
Morphological Analyses and QTL Mapping of Mottled Leaf in Zucchini (Cucurbita pepo L.)
Abstract
:1. Introduction
2. Results
2.1. Observation of the Phenotypes Associated with the Mottled Leaf Trait
2.2. Determination of the Content of Photosynthetic Pigment in Leaves
2.3. Analysis of Photosynthetic Parameters in Leaves
2.4. Shading Analysis of Leaf
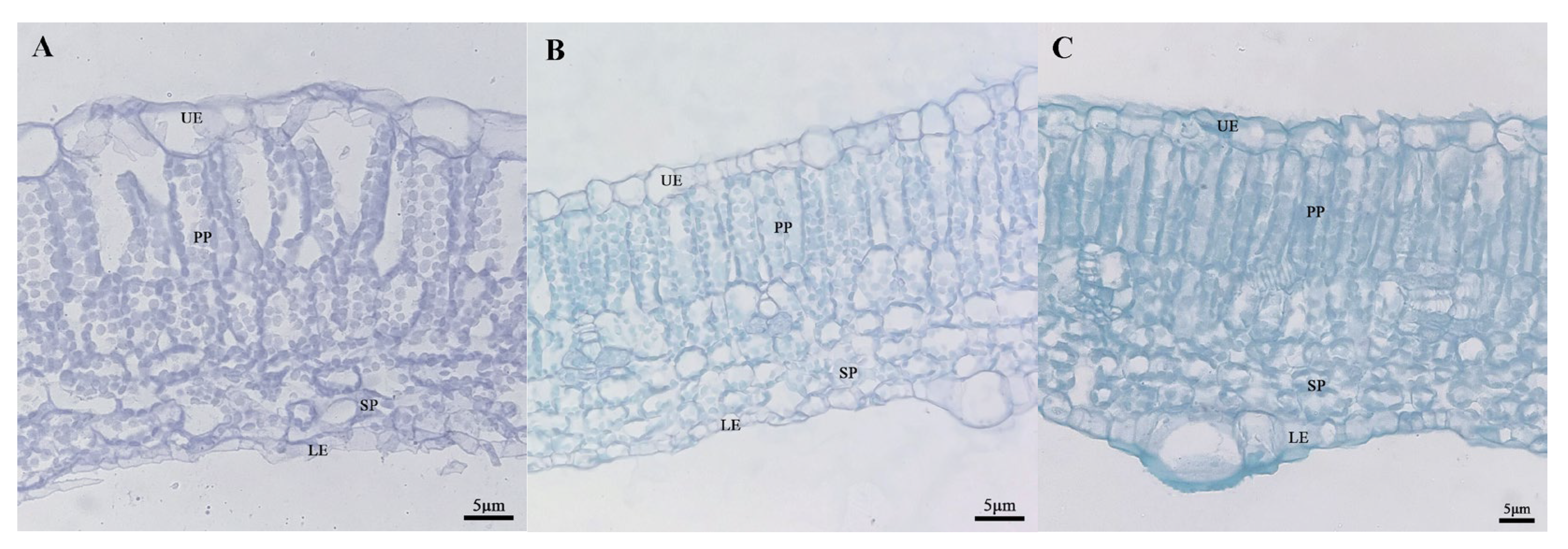
2.5. Analysis of Differences in the Microstructure of Leaves
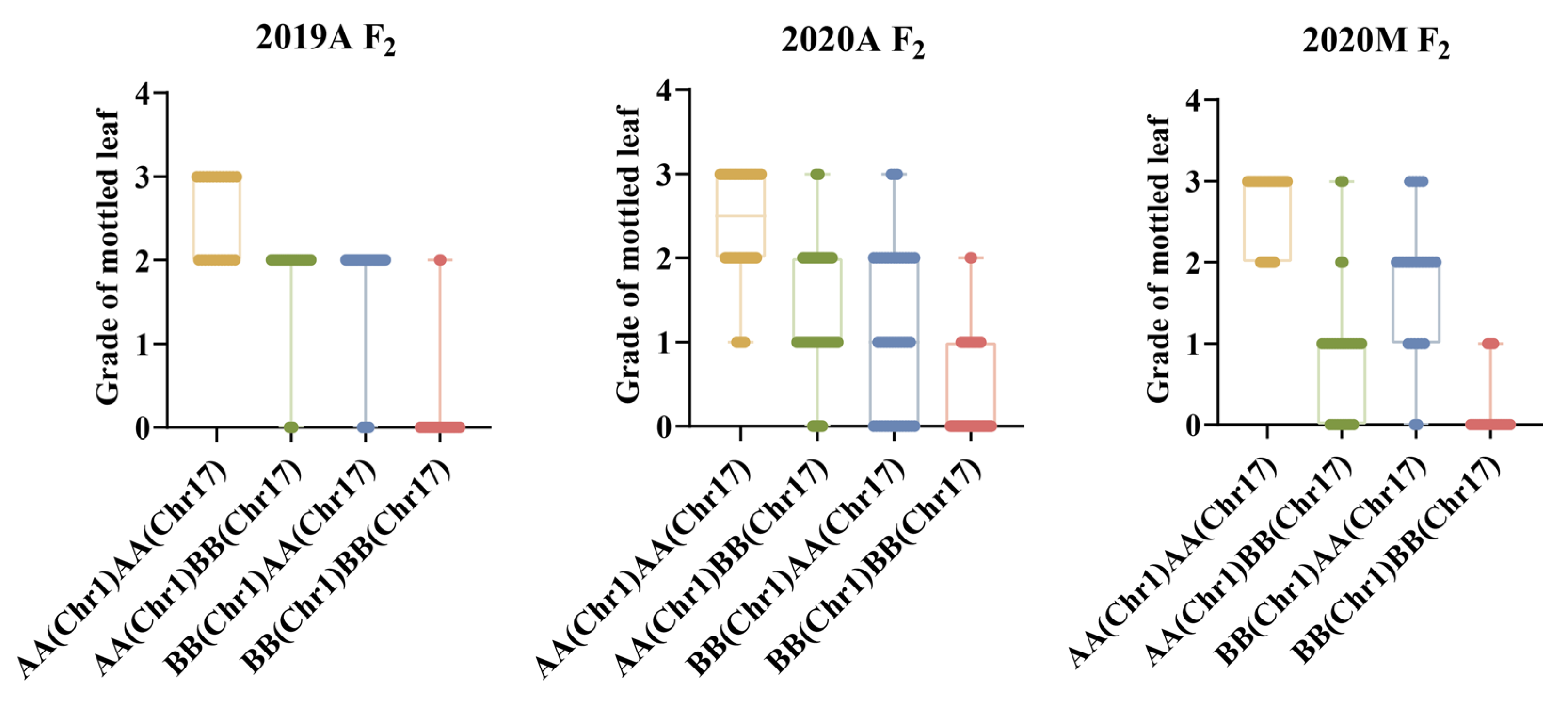
2.6. Genetic Analysis of Mottled Leaf Trait
2.7. QTL Analysis of the Mottled Leaf Trait
2.8. QTL Interaction Analysis of Mottled Leaf Trait
2.9. Candidate Gene Analysis of Major QTLs
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Determination of Photosynthetic Pigments
4.3. Determination of Photosynthetic Parameters
4.4. Leaf Anatomy Assay
4.5. Inheritance Analysis of the Mottled Leaf Trait
4.6. Pool Construction and QTL-seq Analysis
4.7. QTL Analysis with Molecular Markers
4.8. Data Analysis
4.9. Prediction of Candidate Genes and qRT-PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vairam, N.; Ibrahim, S.-M.; Vanniarajan, C. Frequency and spectrum of chlorophyll mutations in greengram [Vigna radiata (L.) Wilczek]. Asian J. Biol. Sci. 2014, 9, 204–207. [Google Scholar] [CrossRef]
- Zhang, J.-H.; Zeng, J.-C.; Wang, X.-M.; Chen, S.-F.; Albach, D.C.; Li, H.-Q. A revised classification of leaf variegation types. Flora 2020, 272, 151703. [Google Scholar] [CrossRef]
- Aluru, M.-R.; Bae, H.; Wu, D.; Rodermel, S.-R. The Arabidopsis immutans mutation affects plastid differentiation and the morphogenesis of white and green sectors in variegated plants. Plant Physiol. 2001, 127, 67–77. [Google Scholar] [CrossRef]
- Wetzel, C.-M.; Jiang, C.-Z.; Meehan, L.-J.; Voytas, D.F.; Rodermel, S.-R. Nuclear-organelle interactions: The immutans variegation mutant of Arabidopsis is plastid autonomous and impaired in carotenoid biosynthesis. Plant J. 1994, 6, 161–175. [Google Scholar] [CrossRef]
- Carol, P.; Stevenson, D.; Bisanz, C.; Breitenbach, J.; Sandmann, G.; Mache, R.; Coupland, G.; Kuntz, M. Mutations in the Arabidopsis gene IMMUTANS cause a variegated phenotype by inactivating a chloroplast terminal oxidase associated with phytoene desaturation. Plant Cell 1999, 11, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Næsted, H.; Holm, A.; Jenkins, T.; Nielsen, H.-B.; Harris, C.-A.; Beale, M.-H.; Andersen, M.; Mant, A.; Scheller, H.; Camara, B.; et al. Arabidopsis VARIEGATED 3 encodes a chloroplast-targeted, zinc-finger protein required for chloroplast and palisade cell development. J. Cell Sci. 2004, 117, 4807–4818. [Google Scholar] [CrossRef]
