
Role of microRNAs in Immune Regulation with Translational and Clinical Applications

Abstract
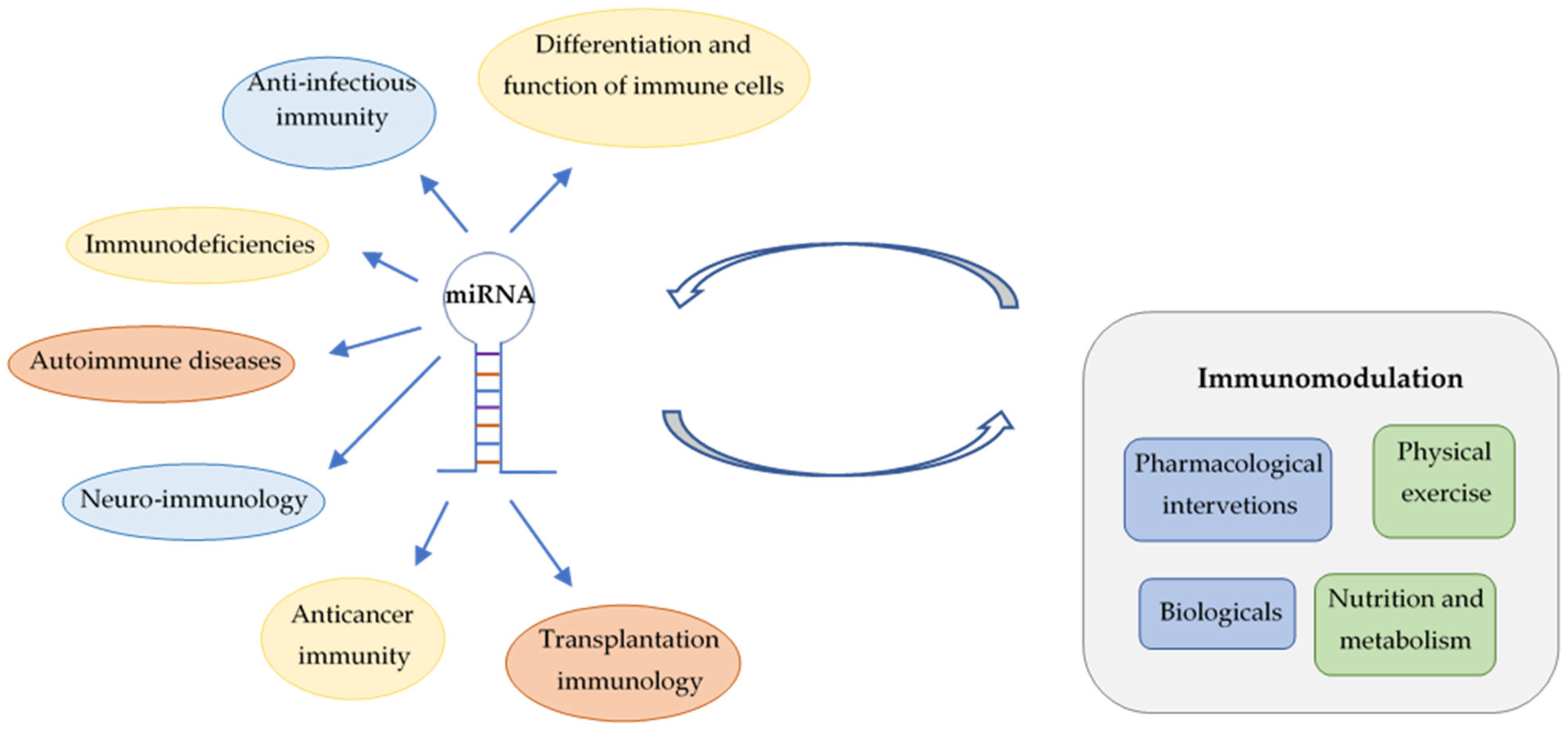
:1. Introduction
2. miRNAs Regulate the Differentiation and Function of Immune Cells
2.1. T Lymphocyte
2.2. Natural Killer Cell
2.3. B Lymphocyte
2.4. Macrophage
2.5. Dendritic Cell
2.6. Neutrophil
2.7. Mast Cell
3. Role of miRNAs in Anti-Infectious Immunity
3.1. Antiviral Immunity
3.2. Immune Response against Bacteria
3.3. miRNA Fingerprints in Fungal Infections
3.4. Antiparasite Immunity
3.5. miRNA Biomarkers in Sepsis
4. Implications of miRNAs in Immunodeficiencies
5. miRNAs as Potential Biomarkers and Therapeutic Targets in Autoimmune Diseases
5.1. Rheumatoid Arthritis
5.2. Systemic Lupus Erythematosus
5.3. Antiphospholipid Syndrome
5.4. Scleroderma and Systemic Sclerosis
5.5. Sjögren’s Syndrome
5.6. Autoimmune Vasculitis
6. Neuro-Immunology and miRNAs
6.1. Multiple Sclerosis
6.2. Myasthenia Gravis
7. Implications of miRNAs in Anticancer Immunity
7.1. Hematological Malignancies
7.2. Solid Tumors
8. Applications of miRNAs in Transplantation Immunology
9. Impact of Immunomodulation on miRNA Fingerprints
9.1. Pharmacological Interventions
9.2. Biologicals
9.3. Physical Exercise
9.4. Metabolism and Nutrition
10. Discussion
11. Concluding Remarks
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, S.; Pan, W.; Qian, Y. MicroRNA in immunity and autoimmunity. J. Mol. Med. 2013, 91, 1039–1050. [Google Scholar] [CrossRef]
- Clayton, S.A.; Jones, S.W.; Kurowska-Stolarska, M.; Clark, A.R. The role of microRNAs in glucocorticoid action. J. Biol. Chem. 2018, 293, 1865–1874. [Google Scholar] [CrossRef] [PubMed]
- Bofill-De Ros, X.; Vang Orom, U.A. Recent progress in miRNA biogenesis and decay. RNA Biol. 2024, 21, 1–8. [Google Scholar] [CrossRef]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, H.; Suzuki, H.I. Systems and Synthetic microRNA Biology: From Biogenesis to Disease Pathogenesis. Int. J. Mol. Sci. 2019, 21, 132. [Google Scholar] [CrossRef]
- Iwakawa, H.O.; Tomari, Y. Life of RISC: Formation, action, and degradation of RNA-induced silencing complex. Mol. Cell 2022, 82, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Morales, S.; Monzo, M.; Navarro, A. Epigenetic regulation mechanisms of microRNA expression. Biomol. Concepts 2017, 8, 203–212. [Google Scholar] [CrossRef]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Plotnikova, O.; Baranova, A.; Skoblov, M. Comprehensive Analysis of Human microRNA-mRNA Interactome. Front. Genet. 2019, 10, 933. [Google Scholar] [CrossRef]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef]
- Wang, Z.; Dai, R.; Ahmed, S.A. MicroRNA-183/96/182 cluster in immunity and autoimmunity. Front. Immunol. 2023, 14, 1134634. [Google Scholar] [CrossRef]
- Chen, C.Z.; Li, L.; Lodish, H.F.; Bartel, D.P. MicroRNAs modulate hematopoietic lineage differentiation. Science 2004, 303, 83–86. [Google Scholar] [CrossRef]
- Sekar, D.; Lakshmanan, G. Methylation of N6-adenosine (m6A) modification in miRNAs and its implications in immunity. Epigenomics 2020, 12, 1083–1085. [Google Scholar] [CrossRef] [PubMed]
- Matias-Garcia, P.R.; Wilson, R.; Mussack, V.; Reischl, E.; Waldenberger, M.; Gieger, C.; Anton, G.; Peters, A.; Kuehn-Steven, A. Impact of long-term storage and freeze-thawing on eight circulating microRNAs in plasma samples. PLoS ONE 2020, 15, e0227648. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Xu, L.; Zhang, R.; Jin, Y.; Jiang, P.; Wei, K.; Xu, L.; Shi, Y.; Zhao, J.; Xiong, M.; et al. MicroRNA-Mediated Epigenetic Regulation of Rheumatoid Arthritis Susceptibility and Pathogenesis. Front. Immunol. 2022, 13, 838884. [Google Scholar] [CrossRef]
- Tong, N.; Liu, D.; Lu, L.; Lin, R.; Jin, R. miR-410 Regulates Helper T Cell Differentiation in Ovalbumin-Induced Asthma through the PI3K-AKT-VEGF Signaling Pathway. Int. Arch. Allergy Immunol. 2023, 185, 1–9. [Google Scholar] [CrossRef]
- Lu, T.X.; Hartner, J.; Lim, E.J.; Fabry, V.; Mingler, M.K.; Cole, E.T.; Orkin, S.H.; Aronow, B.J.; Rothenberg, M.E. MicroRNA-21 limits in vivo immune response-mediated activation of the IL-12/IFN-gamma pathway, Th1 polarization, and the severity of delayed-type hypersensitivity. J. Immunol. 2011, 187, 3362–3373. [Google Scholar] [CrossRef] [PubMed]
- Fu, D.; Yu, W.; Li, M.; Wang, H.; Liu, D.; Song, X.; Li, Z.; Tian, Z. MicroRNA-138 regulates the balance of Th1/Th2 via targeting RUNX3 in psoriasis. Immunol. Lett. 2015, 166, 55–62. [Google Scholar] [CrossRef]
- Simpson, L.J.; Patel, S.; Bhakta, N.R.; Choy, D.F.; Brightbill, H.D.; Ren, X.; Wang, Y.; Pua, H.H.; Baumjohann, D.; Montoya, M.M.; et al. A microRNA upregulated in asthma airway T cells promotes TH2 cytokine production. Nat. Immunol. 2014, 15, 1162–1170. [Google Scholar] [CrossRef]
- Hart, M.; Walch-Ruckheim, B.; Friedmann, K.S.; Rheinheimer, S.; Tanzer, T.; Glombitza, B.; Sester, M.; Lenhof, H.P.; Hoth, M.; Schwarz, E.C.; et al. miR-34a: A new player in the regulation of T cell function by modulation of NF-kappaB signaling. Cell Death Dis. 2019, 10, 46. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, Z.; Li, F.; Ping, Y.; Qin, G.; Zhang, C.; Zhang, Y. miR-143 Regulates Memory T Cell Differentiation by Reprogramming T Cell Metabolism. J. Immunol. 2018, 201, 2165–2175. [Google Scholar] [CrossRef]
- Zhou, S.; Dong, X.; Zhang, C.; Chen, X.; Zhu, J.; Li, W.; Song, X.; Xu, Z.; Zhang, W.; Yang, X.; et al. MicroRNAs are implicated in the suppression of CD4+CD25− conventional T cell proliferation by CD4+CD25+ regulatory T cells. Mol. Immunol. 2015, 63, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Johansson, K.; Gagnon, J.D.; Zhou, S.K.; Fassett, M.S.; Schroeder, A.W.; Kageyama, R.; Bautista, R.A.; Pham, H.; Woodruff, P.G.; Ansel, K.M. An essential role for miR-15/16 in Treg suppression and restriction of proliferation. Cell Rep. 2023, 42, 113298. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Li, G.; Kim, C.; Hu, B.; Jadhav, R.R.; Weyand, C.M.; Goronzy, J.J. Regulation of miR-181a expression in T cell aging. Nat. Commun. 2018, 9, 3060. [Google Scholar] [CrossRef] [PubMed]
- Saultz, J.N.; Freud, A.G.; Mundy-Bosse, B.L. MicroRNA regulation of natural killer cell development and function in leukemia. Mol. Immunol. 2019, 115, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, R.P.; Leong, J.W.; Schneider, S.E.; Ireland, A.R.; Berrien-Elliott, M.M.; Singh, A.; Schappe, T.; Jewell, B.A.; Sexl, V.; Fehniger, T.A. MicroRNA-15/16 Antagonizes Myb To Control NK Cell Maturation. J. Immunol. 2015, 195, 2806–2817. [Google Scholar] [CrossRef] [PubMed]
- Bezman, N.A.; Chakraborty, T.; Bender, T.; Lanier, L.L. miR-150 regulates the development of NK and iNKT cells. J. Exp. Med. 2011, 208, 2717–2731. [Google Scholar] [CrossRef] [PubMed]
- Trotta, R.; Chen, L.; Ciarlariello, D.; Josyula, S.; Mao, C.; Costinean, S.; Yu, L.; Butchar, J.P.; Tridandapani, S.; Croce, C.M.; et al. miR-155 regulates IFN-gamma production in natural killer cells. Blood 2012, 119, 3478–3485. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.D.; Lee, S.U.; Yun, S.; Sun, H.N.; Lee, S.H.; Kim, J.W.; Kim, H.M.; Park, S.K.; Lee, C.W.; Yoon, S.R.; et al. Human microRNA-27a* targets Prf1 and GzmB expression to regulate NK-cell cytotoxicity. Blood 2011, 118, 5476–5486. [Google Scholar] [CrossRef]
- Belver, L.; de Yebenes, V.G.; Ramiro, A.R. MicroRNAs prevent the generation of autoreactive antibodies. Immunity 2010, 33, 713–722. [Google Scholar] [CrossRef]
