
Extracellular Vesicles from Cerebrospinal Fluid of Leptomeningeal Metastasis Patients Deliver MiR-21 and Induce Methotrexate Resistance in Lung Cancer Cells
 ,
,  , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Increased CSF miR-21 Levels in Patients with LM Were Associated with Poor Survival
2.2. Cellular miR-21 Levels Are Inversely Correlated to MTX Sensitivity in NSCLC Cell Lines
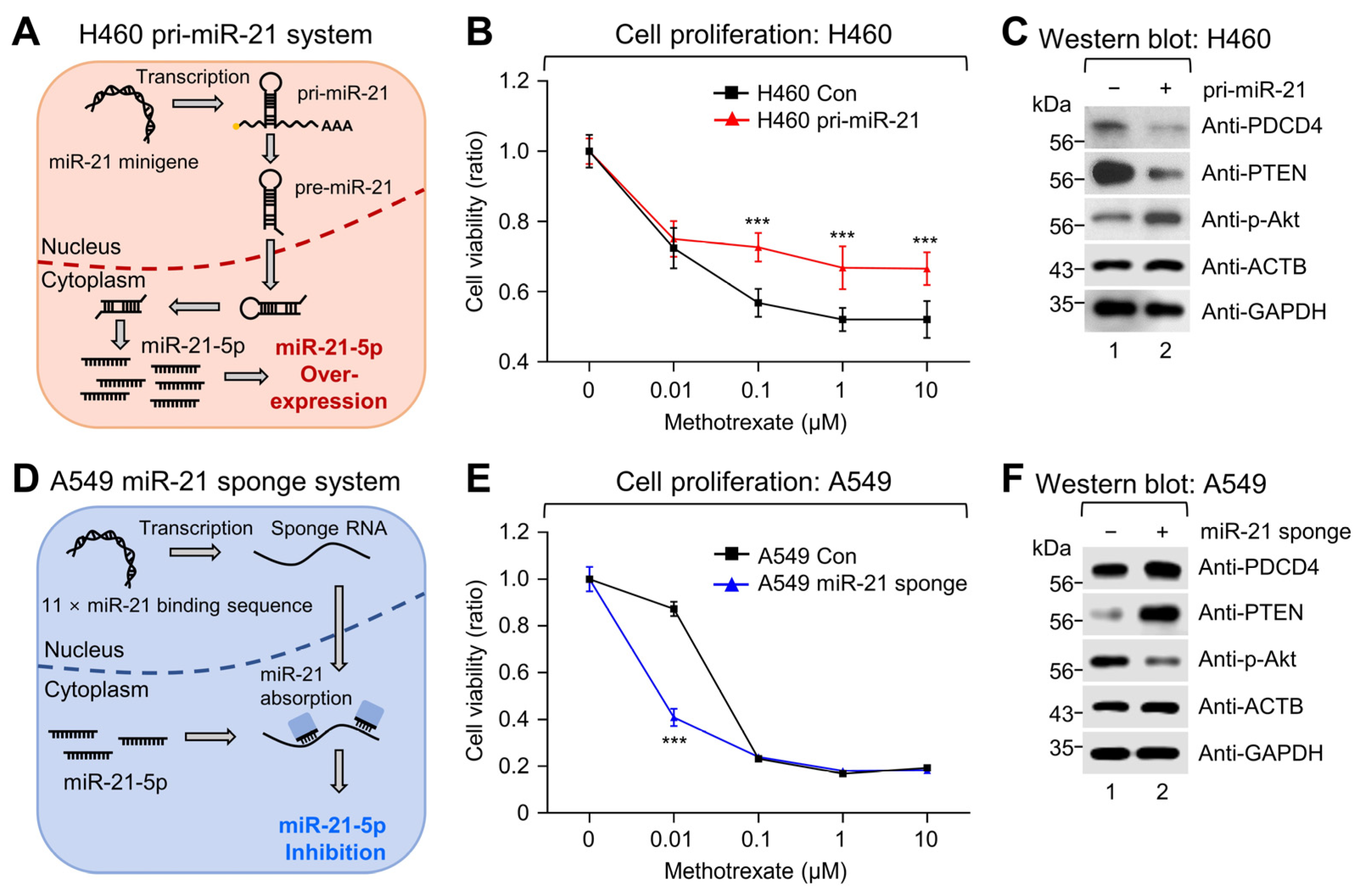
2.3. Modulation of miR-21 Expression Affects MTX Sensitivity and the PTEN-Akt Pathway in NSCLC Cell Lines
2.4. Extracellular Vesicles Transfer miR-21 into Recipient Cancer Cells
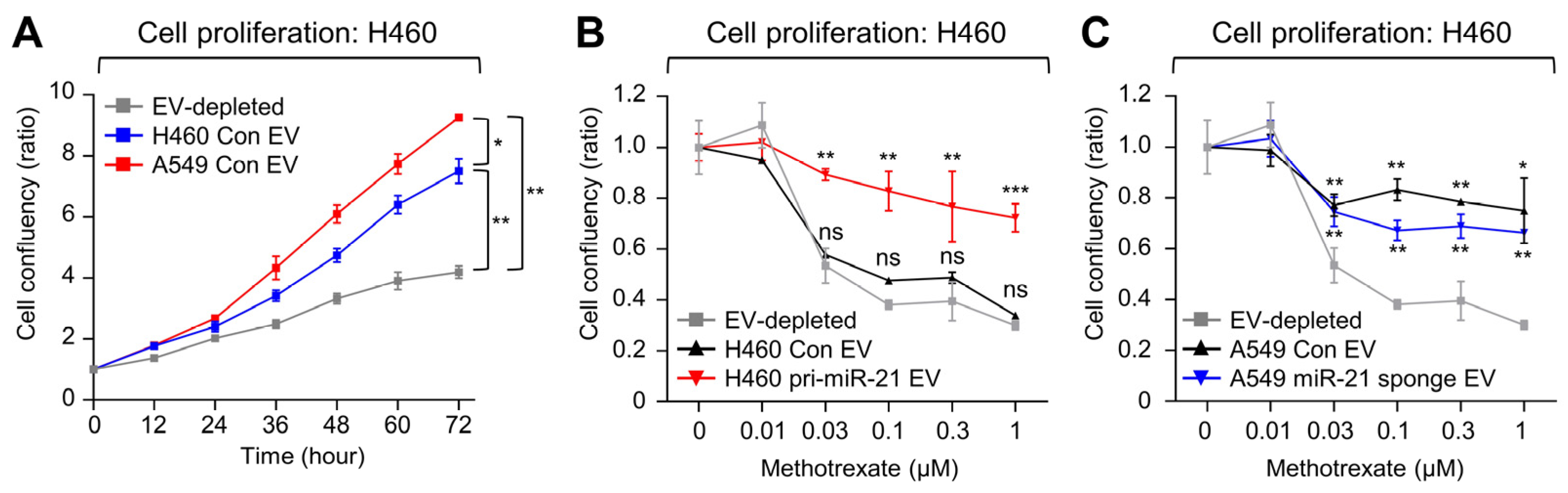
2.5. EVs Containing miR-21 Induce MTX Resistance in Low-miR-21 Recipient Cancer Cells
2.6. EVs from the CSF of Patients with LM Promote MTX Resistance in Recipient Cells
3. Discussion
3.1. miR-21 Mediates MTX Drug Resistance
3.2. EVs Can Modulate the Phenotype of Recipient Cells including Chemo-Resistance
3.3. Clinical Implication of Acquired MTX Resistance in Intra-CSF Chemotherapy
3.4. Limitations
4. Materials and Methods
4.1. Clinical Samples and Patient Survival Analysis
4.2. Cell Proliferation and Determination of the Half-Maximal Inhibitory Concentration (IC50) of MTX
4.3. EV Isolation and Quantification by Nanoparticle Tracking Analysis (NTA)
4.4. Luminescence-Based miR-21-Sensing System
4.5. Western Blot Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taillibert, S.; Laigle-Donadey, F.; Chodkiewicz, C.; Sanson, M.; Hoang-Xuan, K.; Delattre, J.Y. Leptomeningeal metastases from solid malignancy: A review. J. Neurooncol. 2005, 75, 85–99. [Google Scholar] [CrossRef]
- Chamberlain, M.C. Leptomeningeal metastasis. Curr. Opin. Oncol. 2010, 22, 627–635. [Google Scholar] [CrossRef]
- Chowdhary, S.; Chamberlain, M. Leptomeningeal metastases: Current concepts and management guidelines. J. Natl. Compr. Canc. Netw. 2005, 3, 693–703. [Google Scholar] [CrossRef]
- Gwak, H.S.; Lee, S.H.; Park, W.S.; Shin, S.H.; Yoo, H.; Lee, S.H. Recent Advancements of Treatment for Leptomeningeal Carcinomatosis. J. Korean Neurosurg. Soc. 2015, 58, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bokstein, F.; Lossos, A.; Siegal, T. Leptomeningeal metastases from solid tumors: A comparison of two prospective series treated with and without intra-cerebrospinal fluid chemotherapy. Cancer 1998, 82, 1756–1763. [Google Scholar] [CrossRef]