- Haswell, E.-S.; Meyerowitz, E.-M. MscS-like proteins control plastid size and shape in Arabidopsis thaliana. Curr. Biol. 2006, 16, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, W.; Tamura, T.; Hanba-Tomita, Y.; Murata, M.; Sodmergen. The VAR1 locus of Arabidopsis encodes a chloroplastic FtsH and is responsible for leaf variegation in the mutant alleles. Genes Cells 2002, 7, 769–780. [Google Scholar] [CrossRef]
- Chen, M.; Choi, Y.; Voytas, D.-F.; Rodermel, S. Mutations in the Arabidopsis VAR2 locus cause leaf variegation due to the loss of a chloroplast FtsH protease. Plant J. 2000, 22, 303–313. [Google Scholar] [CrossRef]
- Bailey, S.; Thompson, E.; Nixon, P.-J.; Horton, P.; Mullineaux, C.-W.; Robinson, C.; Mann, N. A critical role for the Var2 FtsH homologue of Arabidopsis thaliana in the photosystem II repair cycle in vivo. J. Biol. Chem. 2002, 277, 2006–2011. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yu, F.; Rodermel, S. Arabidopsis chloroplast FtsH, var2 and suppressors of var2 leaf variegation: A review. J. Integr. Plant Biol. 2010, 52, 750–761. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-W.; Wang, Z.-Y.; Gu, H.; You, J.; Hu, M.-M.; Zhang, Y.-J.; Zhu, Z.; Wang, Y.-H.; Liu, S.-J.; Chen, L.-M.; et al. Identification and Phenotypic Characterization of ZEBRA LEAF16 Encoding a β-Hydroxyacyl-ACP Dehydratase in Rice. Front. Plant Sci. 2018, 9, 782. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-Q.; Zhong, P.; Gao, Z.-Y.; Zhu, B.-Y.; Chen, D.; Su, C.-H.; Wang, P.-R.; Deng, X.-J. Morphological characterization and candidate gene analysis of zebra leaf mutant zebra524 in rice. Sci. Agric. Sin. 2014, 47, 2907–2915. [Google Scholar]
- Qiu, Z.-N.; Chen, D.-D.; He, L.; Zhang, S.; Yang, Z.-N.; Wang, Z.-W.; Ren, D.-Y.; Qian, Q.; Guo, L.-B.; Zhu, L. The rice white green leaf 2 gene causes defects in chloroplast development and affects the plastid ribosomal protein S9. Rice 2018, 11, 39. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Feng, P.; Li, Y.-F.; Yu, G.-L.; Sang, X.-C.; Ling, Y.-H.; Zeng, X.-Q.; Li, Y.-D.; Huang, J.-Y. VIRESCENT-ALBINO LEAF 1 regulates leaf colour development and cell division in rice. J. Exp. Bot. 2018, 69, 4791–4804. [Google Scholar] [CrossRef]