- Hu, Y.Z.; Li, Q.; Wang, P.F.; Li, X.P.; Hu, Z.L. Multiple functions and regulatory network of miR-150 in B lymphocyte-related diseases. Front. Oncol. 2023, 13, 1140813. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Zhao, J.; Wu, W.; Hao, C.; Yang, Y.; Zhang, J. miR-130b regulates B cell proliferation via CYLD-mediated NF-kappaB signaling. Exp. Cell Res. 2023, 434, 113870. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, R.; Llorian, M.; Varsani-Brown, S.; Chakravarty, P.; Camarillo, J.M.; Barry, D.; George, R.; Blackledge, N.P.; Duddy, G.; Kelleher, N.L.; et al. Epi-microRNA mediated metabolic reprogramming ensures affinity maturation. bioRxiv 2023. [Google Scholar] [CrossRef]
- Roy, S. miRNA in Macrophage Development and Function. Antioxid. Redox Signal 2016, 25, 795–804. [Google Scholar] [CrossRef]
- Khayati, S.; Dehnavi, S.; Sadeghi, M.; Tavakol Afshari, J.; Esmaeili, S.A.; Mohammadi, M. The potential role of miRNA in regulating macrophage polarization. Heliyon 2023, 9, e21615. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, M.; Zhong, M.; Suo, Q.; Lv, K. Expression profiles of miRNAs in polarized macrophages. Int. J. Mol. Med. 2013, 31, 797–802. [Google Scholar] [CrossRef]
- Qu, R.; Peng, Y.; Zhou, M.; Xu, S.; Yin, X.; Qiu, Y.; Liu, B.; Gao, Y.; Bi, H.; Guo, D. MiR-223-3p attenuates M1 macrophage polarization via suppressing the Notch signaling pathway and NLRP3-mediated pyroptosis in experimental autoimmune uveitis. Eur. J. Pharmacol. 2023, 960, 176139. [Google Scholar] [CrossRef]
- Peng, L.Y.; Li, B.B.; Deng, K.B.; Wang, W.G. MicroRNA-214-3p facilitates M2 macrophage polarization by targeting GSK3B. Kaohsiung J. Med. Sci. 2022, 38, 347–356. [Google Scholar] [CrossRef]
- Singh, S.; Henderson, J.; Patil, M.; Dubey, P.K.; Dubey, S.; Kannappan, R.; Zhang, J.; Krishnamurthy, P. MicroRNA-181c-5p modulates phagocytosis efficiency in bone marrow-derived macrophages. Inflamm. Res. 2022, 71, 321–330. [Google Scholar] [CrossRef]
- Palucka, K.; Banchereau, J. Dendritic cells: A link between innate and adaptive immunity. J. Clin. Immunol. 1999, 19, 12–25. [Google Scholar] [CrossRef]
- Cordeiro, B.; Jeon, P.; Boukhaled, G.M.; Corrado, M.; Lapohos, O.; Roy, D.G.; Williams, K.; Jones, R.G.; Gruenheid, S.; Sagan, S.M.; et al. MicroRNA-9 Fine-Tunes Dendritic Cell Function by Suppressing Negative Regulators in a Cell-Type-Specific Manner. Cell Rep. 2020, 31, 107585. [Google Scholar] [CrossRef] [PubMed]
- Smyth, L.A.; Boardman, D.A.; Tung, S.L.; Lechler, R.; Lombardi, G. MicroRNAs affect dendritic cell function and phenotype. Immunology 2015, 144, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Yang, C.; Liu, Y.; Sun, D.; Li, X.; Wei, R.; Nian, H. Mettl3 induced miR-338-3p expression in dendritic cells promotes antigen-specific Th17 cell response via regulation of Dusp16. FASEB J. 2023, 37, e23277. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, J.; Xia, W.; Chen, C.; Zhu, H.; Chen, J.; Li, S.; Su, X.; Qin, X.; Ding, H.; et al. MiR-200b modulates the properties of human monocyte-derived dendritic cells by targeting WASF3. Life Sci. 2015, 122, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Kumar Kingsley, S.M.; Vishnu Bhat, B. Role of MicroRNAs in the development and function of innate immune cells. Int. Rev. Immunol. 2017, 36, 154–175. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Kim, S.E.; Yano, H.; Matsumoto, G.; Ohuchida, R.; Ishikura, Y.; Araki, M.; Araki, K.; Park, S.; Komatsu, T.; et al. MiR-142 Is Required for Staphylococcus aureus Clearance at Skin Wound Sites via Small GTPase-Mediated Regulation of the Neutrophil Actin Cytoskeleton. J. Investig. Dermatol. 2017, 137, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Muraleedharan, C.K.; McClellan, S.A.; Barrett, R.P.; Li, C.; Montenegro, D.; Carion, T.; Berger, E.; Hazlett, L.D.; Xu, S. Inactivation of the miR-183/96/182 Cluster Decreases the Severity of Pseudomonas aeruginosa-Induced Keratitis. Investig. Ophthalmol. Vis. Sci. 2016, 57, 1506–1517. [Google Scholar] [CrossRef] [PubMed]
- Tay, H.L.; Kaiko, G.E.; Plank, M.; Li, J.; Maltby, S.; Essilfie, A.T.; Jarnicki, A.; Yang, M.; Mattes, J.; Hansbro, P.M.; et al. Antagonism of miR-328 increases the antimicrobial function of macrophages and neutrophils and rapid clearance of non-typeable Haemophilus influenzae (NTHi) from infected lung. PLoS Pathog. 2015, 11, e1004549. [Google Scholar] [CrossRef]
- Hsu, A.Y.; Liu, S.; Syahirah, R.; Brasseale, K.A.; Wan, J.; Deng, Q. Inducible overexpression of zebrafish microRNA-722 suppresses chemotaxis of human neutrophil like cells. Mol. Immunol. 2019, 112, 206–214. [Google Scholar] [CrossRef]
- Shefler, I.; Salamon, P.; Mekori, Y.A. MicroRNA Involvement in Allergic and Non-Allergic Mast Cell Activation. Int. J. Mol. Sci. 2019, 20, 2145. [Google Scholar] [CrossRef]
- Monticelli, S.; Ansel, K.M.; Xiao, C.; Socci, N.D.; Krichevsky, A.M.; Thai, T.H.; Rajewsky, N.; Marks, D.S.; Sander, C.; Rajewsky, K.; et al. MicroRNA profiling of the murine hematopoietic system. Genome Biol. 2005, 6, R71. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, J.; Zhang, J.; Zhang, W.; Wu, Z. Characterization of microRNA profile in IgE-mediated mouse BMMCs degranulation. J. Microbiol. Immunol. Infect. 2020, 53, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liu, F.; Wang, J.; Long, M.; Wang, Z. MicroRNA-21-Mediated Inhibition of Mast Cell Degranulation Involved in the Protective Effect of Berberine on 2,4-Dinitrofluorobenzene-Induced Allergic Contact Dermatitis in Rats via p38 Pathway. Inflammation 2018, 41, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Zhen, S.; Yu, Q.; Gong, Z. HCV-E2 inhibits hepatocellular carcinoma metastasis by stimulating mast cells to secrete exosomal shuttle microRNAs. Oncol. Lett. 2017, 14, 2141–2146. [Google Scholar] [CrossRef] [PubMed]
- Entwistle, L.J.; Wilson, M.S. MicroRNA-mediated regulation of immune responses to intestinal helminth infections. Parasite Immunol. 2017, 39, e12406. [Google Scholar] [CrossRef]
- Kimura, M.; Kothari, S.; Gohir, W.; Camargo, J.F.; Husain, S. MicroRNAs in infectious diseases: Potential diagnostic biomarkers and therapeutic targets. Clin. Microbiol. Rev. 2023, 36, e00015-23. [Google Scholar] [CrossRef]
- Kohno, T.; Tsuge, M.; Murakami, E.; Hiraga, N.; Abe, H.; Miki, D.; Imamura, M.; Ochi, H.; Hayes, C.N.; Chayama, K. Human microRNA hsa-miR-1231 suppresses hepatitis B virus replication by targeting core mRNA. J. Viral Hepat. 2014, 21, e89–e97. [Google Scholar] [CrossRef]
- Lanford, R.E.; Hildebrandt-Eriksen, E.S.; Petri, A.; Persson, R.; Lindow, M.; Munk, M.E.; Kauppinen, S.; Orum, H. Therapeutic silencing of microRNA-122 in primates with chronic hepatitis C virus infection. Science 2010, 327, 198–201. [Google Scholar] [CrossRef]
- Ong, J.W.J.; Tan, K.S.; Lee, J.J.X.; Seet, J.E.; Choi, H.W.; Ler, S.G.; Gunaratne, J.; Narasaraju, T.; Sham, L.T.; Patzel, V.; et al. Differential effects of microRNAs miR-21, miR-99 and miR-145 on lung regeneration and inflammation during recovery from influenza pneumonia. J. Med. Virol. 2023, 95, e29286. [Google Scholar] [CrossRef]
- Martinez-Espinoza, I.; Bungwon, A.D.; Guerrero-Plata, A. Human Metapneumovirus-Induced Host microRNA Expression Impairs the Interferon Response in Macrophages and Epithelial Cells. Viruses 2023, 15, 2272. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Wang, C.; Kessler, P.; Sen, G.C. Herpes simplex virus 1 evades cellular antiviral response by inducing microRNA-24, which attenuates STING synthesis. PLoS Pathog. 2021, 17, e1009950. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Sun, T.; Zhang, M.; Cheng, J.; Gu, J.; Huang, L.; Xiao, M.; Zhou, J.; Luo, H. EBV-Encoded MicroRNA-BART17-3p Targets DDX3X and Promotes EBV Infection in EBV-Associated T/Natural Killer-Cell Lymphoproliferative Diseases. Open Forum Infect. Dis. 2023, 10, ofad516. [Google Scholar] [CrossRef] [PubMed]
