
Differential Photosensitivity of Fibroblasts Obtained from Normal Skin and Hypertrophic Scar Tissues

Abstract
:1. Introduction
2. Results
2.1. Cellular Characteristics
2.2. Expression of Opsin Types
2.3. Peripheral Circadian Rhythm Formation by BL
2.4. Changes in αSMA Expression and the Role of OPN3 following BL Irradiation
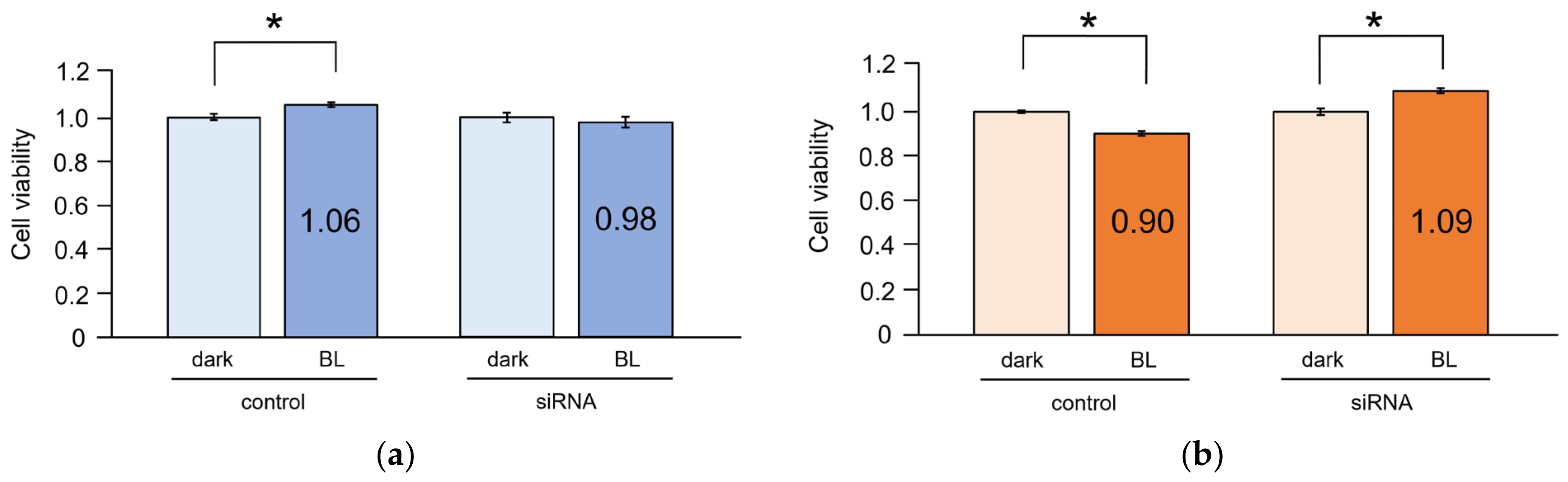
2.5. Cell Viability
3. Discussion
4. Materials and Methods
4.1. Primary Cell Culture
4.2. Total RNA Extraction
4.3. qRT-PCR
4.4. Circadian Rhythm of NDFs and HSFs
4.5. Irradiation
4.6. siRNA Transfection
4.7. Cell Viability
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Niessen, F.B.; Spauwen, P.H.; Schalkwijk, J.; Kon, M. On the nature of hypertrophic scars and keloids: A review. Plast. Reconstr. Surg. 1999, 104, 1435–1458. [Google Scholar] [CrossRef]
- Slemp, A.E.; Kirschner, R. Keloids and scars: A review of keloids and scars, their pathogenesis, risk factors, and management. Curr. Opin. Pediatr. 2006, 18, 396–402. [Google Scholar] [CrossRef]
- Desmouliere, A. Factors influencing myofibroblast differentiation during wound healing and fibrosis. Cell. Biol. Int. 1995, 19, 471–476. [Google Scholar] [CrossRef]
- Gauglitz, G.G.; Korting, H.C.; Pavicic, T.; Ruzicka, T.; Jeschke, M.G. Hypertrophic scarring and keloids: Pathomechanisms and current and emerging treatment strategies. Mol. Med. 2011, 17, 113–125. [Google Scholar] [CrossRef]
- Kim, M.; Juang, H.Y.; Park, H.J. Topical PDT in the treatment of benign skin diseases: Principles and new application. Int. J. Mol. Sci. 2015, 16, 23259–23278. [Google Scholar] [CrossRef]
- Kurz, B.; Berneburg, M.; Bäumler, W.; Karrer, S. Phototherapy: Theory and practice. J. Dtsch. Dermatol. Ges. 2023, 21, 882–897. [Google Scholar] [CrossRef]
- Fushimi, T.; Inui, S.; Nakajima, T.; Ogasawara, M.; Hosokawa, K.; Itami, S. Green light emitting diodes accelerate wound healing: Characterization of the effect and its molecular basis in vitro and in vivo. Wound Repair Regen. 2012, 20, 226–235. [Google Scholar] [CrossRef]
- Sheen, Y.S.; Fan, S.M.; Chan, C.C.; Wu, Y.F.; Jee, S.H.; Lin, S.J. Visible red light enhances physiological anagen entry in vivo and has direct and indirect stimulative effects in vitro. Lasers Surg. Med. 2015, 47, 50–59. [Google Scholar] [CrossRef]
- Uzunbajakava, N.E.; Tobin, D.J.; Botchkareva, N.V.; Dierickx, C.; Bjerring, P.; Town, G. Highlighting nuances of blue light phototherapy: Mechanisms and safety considerations. J. Biophotonics 2023, 16, e202200257. [Google Scholar] [CrossRef]
- Andrabi, M.; Upton, B.A.; Lang, R.A.; Vemaraju, S. An expanding role for nonvisual opsins in extraocular light sensing physiology. Annu. Rev. Vis. Sci. 2023, 9, 245–267. [Google Scholar] [CrossRef]
- Shichida, Y.; Matsuyama, T. Evolution of opsins and phototransduction. Philo. Trans. R Soc. Lond. B Biol. Sci. 2009, 364, 2881–2895. [Google Scholar] [CrossRef]