- Feng, P.; Shi, J.-Q.; Zhang, T.; Zhong, Y.-Q.; Zhang, L.-S.; Yu, G.-L.; Zhang, T.-Q.; Zhu, X.-Y.; Xing, Y.-D.; Yin, W.-Z.; et al. Zebra leaf 15, a receptor-like protein kinase involved in moderate low temperature signaling pathway in rice. Rice 2019, 12, 1–13. [Google Scholar] [CrossRef]
- Li, Q.-G.; Du, J.-Y.; Qiao, Z.-H.; Pan, C.; He, W.-Q.; Zhang, L.; Li, X.-H.; Nie, Y.-X.; Li, X.-Z.; Pan, G.-T.; et al. White and green striate leaves 1, predicted to encode a 16S rRNA processing protein, plays a critical role in the processing of chloroplast ribosomes in maize (Zea mays L.). Mol. Breed. 2023, 43, 65. [Google Scholar] [CrossRef]
- Hu, L.-L.; Zhang, H.-Q.; Xie, C.; Wang, J.; Zhang, J.-Y.; Wang, H.; Weng, Y.-Q.; Chen, P.; Li, Y.-H. A mutation in CsHD encoding a histidine and aspartic acid domain-containing protein leads to yellow young leaf-1 (yyl-1) in cucumber (Cucumis sativus L.). Plant Sci. 2020, 293, 110407. [Google Scholar] [CrossRef]
- Ding, Y.; Yang, W.; Su, C.-G.; Ma, H.-H.; Pan, Y.; Zhang, X.-G.; Li, J.-H. Tandem 13-lipoxygenase genes in a cluster confers yellow-green leaf in cucumber. Int. J. Mol. Sci. 2019, 20, 3102. [Google Scholar] [CrossRef]
- Scarchuk, J. Fruit and leaf characters in summer squash: The interrelationship of striped-fruit and mottled-leaf. J. Hered. 1954, 45, 295–297. [Google Scholar] [CrossRef]
- Scott, D.-H.; Riner, M.-E. A Mottled-Leaf Character in Winter Squash: Inherited as a Dominant Mendelian Character. J. Hered. 1946, 37, 27–28. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Anido, F.; Cointry, E.; Firpo, I.; García, S.-M.; Gattuso, S. Inheritance of gray leaf color in a material derived from a Cucurbita maxima Duch. x C. moschata Duch. hybrid. Rep. Cucurbit Genet. Coop. 2002, 25, 46–48. [Google Scholar]
- Ribeiro, A.; da Costa, C.-P. Inheritance of mottled leaf in Cucurbita moschata. Cucurbit Genet. Coop. 1989, 29, 70–71. [Google Scholar]
- Shifriss, O. Further notes on the silvery-leaf trait in Cucurbita. Cucurbit Genet Coop. Rep. 1984, 7, 81–83. [Google Scholar]
- Shifriss, O. Do Cucurbita plants with silvery leaves escape virus infection? Origin and characteristics of NJ260. Cucurbit Genet. Coop. 1981, 4, 42–43. [Google Scholar]
- Shifriss, O. Reflected light spectra from silvery and nonsilvery leaves of Cucurbita pepo. Cucurbit Genet. Coop. 1983, 44, 89–90. [Google Scholar]
- Mcauslane, H.-J.; Chen, J.; Carle, R.-B.; Schmalstig, J. Influence of Bemisia argentifolii (Homoptera: Aleyrodidae) infestation and squash silverleaf disorder on zucchini seedling growth. J. Econ. Entomol. 2004, 97, 1096–1105. [Google Scholar] [CrossRef]