- Polonio, C.M.; da Silva, P.; Russo, F.B.; Hyppolito, B.R.N.; Zanluqui, N.G.; Benazzato, C.; Beltrao-Braga, P.C.B.; Muxel, S.M.; Peron, J.P.S. microRNAs Control Antiviral Immune Response, Cell Death and Chemotaxis Pathways in Human Neuronal Precursor Cells (NPCs) during Zika Virus Infection. Int. J. Mol. Sci. 2022, 23, 10282. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Yin, W.; Zhang, X.; Lu, X.; Wu, N. hsa-miR-181-5p inhibits human immunodeficiency virus type 1 replication by downregulating DDX3X expression. Virology 2023, 587, 109868. [Google Scholar] [CrossRef] [PubMed]
- Haroun, R.A.; Osman, W.H.; Eessa, A.M. Prognostic significance of serum miR-18a-5p in severe COVID-19 Egyptian patients. J. Genet. Eng. Biotechnol. 2023, 21, 114. [Google Scholar] [CrossRef] [PubMed]
- Giavina-Bianchi, P.; Cua, E.; Risso, K.; Mondain, V.; Vissian, A.; Joie, C.; Pouletty, P.; Gineste, P.; Ehrlich, H.J.; Kalil, J. ABX464 (obefazimod) for patients with COVID-19 at risk for severe disease: miR-AGE, a randomized, double-blind placebo-controlled trial. J. Allergy Clin. Immunol. Glob. 2023, 2, 100140. [Google Scholar] [CrossRef]
- Guiot, J.; Henket, M.; Remacle, C.; Cambier, M.; Struman, I.; Winandy, M.; Moermans, C.; Louis, E.; Malaise, M.; Ribbens, C.; et al. Systematic review of overlapping microRNA patterns in COVID-19 and idiopathic pulmonary fibrosis. Respir. Res. 2023, 24, 112. [Google Scholar] [CrossRef]
- Abdalla, A.E.; Alanazi, A.; Abosalif, K.O.A.; Alameen, A.A.M.; Junaid, K.; Manni, E.; Talha, A.A.; Ejaz, H. MicroRNA-155, a double-blade sword regulator of innate tuberculosis immunity. Microb. Pathog. 2023, 185, 106438. [Google Scholar] [CrossRef]
- Deng, F.; Xu, P.; Miao, J.; Jin, C.; Tu, H.; Zhang, J. Pulmonary tuberculosis biomarker miR-215-5p inhibits autophagosome-lysosome fusion in macrophages. Tuberculosis 2023, 143, 102422. [Google Scholar] [CrossRef]
- Wang, Z.; Kong, L.C.; Jia, B.Y.; Chen, J.R.; Dong, Y.; Jiang, X.Y.; Ma, H.X. Analysis of the microRNA Expression Profile of Bovine Monocyte-derived Macrophages Infected with Mycobacterium avium subsp. Paratuberculosis Reveals that miR-150 Suppresses Cell Apoptosis by Targeting PDCD4. Int. J. Mol. Sci. 2019, 20, 2708. [Google Scholar] [CrossRef] [PubMed]
- Goekeri, C.; Pennitz, P.; Groenewald, W.; Behrendt, U.; Kirsten, H.; Zobel, C.M.; Berger, S.; Heinz, G.A.; Mashreghi, M.F.; Wienhold, S.M.; et al. MicroRNA-223 Dampens Pulmonary Inflammation during Pneumococcal Pneumonia. Cells 2023, 12, 959. [Google Scholar] [CrossRef]
- Liu, M.; John, C.M.; Jarvis, G.A. Induction of endotoxin tolerance by pathogenic Neisseria is correlated with the inflammatory potential of lipooligosaccharides and regulated by microRNA-146a. J. Immunol. 2014, 192, 1768–1777. [Google Scholar] [CrossRef] [PubMed]
- Jentho, E.; Bodden, M.; Schulz, C.; Jung, A.L.; Seidel, K.; Schmeck, B.; Bertrams, W. microRNA-125a-3p is regulated by MyD88 in Legionella pneumophila infection and targets NTAN1. PLoS ONE 2017, 12, e0176204. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.J.; Chen, X.C.; Xu, L.; Rui, X.H.; Wan, L.; Lu, J.; Liu, J.; Pei, H. MiR-181a-5p Delivered by Adipose-Derived Mesenchymal Stem Cell Exosomes Alleviates Klebsiella pneumonia Infection-Induced Lung Injury by Targeting STAT3 Signaling. Mediators Inflamm. 2022, 2022, 5188895. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Wang, L.; Sun, C.; Yang, L.; Sun, W.; Peng, Q. MicroRNA-125b-5p suppresses Brucella abortus intracellular survival via control of A20 expression. BMC Microbiol. 2016, 16, 171. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Gong, Y.; Lin, X.; Lin, Q.; Luo, J.; Yu, T.; Xu, J.; Chen, L.; Xu, L.; Hu, Y. Down-regulation of microRNA-155 suppressed Candida albicans induced acute lung injury by activating SOCS1 and inhibiting inflammation response. J. Microbiol. 2022, 60, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, S.A.; Fatima, N.; Syed, N.I.; Wu, X.; Yang, X.F.; Chen, J.Y. MicroRNA Expression Profiling of Human Respiratory Epithelium Affected by Invasive Candida Infection. PLoS ONE 2015, 10, e0136454. [Google Scholar] [CrossRef] [PubMed]
- Arghavan, B.; Sharifi, M.; Shafiee, M.; Mohammadi, R. Evaluation of miR-146a expression level in macrophages exposed to Candida glabrata. Curr. Med. Mycol. 2016, 2, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Tolnai, E.; Fidler, G.; Szasz, R.; Rejto, L.; Nwozor, K.O.; Biro, S.; Paholcsek, M. Free circulating mircoRNAs support the diagnosis of invasive aspergillosis in patients with hematologic malignancies and neutropenia. Sci. Rep. 2020, 10, 16532. [Google Scholar] [CrossRef]
- Hart, M.; Nickl, L.; Walch-Rueckheim, B.; Krammes, L.; Rheinheimer, S.; Diener, C.; Taenzer, T.; Kehl, T.; Sester, M.; Lenhof, H.P.; et al. Wrinkle in the plan: miR-34a-5p impacts chemokine signaling by modulating CXCL10/CXCL11/CXCR3-axis in CD4+, CD8+ T cells, and M1 macrophages. J. Immunother. Cancer 2020, 8, e001617. [Google Scholar] [CrossRef]
- Jha, B.K.; Varikuti, S.; Seidler, G.R.; Volpedo, G.; Satoskar, A.R.; McGwire, B.S. MicroRNA-155 Deficiency Exacerbates Trypanosoma cruzi Infection. Infect. Immun. 2020, 88, 10–1128. [Google Scholar] [CrossRef]
- Geraci, N.S.; Tan, J.C.; McDowell, M.A. Characterization of microRNA expression profiles in Leishmania-infected human phagocytes. Parasite Immunol. 2015, 37, 43–51. [Google Scholar] [CrossRef]
- Hong, Y.; Fu, Z.; Cao, X.; Lin, J. Changes in microRNA expression in response to Schistosoma japonicum infection. Parasite Immunol. 2017, 39, e12416. [Google Scholar] [CrossRef]
- El-Assaad, F.; Hempel, C.; Combes, V.; Mitchell, A.J.; Ball, H.J.; Kurtzhals, J.A.; Hunt, N.H.; Mathys, J.M.; Grau, G.E. Differential microRNA expression in experimental cerebral and noncerebral malaria. Infect. Immun. 2011, 79, 2379–2384. [Google Scholar] [CrossRef] [PubMed]
- Jafarzadeh, A.; Naseri, A.; Shojaie, L.; Nemati, M.; Jafarzadeh, S.; Bannazadeh Baghi, H.; Hamblin, M.R.; Akhlagh, S.A.; Mirzaei, H. MicroRNA-155 and antiviral immune responses. Int. Immunopharmacol. 2021, 101, 108188. [Google Scholar] [CrossRef] [PubMed]
- Pawar, P.; Gokavi, J.; Wakhare, S.; Bagul, R.; Ghule, U.; Khan, I.; Ganu, V.; Mukherjee, A.; Shete, A.; Rao, A.; et al. MiR-155 Negatively Regulates Anti-Viral Innate Responses among HIV-Infected Progressors. Viruses 2023, 15, 2206. [Google Scholar] [CrossRef] [PubMed]
- Soltane, R.; Almulla, N.; Alasiri, A.; Elashmawy, N.F.; Qumsani, A.T.; Alshehrei, F.M.; Keshek, D.E.; Alqadi, T.; Al-Ghamdi, S.B.; Allayeh, A.K. A Comparative Analysis of MicroRNA Expression in Mild, Moderate, and Severe COVID-19: Insights from Urine, Serum, and Nasopharyngeal Samples. Biomolecules 2023, 13, 1681. [Google Scholar] [CrossRef] [PubMed]
- Giannella, A.; Riccetti, S.; Sinigaglia, A.; Piubelli, C.; Razzaboni, E.; Di Battista, P.; Agostini, M.; Dal Molin, E.; Manganelli, R.; Gobbi, F.; et al. Circulating microRNA signatures associated with disease severity and outcome in COVID-19 patients. Front. Immunol. 2022, 13, 968991. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Krishnamoorthy, P.; Kumar, H. MicroRNA-30e-5p Regulates SOCS1 and SOCS3 During Bacterial Infection. Front. Cell Infect. Microbiol. 2020, 10, 604016. [Google Scholar] [CrossRef] [PubMed]