- Guido, M.E.; Marchese, N.A.; Rios, M.N.; Morera, L.P.; Diaz, N.M.; Garbarino-Pico, E.; Contin, M.A. Non-visual opsins and novel photo-detectors in the vertebrate inner retina mediate light responses within the blue spectrum region. Cell. Mol. Neurobiol. 2022, 42, 59–83. [Google Scholar] [CrossRef]
- Lan, Y.; Wang, Y.; Lu, H. Opsin 3 is a key regulator of ultraviolet A-induced photoageing in human dermal fibroblast cells. Br. J. Dermatol. 2020, 182, 1228–1244. [Google Scholar] [CrossRef]
- Olinski, L.E.; Lin, E.M.; Oancea, E. Illuminating insights into opsin 3 function in the skin. Adv. Biol. Regul. 2020, 75, 100668. [Google Scholar] [CrossRef]
- Kusumoto, J.; Takeo, M.; Hashikawa, K.; Komori, T.; Tsuji, T.; Terashi, H.; Sakakibara, S. OPN4 belongs to the photosensitive system of the human skin. Genes Cells 2020, 25, 215–225. [Google Scholar] [CrossRef]
- Berson, D.M.; Dunn, F.A.; Takao, M. Phototransduction by retinal ganglion cells that set the circadian clock. Science 2002, 295, 1070–1073. [Google Scholar] [CrossRef]
- Ruby, N.F.; Brennan, T.J.; Xie, X.; Cao, V.; Franken, P.; Heller, H.C.; O’Hara, B.F. Role of melanopsin in circadian responses to light. Science 2002, 298, 2211–2213. [Google Scholar] [CrossRef]
- Panda, S.; Sato, T.K.; Castrucci, A.M.; Rollag, M.D.; DeGrip, W.J.; Hogenesch, J.B.; Provencio, I.; Kay, S.A. Melanopsin (Opn4) requirement for normal light-induced circadian phase shifting. Science 2002, 298, 2213–2216. [Google Scholar] [CrossRef]
- Hattar, S.; Liao, H.W.; Takao, M.; Berson, D.M.; Yau, K.W. Melanopsin-containing retinal ganglion cells: Architecture, projections, and intrinsic photosensitivity. Science 2002, 295, 1065–1070. [Google Scholar] [CrossRef]
- Zanello, S.B.; Jackson, D.M.; Holick, M.F. Expression of the circadian clock genes clock and period1 in human skin. J. Investig. Dermatol. 2000, 115, 757–760. [Google Scholar] [CrossRef]
- Nagoshi, E.; Saini, C.; Bauer, C.; Lacroche, T.; Naef, F.; Schibler, U. Circadian gene expression in individual fibroblasts: Cell-autonomous and self-sustained oscillators pass time to daughter cells. Cell 2004, 119, 693–705. [Google Scholar] [CrossRef]
- Reppert, S.M.; Weaver, D.R. Coordination of circadian timing in mammals. Nature 2002, 418, 935–941. [Google Scholar] [CrossRef]
- Cheon, S.; Park, N.; Cho, S.; Kim, K. Glucocorticoid-mediated Period2 induction delays the phase of circadian rhythm. Nucleic Acids Res. 2013, 41, 6161–6174. [Google Scholar] [CrossRef]
- So, A.Y.; Bernal, T.U.; Pillsbury, M.L.; Yamamoto, K.R.; Feldman, B.J. Glucocorticoid regulation of the circadian clock modulates glucose homeostasis. Proc. Natl. Acad. Sci. USA 2009, 106, 17582–17587. [Google Scholar] [CrossRef]
- Matsui, M.S.; Pelle, E.; Dong, K.; Pernodet, N. Biological rhythms in the skin. Int. J. Mol. Sci. 2016, 17, 801. [Google Scholar] [CrossRef]
