate in Parkin- mediated mitophagy by facilitating outer mitochondrial membrane rupture. EMBO Rep. 2023, 24, 55859. [Google Scholar] [CrossRef]
- Yamashima, T.; Seike, T.; Mochly-Rosen, D.; Chen, C.H.; Kikuchi, M.; Mizukoshi, E. Implication of the cooking oil-peroxidation product „hydroxynonenal” for Alzheimer’s disease. Front Aging Neurosci. 2023, 15, 1211141. [Google Scholar] [CrossRef]
- Fornai, F.; Bassi, L.; Torracca, M.T.; Alessandrì, M.G.; Scalori, V.; Corsini, G.U. Region-and neurotransmitter-dependent species and strain differences in DSP-4-induced monoamine depletion in rodents. Neurodegeneration 1996, 5, 241–249. [Google Scholar] [CrossRef]
- Duffy, K.B.; Ray, B.; Lahiri, D.K.; Tilmont, E.M.; Tinkler, G.P.; Herbert, R.L.; Greig, N.H.; Ingram, D.K.; Ottinger, M.A.; Mattison, J.A. Effects of Reducing Norepinephrine Levels via DSP4 Treatment on Amyloid-β Pathology in Female Rhesus Macaques (Macaca Mulatta). J. Alzheimer’s Dis. 2019, 68, 115–126. [Google Scholar] [CrossRef]
- Pozzi, N.G.; Bolzoni, F.; Biella, G.E.M.; Pezzoli, G.; Ip, C.W.; Volkmann, J.; Cavallari, P.; Asan, E.; Isaias, I.U. Brain Noradrenergic Innervation Supports the Development of Parkinson’s Tremor: A Study in a Reserpinized Rat Model. Cells 2023, 12, 2529. [Google Scholar] [CrossRef]
- Fornai, F.; Torracca, M.T.; Bassi, L.; D’Errigo, D.A.; Scalori, V.; Corsini, G.U. Norepinephrine loss selectively enhances chronic nigrostriatal dopamine depletion in mice and rats. Brain Res. 1996, 735, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Iannitelli, A.F.; Kelberman, M.A.; Lustberg, D.J.; Korukonda, A.; McCann, K.E.; Mulvey, B.; Segal, A.; Liles, L.C.; Sloan, S.A.; Dougherty, J.D.; et al. The Neurotoxin DSP-4 Dysregulates the Locus Coeruleus-Norepinephrine System and Recapitulates Molecular and Behavioral Aspects of Prodromal Neurodegenerative Disease. eNeuro 2023, 10, ENEURO.0483-22.2022. [Google Scholar] [CrossRef] [PubMed]
- Chalermpalanupap, T.; Kinkead, B.; Hu, W.T.; Kummer, M.P.; Hammerschmidt, T.; Heneka, M.T.; Weinshenker, D.; Levey, A.I. Targeting norepinephrine in mild cognitive impairment and Alzheimer’s disease. Alzheimer’s Res. Ther. 2013, 5, 21. [Google Scholar] [CrossRef] [PubMed]
- Betts, M.J.; Kirilina, E.; Otaduy, M.C.G.; Ivanov, D.; Acosta-Cabronero, J.; Callaghan, M.F.; Lambert, C.; Cardenas-Blanco, A.; Pine, K.; Passamonti, L.; et al. Locus coeruleus imaging as a biomarker for noradrenergic dysfunction in neurodegenerative diseases. Brain 2019, 142, 2558–2571. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, F.S.; Biagioni, F.; Galgani, A.; Pavese, N.; Lazzeri, G.; Fornai, F. Locus Coeruleus Modulates Neuroinflammation in Parkinsonism and Dementia. Int. J. Mol. Sci. 2020, 21, 8630. [Google Scholar] [CrossRef]
- Mather, M. Noradrenaline in the aging brain: Promoting cognitive reserve or accelerating Alzheimer’s disease? Semin. Cell Dev. Biol. 2021, 116, 108–124. [Google Scholar] [CrossRef]