- Hosseinipour, M.; Shahbazi, S.; Roudbar-Mohammadi, S.; Khorasani, M.; Marjani, M. Differential genes expression analysis of invasive aspergillosis: A bioinformatics study based on mRNA/microRNA. Mol. Biol. Res. Commun. 2020, 9, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Reina Ortiz, M.; Schreiber, F.; Benitez, S.; Broncano, N.; Chico, M.E.; Vaca, M.; Alexander, N.; Lewis, D.J.; Dougan, G.; Cooper, P.J. Effects of chronic ascariasis and trichuriasis on cytokine production and gene expression in human blood: A cross-sectional study. PLoS Negl. Trop. Dis. 2011, 5, e1157. [Google Scholar] [CrossRef]
- Wang, C.; Zheng, Y.; Fan, Q.; Li, Z.; Qi, X.; Chen, F.; Xu, L.; Zhou, S.; Chen, X.; Li, Y.; et al. MiR-155 promotes acute organ injury in LPS-induced endotoxemic mice by enhancing CCL-2 expression in macrophages. Shock 2023. [Google Scholar] [CrossRef]
- Tao, Y.; Xu, X.; Yang, B.; Zhao, H.; Li, Y. Mitigation of Sepsis-Induced Acute Lung Injury by BMSC-Derived Exosomal miR-125b-5p Through STAT3-Mediated Suppression of Macrophage Pyroptosis. Int. J. Nanomed. 2023, 18, 7095–7113. [Google Scholar] [CrossRef]
- Wu, P.; Kong, L.; Li, J. MicroRNA-494-3p protects rat cardiomyocytes against septic shock via PTEN. Exp. Ther. Med. 2019, 17, 1706–1716. [Google Scholar] [CrossRef]
- Zhao, Y.; Lu, C.; Zhu, R.; Hu, X. Study on identification of a three-microRNA panel in serum for diagnosing neonatal early onset sepsis. J. Matern. Fetal Neonatal Med. 2023, 36, 2280527. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Lui, K.Y.; Hu, X.; Liu, E.; Zhang, T.; Tong, L.; Xu, J.; Huang, F.; Zhu, Y.; Lu, M.; et al. Circulating microRNAs as novel diagnostic biomarkers and prognostic predictors for septic patients. Infect. Genet. Evol. 2021, 95, 105082. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Sun, M.; Zhu, J.H. Clinical role of serum microRNA-155 in early diagnosis and prognosis of septic patients with acute kidney injury. Int. Urol. Nephrol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Hamidi Esfahani, Z.; Yazdani, R.; Shahkarami, S.; Babaha, F.; Abolhassani, H.; Sadr, M.; Pourfathollah, A.A.; Aghamohammadi, A. Evaluation of MicroRNA-125b-5p and Transcription Factors BLIMP1 and IRF4 Expression in Unsolved Common Variable Immunodeficiency Patients. Iran. J. Allergy Asthma Immunol. 2021, 20, 700–710. [Google Scholar] [CrossRef] [PubMed]
- Babaha, F.; Yazdani, R.; Shahkarami, S.; Esfahani, Z.H.; Abolhahassani, H.; Sadr, M.; Hosseini, A.Z.; Aghamohammadi, A. Evaluation of miR-210 expression in common variable immunodeficiency: Patients with unsolved genetic defect. Allergol. Immunopathol. 2021, 49, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Amato, G.; Vita, F.; Quattrocchi, P.; Minciullo, P.L.; Pioggia, G.; Gangemi, S. Involvement of miR-142 and miR-155 in Non-Infectious Complications of CVID. Molecules 2020, 25, 4760. [Google Scholar] [CrossRef]
- Barna, B.P.; McPeek, M.; Malur, A.; Fessler, M.B.; Wingard, C.J.; Dobbs, L.; Verbanac, K.M.; Bowling, M.; Judson, M.A.; Thomassen, M.J. Elevated MicroRNA-33 in Sarcoidosis and a Carbon Nanotube Model of Chronic Granulomatous Disease. Am. J. Respir. Cell Mol. Biol. 2016, 54, 865–871. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Noguchi, Y.; Shindo, M.; Morita, Y.; Oda, Y.; Yoshida, E.; Hamada, H.; Harada, M.; Shiokawa, Y.; Nishida, T.; et al. Regulation of human autoimmune regulator (AIRE) gene translation by miR-220b. Gene 2013, 530, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Sumegi, J.; Nestheide, S.; Aronow, B.; Fletcher, D.; Keddache, M.; Villanueva, J.; Zhang, K.; Filipovich, A.H. MicroRNA activation signature in patients with hemophagocytic lymphohistiocytosis and reversibility with disease-specific therapy. J. Allergy Clin. Immunol. 2016, 137, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Xie, Z.; Gao, L.; Liu, C.; Ai, J.; Zhang, L.; Shen, K. Profiling of EBV-Encoded microRNAs in EBV-Associated Hemophagocytic Lymphohistiocytosis. Tohoku J. Exp. Med. 2015, 237, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Wu, X.X.; Wang, X.F.; Di, D.S.; Huang, Q.; Liu, R.S.; Shuai, Z.W.; Ye, D.Q.; Leng, R.X. Genetic variant in microRNA-146a gene is associated with risk of rheumatoid arthritis. Ann. Med. 2021, 53, 824–829. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Gu, J.; Wang, Y.; Guo, L.; Xie, J.; Yang, M. TNF-alpha stimulated exosome derived from fibroblast-like synoviocytes isolated from rheumatoid arthritis patients promotes HUVEC migration, invasion and angiogenesis by targeting the miR-200a-3p/KLF6/VEGFA axis. Autoimmunity 2023, 56, 2282939. [Google Scholar] [CrossRef] [PubMed]
- Hammad, R.; Eldosoky, M.A.; Lambert, C.; Sack, U.; Kujumdshiev, S.; Abd Elhamed, S.S.; Elfishawi, S.; Mohamed, E.F.; Kandeel, E.Z.; Lotfy, A.W.; et al. Hsa-miR-21-5p reflects synovitis and tenosynovitis components of musculoskeletal ultrasonography Seven-joint scores in rheumatoid arthritis disease and predicts the disease flare. Pathol. Res. Pract. 2023, 253, 154960. [Google Scholar] [CrossRef] [PubMed]
- Kurowska, W.; Kuca-Warnawin, E.; Radzikowska, A.; Jakubaszek, M.; Maslinska, M.; Kwiatkowska, B.; Maslinski, W. Monocyte-related biomarkers of rheumatoid arthritis development in undifferentiated arthritis patients—A pilot study. Reumatologia 2018, 56, 10–16. [Google Scholar] [CrossRef]
- Ciechomska, M.; Roszkowski, L.; Burakowski, T.; Massalska, M.; Felis-Giemza, A.; Roura, A.J. Circulating miRNA-19b as a biomarker of disease progression and treatment response to baricitinib in rheumatoid arthritis patients through miRNA profiling of monocytes. Front. Immunol. 2023, 14, 980247. [Google Scholar] [CrossRef]
- Castro-Villegas, C.; Perez-Sanchez, C.; Escudero, A.; Filipescu, I.; Verdu, M.; Ruiz-Limon, P.; Aguirre, M.A.; Jimenez-Gomez, Y.; Font, P.; Rodriguez-Ariza, A.; et al. Circulating miRNAs as potential biomarkers of therapy effectiveness in rheumatoid arthritis patients treated with anti-TNFalpha. Arthritis Res. Ther. 2015, 17, 49. [Google Scholar] [CrossRef]
- Bluml, S.; Bonelli, M.; Niederreiter, B.; Puchner, A.; Mayr, G.; Hayer, S.; Koenders, M.I.; van den Berg, W.B.; Smolen, J.; Redlich, K. Essential role of microRNA-155 in the pathogenesis of autoimmune arthritis in mice. Arthritis Rheum. 2011, 63, 1281–1288. [Google Scholar] [CrossRef]
- Zhang, S.; Meng, T.; Tang, C.; Li, S.; Cai, X.; Wang, D.; Chen, M. MicroRNA-340-5p suppressed rheumatoid arthritis synovial fibroblast proliferation and induces apoptotic cell number by targeting signal transducers and activators of transcription 3. Autoimmunity 2020, 53, 314–322. [Google Scholar] [CrossRef]
- Abdel-Gawad, A.R.; Shaheen, S.; Babteen, N.A.; Toraih, E.A.; Elshazli, R.M.; Fawzy, M.S.; Gouda, N.S. Association of microRNA 17 host gene variant (rs4284505) with susceptibility and severity of systemic lupus erythematosus. Immun. Inflamm. Dis. 2020, 8, 595–604. [Google Scholar] [CrossRef]
- Tang, Y.; Luo, X.; Cui, H.; Ni, X.; Yuan, M.; Guo, Y.; Huang, X.; Zhou, H.; de Vries, N.; Tak, P.P.; et al. MicroRNA-146A contributes to abnormal activation of the type I interferon pathway in human lupus by targeting the key signaling proteins. Arthritis Rheum. 2009, 60, 1065–1075. [Google Scholar] [CrossRef] [PubMed]