- Al-Nuaimi, Y.; Hardman, J.A.; Biró, T.; Haslam, I.S.; Philpott, M.P.; Tóth, B.I.; Farjo, N.; Baier, G.; Watson, R.E.B.; Grimaldi, B.; et al. A meeting of two chronobiological systems: Circadian proteins period1 and BMAL1 modulate the human hair cycle clock. J. Investig. Dermatol. 2014, 134, 610–619. [Google Scholar] [CrossRef]
- Stenger, S.; Grasshoff, H.; Hundt, J.E.; Lange, T. Potential effects of shift work on skin autoimmune diseases. Front. Immunol. 2022, 13, 1000951. [Google Scholar] [CrossRef]
- Buhr, E.D.; Vemaraju, S.; Diaz, N.; Lang, R.A.; Van Gelder, R.N. Neuropsin (OPN5) mediates local light-dependent induction of circadian clock genes and circadian photoentrainment in exposed murine skin. Curr. Biol. 2019, 29, 3478–3487. [Google Scholar] [CrossRef]
- Barole, D.; Boucher, A. Prophylactic low-level light therapy for the treatment of hypertrophic scars and keloids: A case series. Lasers Surg. Med. 2010, 42, 597–601. [Google Scholar] [CrossRef]
- Yamazaki, S.; Numano, R.; Abe, M.; Hida, A.; Takahashi, R.; Ueda, M.; Block, G.D.; Sakaki, Y.; Menaker, M.; Tei, H. Resetting central and peripheral circadian oscillators in transgenic rats. Science 2000, 288, 682–685. [Google Scholar] [CrossRef]
- Sandu, C.; Dumas, M.; Malan, A.; Sambakhe, D.; Marteau, C.; Nizard, C.; Schnebert, S.; Perrier, E.; Challet, E.; Pévet, P.; et al. Human skin keratinocytes, melanocytes, and fibroblasts contain distinct circadian clock machineries. Cell. Mol. Life. Sci. 2012, 69, 3329–3339. [Google Scholar] [CrossRef]
- Balsalobre, A.; Brown, S.A.; Marcacci, L.; Tronche, F.; Kekkendonk, C.; Reichardt, H.M.; Schütz, G.; Schibler, U. Resetting of circadian time in peripheral tissues by glucocorticoid signaling. Science 2000, 289, 2344–2347. [Google Scholar] [CrossRef]
- Salazar, A.; von Hagen, J. Circadian oscillations in skin and their interconnection with the cycle of life. Int. J. Mol. Sci. 2023, 24, 5635. [Google Scholar] [CrossRef]
- Geyfman, M.; Kumar, V.; Liu, Q.; Ruiz, R.; Gordon, W.; Espitia, F.; Cam, E.; Millar, S.E.; Smyth, P.; Ihler, A.; et al. Brain and muscle Arnt-like protein-1 (BMAL1) controls circadian cell proliferation and susceptibility to UVB-induced DNA damage in the epidermis. Proc. Natl. Acad. Sci. USA 2012, 109, 11758–11763. [Google Scholar] [CrossRef]
- Luengas-Martinez, A.; Paus, R.; Iqbal, M.; Bailey, L.; Ray, D.W.; Young, H.S. Circadian rhythms in psoriasis and the potential of chronotherapy in psoriasis management. Exp. Dermatol. 2022, 31, 1800–1809. [Google Scholar] [CrossRef]
- Vaughn, A.R.; Clark, A.K.; Sivamani, R.K.; Shi, V.Y. Circadian rhythm in atopic dermatitis-pathophysiology and implications for chronotherapy. Pediatr. Dermatol. 2018, 35, 152–157. [Google Scholar] [CrossRef]