- Mercan, D.; Heneka, M.T. The Contribution of the Locus Coeruleus-Noradrenaline System Degeneration during the Progression of Alzheimer’s Disease. Biology 2022, 11, 1822. [Google Scholar] [CrossRef]
- Weinshenker, D.; Ferrucci, M.; Busceti, C.L.; Biagioni, F.; Lazzeri, G.; Liles, L.C.; Lenzi, P.; Pasquali, L.; Murri, L.; Paparelli, A.; et al. Genetic or pharmacological blockade of noradrenaline synthesis enhances the neurochemical, behavioral, and neurotoxic effects of methamphetamine. J. Neurochem. 2008, 105, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Ferrucci, M.; Giorgi, F.S.; Bartalucci, A.; Busceti, C.L.; Fornai, F. The effects of locus coeruleus and norepinephrine in methamphetamine toxicity. Curr. Neuropharmacol. 2013, 11, 80–94. [Google Scholar] [CrossRef]
- Lazzeri, G.; Busceti, C.L.; Biagioni, F.; Fabrizi, C.; Morucci, G.; Giorgi, F.S.; Ferrucci, M.; Lenzi, P.; Puglisi-Allegra, S.; Fornai, F. Norepinephrine Protects against Methamphetamine Toxicity through β2-Adrenergic Receptors Promoting LC3 Compartmentalization. Int. J. Mol. Sci. 2021, 22, 7232. [Google Scholar] [CrossRef]
- Choudhary, P.; Pacholko, A.G.; Palaschuk, J.; Bekar, L.K. The locus coeruleus neurotoxin, DSP4, and/or a high sugar diet induce behavioral and biochemical alterations in wild-type mice consistent with Alzheimers related pathology. Metab. Brain Dis. 2018, 33, 1563–1571. [Google Scholar] [CrossRef]
- Kulkarni, A.S.; Del Mar Cortijo, M.; Roberts, E.R.; Suggs, T.L.; Stover, H.B.; Pena-Bravo, J.I.; Steiner, J.A.; Luk, K.C.; Brundin, P.; Wesson, D.W. Perturbation of in vivo Neural Activity Following α-Synuclein Seeding in the Olfactory Bulb. J. Parkinsons Dis. 2020, 10, 1411–1427. [Google Scholar] [CrossRef]
- Yan, Y.; Aierken, A.; Wang, C.; Song, D.; Ni, J.; Wang, Z.; Quan, Z.; Qing, H. A potential biomarker of preclinical Alzheimer’s disease: The olfactory dysfunction and its pathogenesis-based neural circuitry impairments. Neurosci. Biobehav. Rev. 2022, 132, 857–869. [Google Scholar] [CrossRef]
- Tu, L.; Wang, Z.; Lv, X.; Xie, T.; Fan, Z.; Zhang, M.; Wang, H.; Yu, X. Characteristics of Odor Identification and Hedonics and Their Association with Piriform Cortex-Based Resting-State Functional Connectivity in Amnestic Mild Cognitive Impairment. J. Alzheimer’s Dis. 2023, 94, 247–258. [Google Scholar] [CrossRef]
- Mesholam, R.I.; Moberg, P.J.; Mahr, R.N.; Doty, R.L. Olfaction in neurodegenerative disease: A meta-analysis of olfactory functioning in Alzheimer’s and Parkinson’s diseases. Arch. Neurol. 1998, 55, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Ruffoli, R.; Giorgi, F.S.; Pizzanelli, C.; Murri, L.; Paparelli, A.; Fornai, F. The chemical neuroanatomy of vagus nerve stimulation. J. Chem. Neuroanat. 2011, 42, 288–296. [Google Scholar] [CrossRef]