- Xiang, M.; Wang, Y.; Chen, Q.; Wang, J.; Gao, Z.; Liang, J.; Xu, J. LncRNA NEAT1 promotes IL-6 secretion in monocyte-derived dendritic cells via sponging miR-365a-3p in systemic lupus erythematosus. Epigenetics 2023, 18, 2226492. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Li, S.; Jia, Y.; Wu, Q.; Chen, X.; Zhang, M.; Miao, Q.; Zhong, Z.; Zhai, Z.; Ni, B.; et al. CircPTPN22 modulates T-cell activation by sponging miR-4689 to regulate S1PR1 expression in patients with systemic lupus erythematosus. Arthritis Res. Ther. 2023, 25, 206. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.R.K.; Waly, N.G.; Moness, H.; Ahmed, S.S.; Ibrahem, R. Serum miRNA-21, miRNA-146a and plasma cell free DNA as novel biomarkers for assessing systemic lupus erythematosus activity. Mol. Biol. Rep. 2023, 50, 10025–10036. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, R.; Wang, D.; Zhang, S.; Song, H.; Ding, S.; Zhu, Y.; Wen, X.; Li, H.; Chen, H.; et al. Circulating miR-320b Contributes to CD4+ T-Cell Proliferation in Systemic Lupus Erythematosus via MAP3K1. J. Immunol. Res. 2023, 2023, 6696967. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Jiang, L.; Wang, B.; Hu, Q.; Deng, S.; Huang, J.; Sun, X.; Zhang, Y.; Feng, L.; Chen, W. Novel microRNA biomarkers of systemic lupus erythematosus in plasma: miR-124-3p and miR-377-3p. Clin. Biochem. 2022, 107, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Maksoud, R.S.; Rashad, N.M.; Elsayed, W.S.H.; Ali, M.A.; Kamal, N.M.; Zidan, H.E. Circulating miR-181a and miR-223 expression with the potential value of biomarkers for the diagnosis of systemic lupus erythematosus and predicting lupus nephritis. J. Gene Med. 2021, 23, e3326. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Qin, Y.; Shi, M.; Yan, L. Diagnostic significance of circulating miR-485-5p in patients with lupus nephritis and its predictive value evaluation for the clinical outcomes. J. Chin. Med. Assoc. 2021, 84, 491–497. [Google Scholar] [CrossRef]
- Wu, X.N.; Ye, Y.X.; Niu, J.W.; Li, Y.; Li, X.; You, X.; Chen, H.; Zhao, L.D.; Zeng, X.F.; Zhang, F.C.; et al. Defective PTEN regulation contributes to B cell hyperresponsiveness in systemic lupus erythematosus. Sci. Transl. Med. 2014, 6, 246ra99. [Google Scholar] [CrossRef]
- Guo, A.; Sun, Y.; Xu, X.; Xing, Q. MicroRNA-30a Targets Notch1 to Alleviate Podocyte Injury in Lupus Nephritis. Immunol. Investig. 2022, 51, 1694–1706. [Google Scholar] [CrossRef]
- Chai, F.; Peng, H.; Qin, L.; Liu, C.; Zeng, Y.; Wang, R.; Xu, G.; Wang, R.; Wei, G.; Huang, H.; et al. MicroRNA miR-181d-5p regulates the MAPK signaling pathway by targeting mitogen-activated protein kinase 8 (MAPK8) to improve lupus nephritis. Gene 2024, 893, 147961. [Google Scholar] [CrossRef]
- Huang, J.; Xu, X.; Wang, X.; Yang, J.; Xue, M.; Yang, Y.; Zhang, R.; Yang, X.; Yang, J. MicroRNA-590-3p inhibits T helper 17 cells and ameliorates inflammation in lupus mice. Immunology 2022, 165, 260–273. [Google Scholar] [CrossRef] [PubMed]
- Sole, C.; Royo, M.; Sandoval, S.; Moline, T.; Cortes-Hernandez, J. Small-Extracellular-Vesicle-Derived miRNA Profile Identifies miR-483-3p and miR-326 as Regulators in the Pathogenesis of Antiphospholipid Syndrome (APS). Int. J. Mol. Sci. 2023, 24, 11607. [Google Scholar] [CrossRef] [PubMed]
- Juarez-Vicuna, Y.; Guzman-Martin, C.A.; Martinez-Martinez, L.A.; Hernandez-Diazcouder, A.; Huesca-Gomez, C.; Gamboa, R.; Amezcua-Guerra, L.M.; Chacon-Perez, M.; Amigo, M.C.; Sanchez-Munoz, F. miR-19b-3p and miR-20a-5p are associated with the levels of antiphospholipid antibodies in patients with antiphospholipid syndrome. Rheumatol. Int. 2021, 41, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Wang, Y.; Tian, W.; Liu, Y.; Gu, M.; Jiang, X.; Cai, Y.; Huo, R.; Li, Y.; Li, L.; et al. Exosomal miR-146a-5p derived from human umbilical cord mesenchymal stem cells can alleviate antiphospholipid antibody-induced trophoblast injury and placental dysfunction by regulating the TRAF6/NF-kappaB axis. J. Nanobiotechnol. 2023, 21, 419. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Luo, H.; Li, Y.; Zhou, Y.; Jiang, Y.; Chai, J.; Xiao, X.; You, Y.; Zuo, X. MicroRNA-21 in scleroderma fibrosis and its function in TGF-beta-regulated fibrosis-related genes expression. J. Clin. Immunol. 2013, 33, 1100–1109. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, J.; Kahaleh, B. Epigenetic down-regulation of microRNA-126 in scleroderma endothelial cells is associated with impaired responses to VEGF and defective angiogenesis. J. Cell Mol. Med. 2021, 25, 7078–7088. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Lv, Q.; Guo, K.; Li, L.; Zhang, H.; Bian, H. Th17 cell-derived miR-155-5p modulates interleukin-17 and suppressor of cytokines signaling 1 expression during the progression of systemic sclerosis. J. Clin. Lab. Anal. 2022, 36, e24489. [Google Scholar] [CrossRef]
- Shumnalieva, R.; Kachakova, D.; Kaneva, R.; Kolarov, Z.; Monov, S. Serum miR-21 and miR-29a expression in systemic sclerosis patients. Clin. Exp. Rheumatol. 2023, 41, 1688–1694. [Google Scholar] [CrossRef]
- Bayati, P.; Kalantari, M.; Assarehzadegan, M.A.; Poormoghim, H.; Mojtabavi, N. MiR-27a as a diagnostic biomarker and potential therapeutic target in systemic sclerosis. Sci. Rep. 2022, 12, 18932. [Google Scholar] [CrossRef] [PubMed]
- Bayati, P.; Poormoghim, H.; Mojtabavi, N. Aberrant expression of miR-138 as a novel diagnostic biomarker in systemic sclerosis. Biomark. Insights 2022, 17, 11772719221135442. [Google Scholar] [CrossRef] [PubMed]
- Christmann, R.B.; Wooten, A.; Sampaio-Barros, P.; Borges, C.L.; Carvalho, C.R.; Kairalla, R.A.; Feghali-Bostwick, C.; Ziemek, J.; Mei, Y.; Goummih, S.; et al. miR-155 in the progression of lung fibrosis in systemic sclerosis. Arthritis Res. Ther. 2016, 18, 155. [Google Scholar] [CrossRef] [PubMed]
- Wolska-Gawron, K.; Bartosinska, J.; Rusek, M.; Kowal, M.; Raczkiewicz, D.; Krasowska, D. Circulating miRNA-181b-5p, miRNA-223-3p, miRNA-210-3p, let 7i-5p, miRNA-21-5p and miRNA-29a-3p in patients with localized scleroderma as potential biomarkers. Sci. Rep. 2020, 10, 20218. [Google Scholar] [CrossRef]
- Cheng, Q.; Chen, M.; Wang, H.; Chen, X.; Wu, H.; Du, Y.; Xue, J. MicroRNA-27a-3p inhibits lung and skin fibrosis of systemic sclerosis by negatively regulating SPP1. Genomics 2022, 114, 110391. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Long, X.; Mo, M.; Jiang, J.; Zhang, Q.; Long, M.; Li, M. Bone marrow mesenchymal stem cell-derived exosomes alleviate skin fibrosis in systemic sclerosis by inhibiting the IL-33/ST2 axis via the delivery of microRNA-214. Mol. Immunol. 2023, 157, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Castro, I.; Carvajal, P.; Jara, D.; Aguilera, S.; Heathcote, B.; Barrera, M.J.; Aliaga-Tobar, V.; Maracaja-Coutinho, V.; Urzua, U.; Quest, A.F.G.; et al. Small RNA Expression Profiling Reveals hsa-miR-181d-5p Downregulation Associated with TNF-alpha Overexpression in Sjogren’s Syndrome Patients. Front. Immunol. 2022, 13, 870094. [Google Scholar] [CrossRef]
- Qi, X.; Wang, R.; Jin, L.; Tian, Y.; Jin, H.; Han, Y.; Sun, C.; Ding, M.; Guo, H. MiR-223-3p Aggravates Ocular Inflammation in Sjogren’s Syndrome. Endocr. Metab. Immune Disord. Drug Targets 2023, 23, 1087–1095. [Google Scholar] [CrossRef]
- Wang-Renault, S.F.; Boudaoud, S.; Nocturne, G.; Roche, E.; Sigrist, N.; Daviaud, C.; Bugge Tinggaard, A.; Renault, V.; Deleuze, J.F.; Mariette, X.; et al. Deregulation of microRNA expression in purified T and B lymphocytes from patients with primary Sjogren’s syndrome. Ann. Rheum. Dis. 2018, 77, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Kakan, S.S.; Edman, M.C.; Yao, A.; Okamoto, C.T.; Nguyen, A.; Hjelm, B.E.; Hamm-Alvarez, S.F. Tear miRNAs Identified in a Murine Model of Sjogren’s Syndrome as Potential Diagnostic Biomarkers and Indicators of Disease Mechanism. Front. Immunol. 2022, 13, 833254. [Google Scholar] [CrossRef] [PubMed]