- Jiménez, D.-R.; Yokomi, R.-K.; Mayer, R.-T.; Shapiro, J.-P. Cytology and physiology of silver leaf whitefly-induced squash silver leaf. Physiol. Mol. Plant Pathol. 1995, 46, 227–242. [Google Scholar] [CrossRef]
- Brown, R.-N.; Myers, J.-R. A genetic map of squash (Cucurbita sp.) with randomly amplified polymorphic DNA markers and morphological markers. J. Am. Soc. Hortic. Sci. 2002, 127, 568–575. [Google Scholar] [CrossRef]
- Montero-Pau, J.; Blanca, J.; Esteras, C.; Martínez-Pérez, E.-M.; Gómez, P.; Monforte, A.-J.; Cañizares, J.; Picó, B. An SNP-based saturated genetic map and QTL analysis of fruit-related traits in Zucchini using Genotyping-by-sequencing. BMC Genom. 2017, 18, 94. [Google Scholar] [CrossRef]
- Li, K.-T.; Zhang, J.; Kang, Y.-H.; Chen, M.-C.; Song, T.-T.; Geng, H.; Tian, J. McMYB10 modulates the expression of a Ubiquitin Ligase, McCOP1 during leaf coloration in crabapple. Front. Plant Sci. 2018, 9, 704. [Google Scholar] [CrossRef] [PubMed]
- Scarchuk, J.; Lent, J.-M. The structure of mottled-leaf summer squash. J. Hered. 1965, 56, 167–168. [Google Scholar] [CrossRef]
- Zhang, Y.; Hayashi, T.; Hosokawa, M.; Yazawa, S.; Li, Y. Metallic lustre and the optical mechanism generated from the leaf surface of Begonia rex Putz. Sci. Hortic. 2009, 121, 213–217. [Google Scholar] [CrossRef]
- La Rocca, N.; Pupillo, P.; Puppi, G.; Rascio, N. Erythronium dens-canis L.(Liliaceae): An unusual case of change of leaf mottling. Plant Physiol. Biochem. 2014, 74, 108–117. [Google Scholar] [CrossRef]
- Cao, W.; Du, Y.; Wang, C.; Xu, L.; Wu, T. Cscs encoding chorismate synthase is a candidate gene for leaf variegation mutation in cucumber. Breed. Sci. 2018, 68, 571–581. [Google Scholar] [CrossRef]
- Hamada, T. Microtubule organization and microtubule-associated proteins in plant cells. Int. Rev. Cell Mol. Biol. 2014, 312, 1. [Google Scholar]
- Hashimoto, T. Microtubules in plants. Arab. Book Am. Soc. Plant Biol. 2015, 13, e0179. [Google Scholar] [CrossRef] [PubMed]
- Smertenko, A.; Clare, S.-J.; Effertz, K.; Parish, A.; Ross, A.; Schmidt, S. A guide to plant TPX2-like and WAVE-DAMPENED2-like proteins. J. Exp. Bot. 2021, 72, 1034–1045. [Google Scholar] [CrossRef]
- Dvořák Tomaštíková, E.; Rutten, T.; Dvořák, P.; Tugai, A.; Ptosková, K.; Petrovská, B.; van Damme, D.; Houben, A.; Dolezel, J.; Demidov, D. Functional divergence of microtubule-associated TPX2 family members in Arabidopsis thaliana. Int. J. Mol. Sci. 2020, 21, 2183. [Google Scholar] [CrossRef]