- Giorgi, F.S.; Blandini, F.; Cantafora, E.; Biagioni, F.; Armentero, M.T.; Pasquali, L.; Orzi, F.; Murri, L.; Paparelli, A.; Fornai, F. Activation of brain metabolism and fos during limbic seizures: The role of locus coeruleus. Neurobiol. Dis. 2008, 30, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Radwanska, K.; Nikolaev, E.; Kaczmarek, L. Central noradrenergic lesion induced by DSP-4 impairs the acquisition of avoidance reactions and prevents molecular changes in the amygdala. Neurobiol. Learn. Mem. 2010, 94, 303–311. [Google Scholar] [CrossRef]
- Vadodaria, K.C.; Yanpallewar, S.U.; Vadhvani, M.; Toshniwal, D.; Liles, L.C.; Rommelfanger, K.S.; Weinshenker, D.; Vaidya, V.A. Noradrenergic regulation of plasticity marker expression in the adult rodent piriform cortex. Neurosci. Lett. 2017, 644, 76–82. [Google Scholar] [CrossRef]
- Wilson, R.S.; Arnold, S.E.; Schneider, J.A.; Boyle, P.A.; Buchman, A.S.; Bennett, D.A. Olfactory impairment in presymptomatic Alzheimer’s disease. Ann. N. Y. Acad. Sci. 2009, 1170, 730–735. [Google Scholar] [CrossRef]
- Armstrong, J.N.; Plumier, J.C.; Robertson, H.A.; Currie, R.W. The inducible 70,000 molecular/weight heat shock protein is expressed in the degenerating dentate hilus and piriform cortex after systemic administration of kainic acid in the rat. Neuroscience 1996, 74, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Biagioni, F.; Ferese, R.; Limanaqi, F.; Madonna, M.; Lenzi, P.; Gambardella, S.; Fornai, F. Methamphetamine persistently increases alpha-synuclein and suppresses gene promoter methylation within striatal neurons. Brain Res. 2019, 1719, 157–175. [Google Scholar] [CrossRef] [PubMed]
- Mylvaganam, S.; Earnshaw, R.; Heymann, G.; Kalia, S.K.; Kalia, L.V. C-terminus of Hsp70 Interacting Protein (CHIP) and Neurodegeneration: Lessons from the Bench and Bedside. Curr. Neuropharmacol. 2021, 19, 1038–1068. [Google Scholar] [CrossRef] [PubMed]
- Zatsepina, O.G.; Evgen’ev, M.B.; Garbuz, D.G. Role of a Heat Shock Transcription Factor and the Major Heat Shock Protein Hsp70 in Memory Formation and Neuroprotection. Cells 2021, 10, 1638. [Google Scholar] [CrossRef] [PubMed]
- Akber, U.; Jo, H.; Jeon, S.; Yang, S.J.; Bong, S.; Lim, S.; Kim, Y.K.; Park, Z.Y.; Park, C.S. Cereblon Regulates the Proteotoxicity of Tau by Tuning the Chaperone Activity of DNAJA1. J. Neurosci. 2021, 41, 5138–5156. [Google Scholar] [CrossRef] [PubMed]
- Cecarini, V.; Bonfili, L.; Gogoi, O.; Lawrence, S.; Venanzi, F.M.; Azevedo, V.; Mancha-Agresti, P.; Drumond, M.M.; Rossi, G.; Berardi, S.; et al. Neuroprotective effects of p62(SQSTM1)-engineered lactic acid bacteria in Alzheimer’s disease: A pre-clinical study. Aging 2020, 12, 15995–16020. [Google Scholar] [CrossRef] [PubMed]
- Rocha, S.M.; Kirkley, K.S.; Chatterjee, D.; Aboellail, T.A.; Smeyne, R.J.; Tjalkens, R.B. Microglia-specific knock-out of NF-κB/IKK2 increases the accumulation of misfolded α-synuclein through the inhibition of p62/sequestosome-1-dependent autophagy in the rotenone model of Parkinson’s disease. Glia 2023, 71, 2154–2179. [Google Scholar] [CrossRef]
- Kim, Y.; Park, M.K.; Uhm, D.Y.; Shin, J.; Chung, S. Modulation of delayed rectifier potassium channels by alpha1-adrenergic activation via protein kinase C zeta and p62 in PC12 cells. Neurosci. Lett. 2005, 387, 43–48. [Google Scholar] [CrossRef]