- Yoshimoto, T.; Shimada, M.; Tokunaga, T.; Nakao, T.; Nishi, M.; Takasu, C.; Kashihara, H.; Wada, Y.; Okikawa, S.; Yoshikawa, K. Blue light irradiation inhibits the growth of colon cancer and activation of cancer-associated fibroblasts. Oncol. Rep. 2022, 47, 104. [Google Scholar] [CrossRef]
- Magni, G.; Banchelli, M.; Cherchi, F.; Coppi, E.; Fraccalvieri, M.; Rossi, M.; Tatini, F.; Pugliese, A.M.; Degl’Innocenti, D.R.; Alfieri, D.; et al. Experimental study on blue light interaction with human keloid-derived fibroblasts. Biomedicines 2020, 8, 573. [Google Scholar] [CrossRef]
- Taflinski, L.; Demir, E.; Kauczok, J.; Fuchs, P.C.; Born, M.; Suschek, C.V.; Opländer, C. Blue light inhibits transforming growth factor-1-induced myofibroblast differentiation of human dermal fibroblasts. Exp. Dermatol. 2014, 23, 240–246. [Google Scholar] [CrossRef]
- Krassovka, J.M.; Suschek, C.V.; Prost, M.; Grotheer, V.; Schiefer, J.L.; Demir, E.; Fuchs, P.C.; Windolf, J.; Stürmer, E.K.; Opländer, C. The impact of non-toxic blue light (453nm) on cellular antioxidative capacity, TGF-β1 signaling, and myofibrogenesis of human skin fibroblasts. J. Photochem. Photobiol. B 2020, 209, 111952. [Google Scholar] [CrossRef]
- Opländer, C.; Hidding, S.; Wemers, F.B.; Born, M.; Pallua, N.; Suschek, C.V. Effects of blue light irradiation on human dermal fibroblasts. J. Photochem. Photobiol. B 2011, 103, 118–125. [Google Scholar] [CrossRef]
- Masson-Meyers, D.S.; Bumah, V.V.; Enwemeka, C.S. Blue light does not impair wound healing in vitro. J. Photochem. Photobiol. B 2016, 160, 53–60. [Google Scholar] [CrossRef]
- Espósito, A.C.C.; de Souza, N.P.; Miot, L.D.B.; Miot, H.A. Expression of OPN3 in fibroblasts, melanocytes, and keratinocytes of skin with facial melasma in comparison with unaffected adjacent skin. An. Bras. Dermatol. 2021, 96, 367–369. [Google Scholar] [CrossRef]
- Yu, L.; Ren, L.; Dong, L. BMAL1 plays a critical role in the protection against cardiac hypertrophy through autophagy in vitro. BMC Cardiovasc. Disord. 2022, 22, 381. [Google Scholar] [CrossRef]
- Dong, C.; Gongora, C.; Sosulski, M.L.; Luo, F.; Sanchez, C.G. Regulation of transforming growth factor-beta1 (TGF-β1)-induced pro-fibrotic activities by circadian clock gene BMAL1. Respir. Res. 2016, 17, 4. [Google Scholar] [CrossRef]
- Xu, L.; Yang, T.Y.; Zhou, Y.W.; Wu, M.F.; Shen, J.; Chen, J.L.; Liu, Q.X.; Cao, S.Y.; Wang, J.Q.; Zhang, L. Bmal1 inhibits phenotypic transformation of hepatic stellate cells in liver fibrosis via IDH1/α-KG-mediated glycolysis. Acta Pharmacol. Sin. 2022, 43, 316–329. [Google Scholar] [CrossRef]
- Suitthimeatheforn, O.; Yang, C.; Ma, Y.; Liu, W. Direct and indirect effects of blue light exposure on skin: A review of published literature. Skin Pharmacol. Physiol. 2022, 35, 305–318. [Google Scholar] [CrossRef]
- Duteil, L.; Queille-Roussel, C.; Lacour, J.P.; Montaudie, H.; Passeron, T. Short-term exposure to blue light emitted by electronic devices does not worsen melasma. J. Am. Acad. Dermatol. 2020, 83, 913–914. [Google Scholar] [CrossRef]