- Sun, J.; Ma, Q.; Mao, T. Ethylene regulates the Arabidopsis microtubule-associated protein WAVE-DAMPENED2-LIKE5 in etiolated hypocotyl elongation. Plant Physiol. 2015, 169, 325–337. [Google Scholar] [CrossRef]
- Du, P.-Z.; Kumar, M.; Yao, Y.; Xie, Q.-L.; Wang, J.-Y.; Zhang, B.-L.; Gan, S.-M.; Wang, Y.-Q.; Wu, A.-M. Genome-wide analysis of the TPX2 family proteins in Eucalyptus grandis. BMC Genom. 2016, 17, 967. [Google Scholar] [CrossRef]
- Ambrose, J.-C.; Shoji, T.; Kotzer, A.-M.; Pighin, J.-A.; Wasteneys, G.-O. The Arabidopsis CLASP Gene Encodes a Microtubule-Associated Protein Involved in Cell Expansion and Division. Plant Cell 2007, 19, 2763–2775. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ambrose, C. CLASP balances two competing cell division plane cues during leaf development. Nat. Plants 2022, 8, 682–693. [Google Scholar] [CrossRef]
- Montero-Pau, J.; Blanca, J.; Bombarely, A.; Ziarsolo, P.; Esteras, C.; Marti-Gómez, C.; Ferriol, M.; Gómez, P.; Jamilena, M.; Mueller, L.; et al. De novo assembly of zucchini genome reveals a whole-genome duplication associated with the origin of the Cucurbita genus. Plant Biotechnol. J. 2018, 16, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.-P.; Dan, Y.; Yu, H.-Y.; Chen, F.-Y.; Wang, K.-X.; Ding, W.-Q.; Xu, W.-L.; Wang, Y.-L. QTL analysis of early flowering of female flowers in zucchini (Cucurbita pepo L.). J. Integr. Agric. 2023, 22, 3321–3330. [Google Scholar] [CrossRef]
- Wang, Y.-L.; Wang, G.-C.; Lin, D.-J.; Luo, Q.-F.; Xu, W.-L.; Qu, S.-P. QTL mapping and stability analysis of trichome density in zucchini (Cucurbita pepo L.). Front. Plant Sci. 2023, 14, 1232154. [Google Scholar] [CrossRef]
- Wang, G.-C.; Dai, D.-Y.; Wang, L.; Sheng, Y.-Y.; Wang, D.; Li, D.-D.; Tian, L.-M.; Luan, F.-S. QTL analysis of flowering-related traits by specific length amplified fragment sequencing in melon. Crop Sci. 2022, 62, 203–215. [Google Scholar] [CrossRef]
- Weng, Y.-Q.; Colle, M.; Wang, Y.-H.; Yang, L.-M.; Rubinstein, M.; Sherman, A.; Ophir, R.; Grumet, R. QTL mapping in multiple populations and development stages reveals dynamic QTL for fruit size in cucumbers of different market classes. Theor. Appl. Genet. 2015, 128, 1747–1763. [Google Scholar] [CrossRef]
- Ding, W.-Q.; Wang, Y.-L.; Qi, C.; Luo, Y.-S.; Wang, C.-J.; Xu, W.-L.; Qu, S.-P. Fine mapping identified the gibberellin 2-oxidase gene CpDw leading to a dwarf phenotype in squash (Cucurbita pepo L.). Plant Sci. 2021, 306, 110857. [Google Scholar] [CrossRef]