- Carvajal, P.; Aguilera, S.; Jara, D.; Indo, S.; Barrera, M.J.; Gonzalez, S.; Molina, C.; Heathcote, B.; Hermoso, M.; Castro, I.; et al. hsa-miR-424-5p and hsa-miR-513c-3p dysregulation mediated by IFN-gamma is associated with salivary gland dysfunction in Sjogren’s syndrome patients. J. Autoimmun. 2023, 138, 103037. [Google Scholar] [CrossRef] [PubMed]
- Yamashiro, K.; Hamada, T.; Mori, K.; Nishi, K.; Nakamura, M.; Beppu, M.; Tanaka, A.; Hijioka, H.; Kamikawa, Y.; Sugiura, T. Exosome-Derived microRNAs from Mouthrinse Have the Potential to Be Novel Biomarkers for Sjogren Syndrome. J. Pers. Med. 2022, 12, 1483. [Google Scholar] [CrossRef] [PubMed]
- Yan, T.; Shen, J.; Chen, J.; Zhao, M.; Guo, H.; Wang, Y. Differential expression of miR-17-92 cluster among varying histological stages of minor salivary gland in patients with primary Sjogren’s syndrome. Clin. Exp. Rheumatol. 2019, 37 (Suppl. S118), 49–54. [Google Scholar]
- Zhao, Q.; Bae, E.H.; Zhang, Y.; Shahsavari, A.; Lotey, P.; Lee, R.H.; Liu, F. Inhibitory Effects of Extracellular Vesicles from iPS-Cell-Derived Mesenchymal Stem Cells on the Onset of Sialadenitis in Sjogren’s Syndrome Are Mediated by Immunomodulatory Splenocytes and Improved by Inhibiting miR-125b. Int. J. Mol. Sci. 2023, 24, 5258. [Google Scholar] [CrossRef]
- Altman, J.; Jones, G.; Ahmed, S.; Sharma, S.; Sharma, A. Tear Film MicroRNAs as Potential Biomarkers: A Review. Int. J. Mol. Sci. 2023, 24, 3694. [Google Scholar] [CrossRef]
- Ogata, K.; Moriyama, M.; Kawado, T.; Yoshioka, H.; Yano, A.; Matsumura-Kawashima, M.; Nakamura, S.; Kawano, S. Extracellular vesicles of iPS cells highly capable of producing HGF and TGF-beta1 can attenuate Sjogren’s syndrome via innate immunity regulation. Cell Signal 2024, 113, 110980. [Google Scholar] [CrossRef]
- Kuret, T.; Lakota, K.; Cucnik, S.; Jurcic, V.; Distler, O.; Rotar, Z.; Hocevar, A.; Sodin-Semrl, S.; Frank-Bertoncelj, M. Dysregulated Expression of Arterial MicroRNAs and Their Target Gene Networks in Temporal Arteries of Treatment-Naive Patients with Giant Cell Arteritis. Int. J. Mol. Sci. 2021, 22, 6520. [Google Scholar] [CrossRef]
- Weng, K.P.; Cheng, C.F.; Chien, K.J.; Ger, L.P.; Huang, S.H.; Tsai, K.W. Identifying Circulating MicroRNA in Kawasaki Disease by Next-Generation Sequencing Approach. Curr. Issues Mol. Biol. 2021, 43, 485–500. [Google Scholar] [CrossRef]
- Cebi, A.H.; Demir, F.; Ikbal, M.; Kalyoncu, M. Plasma microRNA levels in childhood IgA vasculitis. Clin. Rheumatol. 2021, 40, 1975–1981. [Google Scholar] [CrossRef]
- Luo, Y.; Yu, M.; Li, P.; Huang, L.; Wu, J.; Kong, M.; Li, Y.; Wu, Z.; Kang, Z.; Yi, L.; et al. The Expression and Role of microRNA-133a in Plasma of Patients with Kawasaki Disease. Immunol. Investig. 2022, 51, 826–838. [Google Scholar] [CrossRef] [PubMed]
- Frydlova, J.; Zednikova, I.; Satrapova, V.; Pazourkova, E.; Santorova, S.; Hruskova, Z.; Tesar, V.; Vokurka, M.; Prikryl, P.; Korabecna, M. Analysis of microRNAs in Small Urinary Extracellular Vesicles and Their Potential Roles in Pathogenesis of Renal ANCA-Associated Vasculitis. Int. J. Mol. Sci. 2022, 23, 4344. [Google Scholar] [CrossRef] [PubMed]
- Surmiak, M.; Wawrzycka-Adamczyk, K.; Kosalka-Wegiel, J.; Polanski, S.; Sanak, M. Profile of circulating extracellular vesicles microRNA correlates with the disease activity in granulomatosis with polyangiitis. Clin. Exp. Immunol. 2022, 208, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Hocevar, A.; Tomsic, M.; Pizem, J.; Bolha, L.; Sodin-Semrl, S.; Glavac, D. MicroRNA expression in the affected skin of adult patients with IgA vasculitis. Clin. Rheumatol. 2019, 38, 339–345. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, L.; Chu, L.; Lan, J.; Wei, J.; Li, W.; Xue, C. Microscopic polyangiitis plasma-derived exosomal miR-1287-5p induces endothelial inflammatory injury and neutrophil adhesion by targeting CBL. PeerJ 2023, 11, e14579. [Google Scholar] [CrossRef] [PubMed]
- De la Rosa, I.A.; Perez-Sanchez, C.; Ruiz-Limon, P.; Patino-Trives, A.; Torres-Granados, C.; Jimenez-Gomez, Y.; Del Carmen Abalos-Aguilera, M.; Cecchi, I.; Ortega, R.; Caracuel, M.A.; et al. Impaired microRNA processing in neutrophils from rheumatoid arthritis patients confers their pathogenic profile. Modulation by biological therapies. Haematologica 2020, 105, 2250–2261. [Google Scholar] [CrossRef]
- Chatzikyriakidou, A.; Voulgari, P.V.; Georgiou, I.; Drosos, A.A. A polymorphism in the 3′-UTR of interleukin-1 receptor-associated kinase (IRAK1), a target gene of miR-146a, is associated with rheumatoid arthritis susceptibility. Jt. Bone Spine 2010, 77, 411–413. [Google Scholar] [CrossRef]
- Fasano, S.; Milone, A.; Nicoletti, G.F.; Isenberg, D.A.; Ciccia, F. Precision medicine in systemic lupus erythematosus. Nat. Rev. Rheumatol. 2023, 19, 331–342. [Google Scholar] [CrossRef]
- Hou, G.; Harley, I.T.W.; Lu, X.; Zhou, T.; Xu, N.; Yao, C.; Qin, Y.; Ouyang, Y.; Ma, J.; Zhu, X.; et al. SLE non-coding genetic risk variant determines the epigenetic dysfunction of an immune cell specific enhancer that controls disease-critical microRNA expression. Nat. Commun. 2021, 12, 135. [Google Scholar] [CrossRef]
- Yang, J.; Hamade, M.; Wu, Q.; Wang, Q.; Axtell, R.; Giri, S.; Mao-Draayer, Y. Current and Future Biomarkers in Multiple Sclerosis. Int. J. Mol. Sci. 2022, 23, 5877. [Google Scholar] [CrossRef] [PubMed]
- De Vito, F.; Balletta, S.; Caioli, S.; Musella, A.; Guadalupi, L.; Vanni, V.; Fresegna, D.; Bassi, M.S.; Gilio, L.; Sanna, K.; et al. MiR-142-3p is a Critical Modulator of TNF-mediated Neuronal Toxicity in Multiple Sclerosis. Curr. Neuropharmacol. 2023, 21, 2567–2582. [Google Scholar] [CrossRef] [PubMed]
- Mandolesi, G.; De Vito, F.; Musella, A.; Gentile, A.; Bullitta, S.; Fresegna, D.; Sepman, H.; Di Sanza, C.; Haji, N.; Mori, F.; et al. miR-142-3p Is a Key Regulator of IL-1beta-Dependent Synaptopathy in Neuroinflammation. J. Neurosci. 2017, 37, 546–561. [Google Scholar] [CrossRef] [PubMed]
- Ahlbrecht, J.; Martino, F.; Pul, R.; Skripuletz, T.; Suhs, K.W.; Schauerte, C.; Yildiz, O.; Trebst, C.; Tasto, L.; Thum, S.; et al. Deregulation of microRNA-181c in cerebrospinal fluid of patients with clinically isolated syndrome is associated with early conversion to relapsing-remitting multiple sclerosis. Mult. Scler. 2016, 22, 1202–1214. [Google Scholar] [CrossRef] [PubMed]
- Bergman, P.; Piket, E.; Khademi, M.; James, T.; Brundin, L.; Olsson, T.; Piehl, F.; Jagodic, M. Circulating miR-150 in CSF is a novel candidate biomarker for multiple sclerosis. Neurol. Neuroimmunol. Neuroinflamm 2016, 3, e219. [Google Scholar] [CrossRef] [PubMed]
- Zanghi, A.; Manuti, V.; Serviddio, G.; D’Amico, E.; Avolio, C. MiRNA 106a-5p in cerebrospinal fluid as signature of early relapsing remitting multiple sclerosis: A cross sectional study. Front. Immunol. 2023, 14, 1226130. [Google Scholar] [CrossRef] [PubMed]
- Casanova, I.; Dominguez-Mozo, M.I.; De Torres, L.; Aladro-Benito, Y.; Garcia-Martinez, A.; Gomez, P.; Abellan, S.; De Antonio, E.; Alvarez-Lafuente, R. MicroRNAs Associated with Disability Progression and Clinical Activity in Multiple Sclerosis Patients Treated with Glatiramer Acetate. Biomedicines 2023, 11, 2760. [Google Scholar] [CrossRef] [PubMed]