- Lee, K.Y.; Im, J.H.; Lin, W.; Gwak, H.S.; Kim, J.H.; Yoo, B.C.; Kim, T.H.; Park, J.B.; Park, H.J.; Kim, H.J.; et al. Nanoparticles in 472 Human Cerebrospinal Fluid: Changes in Extracellular Vesicle Concentration and miR-21 Expression as a Biomarker for Leptomeningeal Metastasis. Cancers 2020, 12, 2745. [Google Scholar] [CrossRef] [PubMed]
- Gebert, L.F.R.; MacRae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell. Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Rhim, J.; Baek, W.; Seo, Y.; Kim, J.H. From Molecular Mechanisms to Therapeutics: Understanding MicroRNA-21 in Cancer. Cells 2022, 11, 2791. [Google Scholar] [CrossRef]
- Teplyuk, N.M.; Mollenhauer, B.; Gabriely, G.; Giese, A.; Kim, E.; Smolsky, M.; Kim, R.Y.; Saria, M.G.; Pastorino, S.; Kesari, S.; et al. MicroRNAs in cerebrospinal fluid identify glioblastoma and metastatic brain cancers and reflect disease activity. Neuro-oncology 2012, 14, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Lu, X.; Liu, L.; Xu, J.; Feng, D.; Shu, Y. MiRNA-21: A biomarker predictive for platinum-based adjuvant chemotherapy response in patients with non-small cell lung cancer. Cancer Biol. Ther. 2012, 13, 330–340. [Google Scholar] [CrossRef]
- Yin, J.; Park, G.; Lee, J.E.; Choi, E.Y.; Park, J.Y.; Kim, T.H.; Park, N.; Jin, X.; Jung, J.E.; Shin, D.; et al. DEAD-box RNA helicase DDX23 modulates glioma malignancy via elevating miR-21 biogenesis. Brain 2015, 138, 2553–2570. [Google Scholar] [CrossRef]
- Kwak, H.J.; Kim, Y.J.; Chun, K.R.; Woo, Y.M.; Park, S.J.; Jeong, J.A.; Jo, S.H.; Kim, T.H.; Min, H.S.; Chae, J.S.; et al. Downregulation of Spry2 by miR-21 triggers malignancy in human gliomas. Oncogene 2011, 30, 2433–2442. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.X.; Lu, B.B.; Wang, H.; Cheng, Z.X.; Yin, Y.M. MicroRNA-21 modulates chemosensitivity of breast cancer cells to doxorubicin by targeting PTEN. Arch. Med. Res. 2011, 42, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.G.; Wang, J.J.; Zhao, F.; Liu, Q.; Jiang, K.; Yang, G.H. MicroRNA-21 (miR-21) represses tumor suppressor PTEN and promotes growth and invasion in non-small cell lung cancer (NSCLC). Clin. Chim. Acta 2010, 411, 846–852. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Seo, Y.; Im, J.H.; Rhim, J.; Baek, W.; Kim, S.; Kwon, J.W.; Yoo, B.C.; Shin, S.H.; Yoo, H.; et al. Molecular Signature of Extracellular Vesicular Small Non-Coding RNAs Derived from Cerebrospinal Fluid of Leptomeningeal Metastasis Patients: Functional Implication of miR-21 and Other Small RNAs in Cancer Malignancy. Cancers 2021, 13, 209. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Odenthal, M.; Fries, J.W. Exosomes as miRNA Carriers: Formation-Function-Future. Int. J. Mol. Sci. 2016, 17, 2028. [Google Scholar] [CrossRef] [PubMed]
- Gwak, H.S.; Joo, J.; Shin, S.H.; Yoo, H.; Han, J.Y.; Kim, H.T.; Yun, T.; Ro, J.; Lee, J.S.; Lee, S.H. Ventriculolumbar perfusion chemotherapy with methotrexate for treating leptomeningeal carcinomatosis: A Phase II Study. Oncologist 2014, 19, 1044–1045. [Google Scholar] [CrossRef] [PubMed]
- Bica-Pop, C.; Cojocneanu-Petric, R.; Magdo, L.; Raduly, L.; Gulei, D.; Berindan-Neagoe, I. Overview upon miR-21 in lung cancer: Focus on NSCLC. Cell. Mol. Life Sci. 2018, 75, 3539–3551. [Google Scholar] [CrossRef]
- Vella, L.J.; Behren, A.; Coleman, B.; Greening, D.W.; Hill, A.F.; Cebon, J. Intercellular Resistance to BRAF Inhibition Can Be Mediated by Extracellular Vesicle-Associated PDGFRbeta. Neoplasia 2017, 19, 932–940. [Google Scholar] [CrossRef]
- Lucero, R.; Zappulli, V.; Sammarco, A.; Murillo, O.D.; Cheah, P.S.; Srinivasan, S.; Tai, E.; Ting, D.T.; Wei, Z.; Roth, M.E.; et al. Glioma-Derived miRNA-Containing Extracellular Vesicles Induce Angiogenesis by Reprogramming Brain Endothelial Cells. Cell Rep. 2020, 30, 2065–2074. [Google Scholar] [CrossRef]
- Peinado, H.; Aleckovic, M.; Lavotshkin, S.; Matei, I.; Costa-Silva, B.; Moreno-Bueno, G.; Hergueta-Redondo, M.; Williams, C.; Garcia-Santos, G.; Ghajar, C.; et al. Melanoma exosomes educate bone marrow progenitor cells toward a pro-metastatic phenotype through MET. Nat. Med. 2012, 18, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.; Kang, Y.; Ham, Y.; Kim, M.H.; Kim, S.J.; Yoon, S.K.; Jang, S.K.; Park, J.B.; Cho, S.; Kim, J.H. PLK1-ELAVL1/HuR-miR-122 signaling facilitates hepatitis C virus proliferation. Proc. Natl. Acad. Sci. USA 2022, 119, e2214911119. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Seo, Y.; Rhim, J.; Jin, X.; Kim, T.H.; Kim, S.S.; Hong, J.-H.; Gwak, H.-S.; Yoo, H.; Park, J.B.; et al. Crosstalk between PARN and EGFR-STAT3 Signaling Facilitates Self-Renewal and Proliferation of Glioblastoma Stem Cells. Cancer Res. 2023, 83, 3693–3709. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.; Kim, S.S.; Kim, N.; Cho, S.; Park, J.B.; Kim, J.H. Development of a miRNA-controlled dual-sensing system and its application for targeting miR-21 signaling in tumorigenesis. Exp. Mol. Med. 2020, 52, 1989–2004. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Zhao, J.; Tao, Y.; Guo, M.; Ya, Z.; Chen, C.; Qin, N.; Zheng, J.; Luo, J.; Xu, L. MicroRNA-21: A promising biomarker for the prognosis and diagnosis of non-small cell lung cancer. Oncol. Lett. 2018, 16, 2777–2782. [Google Scholar] [CrossRef] [PubMed]
- Hummel, R.; Wang, T.; Watson, D.I.; Michael, M.Z.; Van der Hoek, M.; Haier, J.; Hussey, D.J. Chemotherapy-induced modification of microRNA expression in esophageal cancer. Oncol. Rep. 2011, 26, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, X.; Huang, Z.; Wang, J.; Zhu, W.; Shu, Y.; Liu, P. Prognostic value of miR-21 in various cancers: An updating meta-analysis. PLoS ONE 2014, 9, e102413. [Google Scholar] [CrossRef] [PubMed]
- Buscaglia, L.E.; Li, Y. Apoptosis and the target genes of microRNA-21. Chin. J. Cancer 2011, 30, 371–380. [Google Scholar] [CrossRef]
- Feng, Y.H.; Tsao, C.J. Emerging role of microRNA-21 in cancer. Biomed. Rep. 2016, 5, 395–402. [Google Scholar] [CrossRef]
- Xu, L.; Huang, Y.; Chen, D.; He, J.; Zhu, W.; Zhang, Y.; Liu, X. Downregulation of miR-21 increases cisplatin sensitivity of non-small-cell lung cancer. Cancer Genet. 2014, 207, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Zhu, Y.; Ali, D.J.; Tian, T.; Xu, H.; Si, K.; Sun, B.; Chen, B.; Xiao, Z. Engineered exosomes for targeted co-delivery of miR-21 inhibitor and chemotherapeutics to reverse drug resistance in colon cancer. J. Nanobiotechnology 2020, 18, 10. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Thery, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell. Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef]
- Zhao, S.; Mi, Y.; Zheng, B.; Wei, P.; Gu, Y.; Zhang, Z.; Xu, Y.; Cai, S.; Li, X.; Li, D. Highly-metastatic colorectal cancer cell released miR-181a-5p-rich extracellular vesicles promote liver metastasis by activating hepatic stellate cells and remodelling the tumour microenvironment. J. Extracell. Vesicles 2022, 11, e12186. [Google Scholar] [CrossRef]
- Zhuang, H.; Wang, H.; Yang, H.; Li, H. Exosome-Encapsulated MicroRNA-21 from Esophageal Squamous Cell Carcinoma Cells Enhances Angiogenesis of Human Umbilical Venous Endothelial Cells by Targeting SPRY1. Cancer Manag. Res. 2020, 12, 10651–10667. [Google Scholar] [CrossRef]