- Fornai, F.; Lenzi, P.; Gesi, M.; Soldani, P.; Ferrucci, M.; Lazzeri, G.; Capobianco, L.; Battaglia, G.; De Blasi, A.; Nicoletti, F.; et al. Methamphetamine produces neuronal inclusions in the nigrostriatal system and in PC12 cells. J. Neurochem. 2004, 88, 114–123. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates, 2nd ed.; Academic Press: San Diego, CA, USA, 2004. [Google Scholar]
- Lowry, O.H.; Rosembrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Dahl, M.J.; Mather, M.; Werkle-Bergner, M.; Kennedy, B.L.; Guzman, S.; Hurth, K.; Miller, C.A.; Qiao, Y.; Shi, Y.; Chui, H.C.; et al. Locus coeruleus integrity is related to tau burden and memory loss in autosomal-dominant Alzheimer’s disease. Neurobiol. Aging 2022, 112, 39–54. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Provider | Catalog Number | RRID | Concentration |
|---|---|---|---|---|
| Monoclonal mouse anti-HSP70 (l.m.; w.b.) | R&D Systems | Cod. MAB-1663 | AB 2119388 | 1:1000 |
| Polyclonal anti-p-Tau (w.b.) | Sigma Aldrich | Cod. P-7444 | * | 1:1000 |
| Horse anti-Mouse IgG antibody (H + L), biotinylated (l.m.) | Vector Labs | BA-2000-1.5 | AB_2313581 | 1:200 |
| Mouse anti-beta-actin (w.b.) | Sigma Aldrich | A1978 | AB_476692 | 1:25,000 |
| Mouse anti-GAPDH (w.b.) | Santa Cruz Biotechnology | SC32233 | AB_627679 | 1:1000 |
| Mouse monoclonal anti-dopamine-beta-hydroxylase (w.b.) | Millipore | MAB308 | AB_2245740 | 1:1000 |
| Rabbit polyclonal anti-alpha-synuclein (l.m.; w.b.) | Sigma Aldrich | SAB4502828 | AB_10746104 | 1:1000 w.b. 1:100 l.m. |
| Horse anti-Rabbit IgG antibody (H + L), Biotinylated (l.m.) | Vector Labs | BA-1100 | AB_2336201 | 1:200 |
| Recombinant rabbit anti-SQSTM1/p62 (l.m.; w.b.) | Abcam | AB109012 | AB_2810880 | 1:1000 w.b. 1:100 l.m. |
| Goat anti-rabbit (w.b.) | Millipore | 401-393 | AB_437797 | 1:3000 |
| Goat anti-mouse (w.b.) | Millipore | 401-215 | AB_10682749 | 1:3000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biagioni, F.; Ferrucci, M.; Lazzeri, G.; Scioli, M.; Frati, A.; Puglisi-Allegra, S.; Fornai, F. Damage to the Locus Coeruleus Alters the Expression of Key Proteins in Limbic Neurodegeneration. Int. J. Mol. Sci. 2024, 25, 3159. https://doi.org/10.3390/ijms25063159
Biagioni F, Ferrucci M, Lazzeri G, Scioli M, Frati A, Puglisi-Allegra S, Fornai F. Damage to the Locus Coeruleus Alters the Expression of Key Proteins in Limbic Neurodegeneration. International Journal of Molecular Sciences. 2024; 25(6):3159. https://doi.org/10.3390/ijms25063159
Chicago/Turabian StyleBiagioni, Francesca, Michela Ferrucci, Gloria Lazzeri, Mariarosaria Scioli, Alessandro Frati, Stefano Puglisi-Allegra, and Francesco Fornai. 2024. "Damage to the Locus Coeruleus Alters the Expression of Key Proteins in Limbic Neurodegeneration" International Journal of Molecular Sciences 25, no. 6: 3159. https://doi.org/10.3390/ijms25063159