- Fattahi, M.; Rezaei, N.; Nematalahi, F.S.; Shaygannejad, V.; Fouladi, S.; Karimi, L.; Fathi, F.; Dehghani, L.; Mirmosayyeb, O.; Eskandari, N. MicroRNA-29b variants and MxA expression change during interferon beta therapy in patients with relapsing-remitting multiple sclerosis. Mult. Scler. Relat. Disord. 2019, 35, 241–245. [Google Scholar] [CrossRef]
- Mousavi, S.R.; Tahmasebivand, M.; Khorrami, M.; Ayromlou, H.; Khalili, S.K.; Khorvash, F.; Rikhtegar, R.; Khademi, B.; Bahmanpour, Z.; Emamalizadeh, B. Connection of miR-185 and miR-320a expression levels with response to interferon-beta in multiple sclerosis patients. Mult. Scler. Relat. Disord. 2020, 44, 102264. [Google Scholar] [CrossRef]
- Saghazadeh, A.; Rezaei, N. MicroRNA expression profiles of peripheral blood and mononuclear cells in myasthenia gravis: A systematic review. Int. Immunopharmacol. 2022, 112, 109205. [Google Scholar] [CrossRef]
- Jiang, L.; Cheng, Z.; Qiu, S.; Que, Z.; Bao, W.; Jiang, C.; Zou, F.; Liu, P.; Liu, J. Altered let-7 expression in Myasthenia gravis and let-7c mediated regulation of IL-10 by directly targeting IL-10 in Jurkat cells. Int. Immunopharmacol. 2012, 14, 217–223. [Google Scholar] [CrossRef]
- Mu, Y.; Huang, X.; Yang, Y.; Huang, Z.; Chen, J.; Li, S.; Dong, Y.; Chen, F.; Zhang, R.; Chen, J.; et al. Study of serum exosome miRNA as a biomarker for early onset adult ouclar myastthenia gravis. Gene 2023, 896, 148034. [Google Scholar] [CrossRef]
- Cavalcante, P.; Mizrachi, T.; Barzago, C.; Scandiffio, L.; Bortone, F.; Bonanno, S.; Frangiamore, R.; Mantegazza, R.; Bernasconi, P.; Brenner, T.; et al. MicroRNA signature associated with treatment response in myasthenia gravis: A further step towards precision medicine. Pharmacol. Res. 2019, 148, 104388. [Google Scholar] [CrossRef]
- Wang, J.; Zheng, S.; Xin, N.; Dou, C.; Fu, L.; Zhang, X.; Chen, J.; Zhang, Y.; Geng, D.; Xiao, C.; et al. Identification of novel MicroRNA signatures linked to experimental autoimmune myasthenia gravis pathogenesis: Down-regulated miR-145 promotes pathogenetic Th17 cell response. J. Neuroimmune Pharmacol. 2013, 8, 1287–1302. [Google Scholar] [CrossRef]
- Saleh, N.A.; Rode, M.P.; Cisilotto, J.; Silva, A.H.; Prigol, A.N.; da Luz Efe, F.; Winter, E.; Filippin-Monteiro, F.B.; Creczynski-Pasa, T.B. MicroRNA-Mediated Antiproliferative Effects of M1 Macrophage-Derived Extracellular Vesicles on Melanoma Cells. Immunol. Investig. 2023, 1–20. [Google Scholar] [CrossRef]
- Pesce, S.; Greppi, M.; Ferretti, E.; Obino, V.; Carlomagno, S.; Rutigliani, M.; Thoren, F.B.; Sivori, S.; Castagnola, P.; Candiani, S.; et al. miRNAs in NK Cell-Based Immune Responses and Cancer Immunotherapy. Front. Cell Dev. Biol. 2020, 8, 119. [Google Scholar] [CrossRef]
- Wells, A.C.; Hioki, K.A.; Angelou, C.C.; Lynch, A.C.; Liang, X.; Ryan, D.J.; Thesmar, I.; Zhanybekova, S.; Zuklys, S.; Ullom, J.; et al. Let-7 enhances murine anti-tumor CD8 T cell responses by promoting memory and antagonizing terminal differentiation. Nat. Commun. 2023, 14, 5585. [Google Scholar] [CrossRef]
- Otmani, K.; Rouas, R.; Lagneaux, L.; Krayem, M.; Duvillier, H.; Berehab, M.; Lewalle, P. Acute myeloid leukemia-derived exosomes deliver miR-24-3p to hinder the T-cell immune response through DENN/MADD targeting in the NF-kappaB signaling pathways. Cell Commun. Signal 2023, 21, 253. [Google Scholar] [CrossRef] [PubMed]
- Mundy-Bosse, B.L.; Scoville, S.D.; Chen, L.; McConnell, K.; Mao, H.C.; Ahmed, E.H.; Zorko, N.; Harvey, S.; Cole, J.; Zhang, X.; et al. MicroRNA-29b mediates altered innate immune development in acute leukemia. J. Clin. Investig. 2016, 126, 4404–4416. [Google Scholar] [CrossRef] [PubMed]
- Del Gaizo, M.; Sergio, I.; Lazzari, S.; Cialfi, S.; Pelullo, M.; Screpanti, I.; Felli, M.P. MicroRNAs as Modulators of the Immune Response in T-Cell Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2022, 23, 829. [Google Scholar] [CrossRef] [PubMed]
- Di Marco, M.; Veschi, S.; Lanuti, P.; Ramassone, A.; Pacillo, S.; Pagotto, S.; Pepe, F.; George-William, J.N.; Curcio, C.; Marchisio, M.; et al. Enhanced Expression of miR-181b in B Cells of CLL Improves the Anti-Tumor Cytotoxic T Cell Response. Cancers 2021, 13, 257. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Yan, F.; Wu, C. Overexpressed miR-195 attenuated immune escape of diffuse large B-cell lymphoma by targeting PD-L1. Biomed. Pharmacother. 2018, 98, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Estephan, R.J.; Wu, X.; Su, C.; Yuan, Y.C.; Qin, H.; Kil, S.H.; Morales, C.; Schmolze, D.; Sanchez, J.F.; et al. MicroRNA Regulation of T-Cell Exhaustion in Cutaneous T Cell Lymphoma. J. Investig. Dermatol. 2022, 142, 603–612.e7. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Coleborn, E.; Bhavsar, C.; Wang, Y.; Alim, L.; Wilkinson, A.N.; Tran, M.A.; Irgam, G.; Atluri, S.; Wong, K.; et al. miR-146a inhibits ovarian tumor growth in vivo via targeting immunosuppressive neutrophils and enhancing CD8+ T cell infiltration. Mol. Ther. Oncolytics 2023, 31, 100725. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Yu, H.; Zhang, Z.; Liu, Y.; Lou, G. Hypoxic tumour-derived exosomal miR-1225-5p regulates M2 macrophage polarisation via toll-like receptor 2 to promote ovarian cancer progress. Autoimmunity 2023, 56, 2281226. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Peng, B.; Yan, Y.; Liu, G.; Yang, D.; Wang, Q.; Li, Y.; Mao, Q.; Chen, Q. The tRF-3024b hijacks miR-192-5p to increase BCL-2-mediated resistance to cytotoxic T lymphocytes in Esophageal Squamous Cell Carcinoma. Int. Immunopharmacol. 2023, 126, 111135. [Google Scholar] [CrossRef]
- Ou, Z.; Lu, Y.; Xu, D.; Luo, Z. Hypoxia mediates immune escape of pancreatic cancer cells by affecting miR-1275/AXIN2 in natural killer cells. Front. Immunol. 2023, 14, 1271603. [Google Scholar] [CrossRef]
- Di Pace, A.L.; Pelosi, A.; Fiore, P.F.; Tumino, N.; Besi, F.; Quatrini, L.; Santopolo, S.; Vacca, P.; Moretta, L. MicroRNA analysis of Natural Killer cell-derived exosomes: The microRNA let-7b-5p is enriched in exosomes and participates in their anti-tumor effects against pancreatic cancer cells. Oncoimmunology 2023, 12, 2221081. [Google Scholar] [CrossRef]
- Liu, N.; Steer, C.J.; Song, G. MicroRNA-206 enhances antitumor immunity by disrupting the communication between malignant hepatocytes and regulatory T cells in c-Myc mice. Hepatology 2022, 76, 32–47. [Google Scholar] [CrossRef]
- Liu, W.; Liu, R.; Yuan, R.; Wang, X. MicroRNA-4458 Regulates PD-L1 Expression to Enhance Anti-tumor Immunity in NSCLC via Targeting STAT3. Mol. Biotechnol. 2021, 63, 1268–1279. [Google Scholar] [CrossRef]
- Li, G.; Chen, W.; Jiang, K.; Huang, J.; Zhong, J.; Liu, X.; Wei, T.; Gong, R.; Li, Z.; Zhu, J.; et al. Exosome-mediated delivery of miR-519e-5p promotes malignant tumor phenotype and CD8+ T-cell exhaustion in metastatic PTC. J. Clin. Endocrinol. Metab. 2023, dgad725. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Shi, Z.; Hao, Z.; Zhou, J.; Han, C.; Li, R.; Lv, Q.; Liu, Y.; Liang, C. Hypoxia-mediated down-regulation of miRNAs’ biogenesis promotes tumor immune escape in bladder cancer. Clin. Transl. Oncol. 2021, 23, 1678–1687. [Google Scholar] [CrossRef]
- Liang, X.; Liu, Y.; Mei, S.; Zhang, M.; Xin, J.; Zhang, Y.; Yang, R. MicroRNA-22 impairs anti-tumor ability of dendritic cells by targeting p38. PLoS ONE 2015, 10, e0121510. [Google Scholar] [CrossRef]
- Sarma, N.J.; Tiriveedhi, V.; Ramachandran, S.; Crippin, J.; Chapman, W.; Mohanakumar, T. Modulation of immune responses following solid organ transplantation by microRNA. Exp. Mol. Pathol. 2012, 93, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.; Suthanthiran, M.; Muthukumar, T. MicroRNAs and Transplantation. Clin. Lab. Med. 2019, 39, 125–143. [Google Scholar] [CrossRef]