- Bumah, V.V.; Masson-Meyers, D.S.; Awosika, O.; Zacharias, S.; Enwemeka, C.S. The viability of human cells irradiated with 470-nm light at various radiant energies in vitro. Lasers Med. Sci. 2021, 36, 1661–1670. [Google Scholar] [CrossRef]
- Ash, C.; Dubec, M.; Donne, K.; Bashford, T. Effect of wavelength and beam width on penetration in light-tissue interaction using computational methods. Lasers Med. Sci. 2017, 32, 1909–1918. [Google Scholar] [CrossRef]
- Ankri, R.; Lubart, R.; Taitelbaum, H. Estimation of the optimal wavelengths for laser-induced wound healing. Lasers Surg. Med. 2010, 42, 760–764. [Google Scholar] [CrossRef] [PubMed]
- Desmoulière, A.; Chaponnier, C.; Gabbiani, G. Tissue repair, contraction, and the myofibroblast. Wound Repair Regen. 2005, 13, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, T.; Morine, Y.; Takasu, C.; Feng, R.; Ikemoto, T.; Yoshikawa, K.; Iwahashi, S.; Saito, Y.; Kashihara, H.; Akutagawa, M.; et al. Blue light-emitting diodes induce autophagy in colon cancer cells by Opsin3. Ann. Gastroenterol. Surg. 2018, 2, 154–161. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Forward (5′-3′) | Reverse (5′-3′) | Probe (5′-3′) |
|---|---|---|---|
| OPN1 | GGCCCTGAAAGCTGTTGCA | GCACGTAGCAGACACAGAAGG | ATCACAACCACCATGCGGCTCACCTC |
| OPN2 | CCGTCAAGGAGGCCGCTG | CACCCAGCAGATCAGGAAAGC | CAGCAGCAGGAGTCAGCCACCACAC |
| OPN3 | GGCAGCCTCTTCGGGATTG | CACTCTGGCATGGACCACG | TTCCATTGCCACCCTAACCGTGCTGG |
| OPN4 | CCCCTGTCTTCTTCACCAGT | GATTACCAGGTAGCGGTCCA | ATAGAACTCGCAGCCTGTCTCCCCAAA |
| OPN5 | CTGCAGCGATGTACAATCCC | GCACAGCAGAAGACTTCCTG | TGCAGCCTGAAGCCTTCCAGAGACTT |
| αSMA | TCCAGGCGGTGCTGTCTC | CTCGGCCAGCCAGATCCA | CCTCTGGACGCACAACTGGCATCGTG |
| Per2 | GGACAGCGTCATCAGGTACTTG | CCGCTTATCACTGGACCTTAGC | CTCGCATTTCCTCTTCAGGGTGGCAGC |
| Bmal1 | CACCAATCCATACACAGAAGCAA | CTTCCCTCGGTCACATCCTAC | TGAAACACCTCATTCTCAGGGCAGCAGATG |
| β-Actin | TTGGCAATGAGCGGTTCCG | GGAGTTGAAGGTAGTTTCGTGGA | CCTTCCTTCCTGGGCATGGAGTCCTGTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kusumoto, J.; Akashi, M.; Terashi, H.; Sakakibara, S. Differential Photosensitivity of Fibroblasts Obtained from Normal Skin and Hypertrophic Scar Tissues. Int. J. Mol. Sci. 2024, 25, 2126. https://doi.org/10.3390/ijms25042126
Kusumoto J, Akashi M, Terashi H, Sakakibara S. Differential Photosensitivity of Fibroblasts Obtained from Normal Skin and Hypertrophic Scar Tissues. International Journal of Molecular Sciences. 2024; 25(4):2126. https://doi.org/10.3390/ijms25042126
Chicago/Turabian StyleKusumoto, Junya, Masaya Akashi, Hiroto Terashi, and Shunsuke Sakakibara. 2024. "Differential Photosensitivity of Fibroblasts Obtained from Normal Skin and Hypertrophic Scar Tissues" International Journal of Molecular Sciences 25, no. 4: 2126. https://doi.org/10.3390/ijms25042126