- Luo, Y.-S.; Wang, C.-J.; Wang, M.-M.; Wang, Y.-L.; Xu, W.-L.; Han, H.-Y.; Wang, Z.-C.; Zhong, Y.-J.; Huang, H.-X.; Qu, S.-P. Accumulation of carotenoids and expression of carotenoid biosynthesis genes in fruit flesh during fruit development in two Cucurbita maxima inbred lines. Hortic. Plant J. 2021, 7, 529–538. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material | Leaf Thickness | Thickness of Upper Epidermis | Thickness of Palisade Parenchyma | Thickness of Sponge Parenchyma | Thickness of Lower Epidermis | CTR (%) | SR (%) |
|---|---|---|---|---|---|---|---|
| ‘19’ mottle area | 45.12 ± 0.09 a | 4.64 ± 0.16 a | 26.24 ± 0.10 a | 8.69 ± 0.05 c | 1.99 ± 0.10 a | 58.15 ± 0.00 a | 19.26 ± 0.00 c |
| ‘19’ green area | 45.09 ± 0.16 a | 3.82 ± 0.08 b | 26.22 ± 0.35 a | 11.20 ± 0.06 a | 2.32 ± 0.04 a | 58.18 ± 0.01 a | 24.83 ± 0.00 a |
| ‘113’ | 43.66 ± 0.14 b | 3.80 ± 0.04 bB | 22.26 ± 0.20 b | 10.24 ± 0.24 b | 2.32 ± 0.41 a | 50.98 ± 0.01 b | 23.45 ± 0.01 b |
| QTL | Environment | Position Interval | Physical Distance (Kb) | LOD | Add | Dom | PVE (%) |
|---|---|---|---|---|---|---|---|
| CpML1.1 | 2019A | CHr01_18713051-CHr01_19638223 | 925.2 | 43.79 | 0.64 | 0.36 | 24.15 |
| 2020A | CHr01_18590758-CHr01_19419173 | 828.4 | 31.18 | 0.56 | 0.04 | 23.82 | |
| 2020M | CHr01_18713051-CHr01_19638223 | 925.2 | 14.02 | 0.43 | 0.15 | 10.51 | |
| CpML17.1 | 2019A | CHr17_5225091-CHr17_5944838 | 719.7 | 38.70 | 0.63 | 0.03 | 20.80 |
| 2020A | CHr17_5225091-CHr17_5944838 | 719.7 | 20.65 | 0.47 | -0.01 | 16.25 | |
| 2020M | CHr17_5225091-CHr17_5944838 | 719.7 | 52.99 | 0.85 | -0.19 | 38.68 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, K.; Wang, X.; Zhang, L.; Chi, Y.; Luo, Y.; Xu, W.; Wang, Y.; Qu, S. Morphological Analyses and QTL Mapping of Mottled Leaf in Zucchini (Cucurbita pepo L.). Int. J. Mol. Sci. 2024, 25, 2491. https://doi.org/10.3390/ijms25052491
Wang K, Wang X, Zhang L, Chi Y, Luo Y, Xu W, Wang Y, Qu S. Morphological Analyses and QTL Mapping of Mottled Leaf in Zucchini (Cucurbita pepo L.). International Journal of Molecular Sciences. 2024; 25(5):2491. https://doi.org/10.3390/ijms25052491
Chicago/Turabian StyleWang, Kexin, Xinyu Wang, Lijing Zhang, Yichen Chi, Yusong Luo, Wenlong Xu, Yunli Wang, and Shuping Qu. 2024. "Morphological Analyses and QTL Mapping of Mottled Leaf in Zucchini (Cucurbita pepo L.)" International Journal of Molecular Sciences 25, no. 5: 2491. https://doi.org/10.3390/ijms25052491