- Ren, W.; Hou, J.; Yang, C.; Wang, H.; Wu, S.; Wu, Y.; Zhao, X.; Lu, C. Extracellular vesicles secreted by hypoxia pre-challenged mesenchymal stem cells promote non-small cell lung cancer cell growth and mobility as well as macrophage M2 polarization via miR-21-5p delivery. J. Exp. Clin. Cancer Res. 2019, 38, 62. [Google Scholar] [CrossRef] [PubMed]
- Cornell, L.; Wander, S.A.; Visal, T.; Wagle, N.; Shapiro, G.I. MicroRNA-Mediated Suppression of the TGF-beta Pathway Confers Transmissible and Reversible CDK4/6 Inhibitor Resistance. Cell Rep. 2019, 26, 2667–2680. [Google Scholar] [CrossRef]
- Chamberlain, M.C.; Kormanik, P. Carcinoma meningitis secondary to non-small cell lung cancer: Combined modality therapy. Arch. Neurol. 1998, 55, 506–512. [Google Scholar] [CrossRef]
- Kim, K.H.; Park, M.; Park, E.Y.; Gwak, H.S.; Kim, S.H.; Seo, J.W.; Hyun, J.W.; Kim, H.J.; Dho, Y.S.; Shin, S.H.; et al. Disseminating Necrotizing Leukoencephalopathy Associated with Intra-CSF Methotrexate Chemotherapy: A Retrospective Observational Study. Neurology 2024, 102, e209167. [Google Scholar] [CrossRef]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Im, J.H.; Lee, K.-Y.; Seo, Y.; Rhim, J.; Dho, Y.-S.; Yoo, B.C.; Park, J.B.; Shin, S.H.; Yoo, H.; Kim, J.H.; et al. Extracellular Vesicles from Cerebrospinal Fluid of Leptomeningeal Metastasis Patients Deliver MiR-21 and Induce Methotrexate Resistance in Lung Cancer Cells. Int. J. Mol. Sci. 2024, 25, 3124. https://doi.org/10.3390/ijms25063124
Im JH, Lee K-Y, Seo Y, Rhim J, Dho Y-S, Yoo BC, Park JB, Shin SH, Yoo H, Kim JH, et al. Extracellular Vesicles from Cerebrospinal Fluid of Leptomeningeal Metastasis Patients Deliver MiR-21 and Induce Methotrexate Resistance in Lung Cancer Cells. International Journal of Molecular Sciences. 2024; 25(6):3124. https://doi.org/10.3390/ijms25063124
Chicago/Turabian StyleIm, Ji Hye, Kyue-Yim Lee, Yoona Seo, Jiho Rhim, Yun-Sik Dho, Byong Chul Yoo, Jong Bae Park, Sang Hoon Shin, Heon Yoo, Jong Heon Kim, and et al. 2024. "Extracellular Vesicles from Cerebrospinal Fluid of Leptomeningeal Metastasis Patients Deliver MiR-21 and Induce Methotrexate Resistance in Lung Cancer Cells" International Journal of Molecular Sciences 25, no. 6: 3124. https://doi.org/10.3390/ijms25063124