- Chancharoenthana, W.; Traitanon, O.; Leelahavanichkul, A.; Tasanarong, A. Molecular immune monitoring in kidney transplant rejection: A state-of-the-art review. Front. Immunol. 2023, 14, 1206929. [Google Scholar] [CrossRef] [PubMed]
- Bezeljak, N.; Kojc, N.; Veceric Haler, Z.; Arnol, M.; Bostjancic, E. Tissue miRNA profile is associated with acute tubular necrosis, rejection phenotypes and BK polyomavirus-associated nephropathy in human kidney allografts. Nephron 2023. [Google Scholar] [CrossRef]
- Matz, M.; Fabritius, K.; Lorkowski, C.; Durr, M.; Gaedeke, J.; Durek, P.; Grun, J.R.; Goestemeyer, A.; Bachmann, F.; Wu, K.; et al. Identification of T Cell-Mediated Vascular Rejection After Kidney Transplantation by the Combined Measurement of 5 Specific MicroRNAs in Blood. Transplantation 2016, 100, 898–907. [Google Scholar] [CrossRef]
- Matz, M.; Lorkowski, C.; Fabritius, K.; Durek, P.; Wu, K.; Rudolph, B.; Neumayer, H.H.; Mashreghi, M.F.; Budde, K. Free microRNA levels in plasma distinguish T-cell mediated rejection from stable graft function after kidney transplantation. Transpl. Immunol. 2016, 39, 52–59. [Google Scholar] [CrossRef]
- Seo, J.W.; Lee, Y.H.; Tae, D.H.; Kim, Y.G.; Moon, J.Y.; Jung, S.W.; Kim, J.S.; Hwang, H.S.; Jeong, K.H.; Jeong, H.Y.; et al. Development and validation of urinary exosomal microRNA biomarkers for the diagnosis of acute rejection in kidney transplant recipients. Front. Immunol. 2023, 14, 1190576. [Google Scholar] [CrossRef]
- Millan, O.; Ruiz, P.; Orts, L.; Ferre, P.; Crespo, G.; Santana, M.; Fortuna, V.; Quintairos, L.; Navasa, M.; Brunet, M. Monitoring of miR-181a-5p and miR-155-5p Plasmatic Expression as Prognostic Biomarkers for Acute and Subclinical Rejection in de novo Adult Liver Transplant Recipients. Front. Immunol. 2019, 10, 873. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wang, Z.; Tan, C.J.; Liao, B.Y.; Zhang, X.; Xu, M.; Dai, Z.; Qiu, S.J.; Huang, X.W.; Sun, J.; et al. Plasma microRNA, a potential biomarker for acute rejection after liver transplantation. Transplantation 2013, 95, 991–999. [Google Scholar] [CrossRef] [PubMed]
- Millan, O.; Ruiz, P.; Julian, J.; Lizana, A.; Fundora, Y.; Crespo, G.; Colmenero, J.; Navasa, M.; Brunet, M. A plasmatic score using a miRNA signature and CXCL-10 for accurate prediction and diagnosis of liver allograft rejection. Front. Immunol. 2023, 14, 1196882. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; He, J.; Teng, X.; Yu, Y.; Ye, W.; Hu, Y.; Shen, Z. Combined intrathymic and intravenous injection of mesenchymal stem cells can prolong the survival of rat cardiac allograft associated with decrease in miR-155 expression. J. Surg. Res. 2013, 185, 896–903. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Su, W.; Chen, X.; Zhao, Q.; Zhang, N.; Li, M.G.; Yang, P.C.; Wang, L. Micro RNA-98 suppresses interleukin-10 in peripheral B cells in patient post-cardio transplantation. Oncotarget 2017, 8, 28237–28246. [Google Scholar] [CrossRef] [PubMed]
- Perez-Carrillo, L.; Sanchez-Lazaro, I.; Trivino, J.C.; Feijoo-Bandin, S.; Lago, F.; Gonzalez-Juanatey, J.R.; Martinez-Dolz, L.; Portoles, M.; Tarazon, E.; Rosello-Lleti, E. Combining Serum miR-144-3p and miR-652-3p as Potential Biomarkers for the Early Diagnosis and Stratification of Acute Cellular Rejection in Heart Transplantation Patients. Transplantation 2023, 107, 2064–2072. [Google Scholar] [CrossRef] [PubMed]
- Coutance, G.; Racape, M.; Baudry, G.; Lecuyer, L.; Roubille, F.; Blanchart, K.; Epailly, E.; Vermes, E.; Pattier, S.; Boignard, A.; et al. Validation of the clinical utility of microRNA as noninvasive biomarkers of cardiac allograft rejection: A prospective longitudinal multicenter study. J. Heart Lung Transplant. 2023, 42, 1505–1509. [Google Scholar] [CrossRef]
- Sang, W.; Zhang, C.; Zhang, D.; Wang, Y.; Sun, C.; Niu, M.; Sun, X.; Zhou, C.; Zeng, L.; Pan, B.; et al. MicroRNA-181a, a potential diagnosis marker, alleviates acute graft versus host disease by regulating IFN-gamma production. Am. J. Hematol. 2015, 90, 998–1007. [Google Scholar] [CrossRef]
- Xiao, B.; Wang, Y.; Li, W.; Baker, M.; Guo, J.; Corbet, K.; Tsalik, E.L.; Li, Q.J.; Palmer, S.M.; Woods, C.W.; et al. Plasma microRNA signature as a noninvasive biomarker for acute graft-versus-host disease. Blood 2013, 122, 3365–3375. [Google Scholar] [CrossRef]
- Liu, W.; Zhou, N.; Liu, Y.; Zhang, W.; Li, X.; Wang, Y.; Zheng, R.; Zhang, Y. Mesenchymal stem cell exosome-derived miR-223 alleviates acute graft-versus-host disease via reducing the migration of donor T cells. Stem Cell Res. Ther. 2021, 12, 153. [Google Scholar] [CrossRef]
- Reikvam, H.; Vo, A.K.; Johansen, S.; Hemsing, A.L.; Solheim, M.H.; Mosevoll, K.A.; Tvedt, T.H.A.; Hatfield, K.J. MicroRNA serum profiles and chronic graft-versus-host disease. Blood Adv. 2022, 6, 5295–5306. [Google Scholar] [CrossRef] [PubMed]
- Pitea, M.; Canale, F.A.; Porto, G.; Verduci, C.; Utano, G.; Policastro, G.; Alati, C.; Santoro, L.; Imbalzano, L.; Martino, M. The Role of MicroRNA in Graft-versus-Host-Disease: A Review. Genes 2023, 14, 1796. [Google Scholar] [CrossRef]
- Hasiuk, M.; Dolz, M.; Marone, R.; Jeker, L.T. Leveraging microRNAs for cellular therapy. Immunol. Lett. 2023, 262, 27–35. [Google Scholar] [CrossRef]
- Rad, S.; Halpin, J.C.; Tawinwung, S.; Suppipat, K.; Hirankarn, N.; McLellan, A.D. MicroRNA-mediated metabolic reprogramming of chimeric antigen receptor T cells. Immunol. Cell Biol. 2022, 100, 424–439. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, J.; Zheng, G.; Wang, Q.; Li, X.; Feng, Y.; Shang, F.; He, S.; Jiang, Q.; Shi, B.; et al. Co-Expression of miR155 or LSD1 shRNA Increases the Anti-Tumor Functions of CD19 CAR-T Cells. Front. Immunol. 2021, 12, 811364. [Google Scholar] [CrossRef] [PubMed]
- Alahdal, M.; Elkord, E. Non-coding RNAs in cancer immunotherapy: Predictive biomarkers and targets. Clin. Transl. Med. 2023, 13, e1425. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.K.; Shah, R.R.; Cidlowski, J.A. Glucocorticoids modulate microRNA expression and processing during lymphocyte apoptosis. J. Biol. Chem. 2010, 285, 36698–36708. [Google Scholar] [CrossRef]
- Ratman, D.; Vanden Berghe, W.; Dejager, L.; Libert, C.; Tavernier, J.; Beck, I.M.; De Bosscher, K. How glucocorticoid receptors modulate the activity of other transcription factors: A scope beyond tethering. Mol. Cell Endocrinol. 2013, 380, 41–54. [Google Scholar] [CrossRef]
- Hong, Y.; Wu, J.; Zhao, J.; Wang, H.; Liu, Y.; Chen, T.; Kan, X.; Tao, Q.; Shen, X.; Yan, K.; et al. miR-29b and miR-29c are involved in Toll-like receptor control of glucocorticoid-induced apoptosis in human plasmacytoid dendritic cells. PLoS ONE 2013, 8, e69926. [Google Scholar] [CrossRef]
- Maunder, D.; Brown, P.M.; Barron-Millar, B.; Lendrem, D.W.; Naamane, N.; Macdonald, J.; Wang, X.N.; Isaacs, J.D.; Anderson, A.E.; Morgan, A.W.; et al. Micro-RNA content of circulating extracellular vesicles in early rheumatoid arthritis as biomarkers and mediators of methotrexate efficacy. Rheumatology 2023, kead569. [Google Scholar] [CrossRef]
- Iwamoto, N.; Furukawa, K.; Endo, Y.; Shimizu, T.; Sumiyoshi, R.; Umeda, M.; Koga, T.; Kawashiri, S.Y.; Igawa, T.; Ichinose, K.; et al. Methotrexate Alters the Expression of microRNA in Fibroblast-like Synovial Cells in Rheumatoid Arthritis. Int. J. Mol. Sci. 2021, 22, 11561. [Google Scholar] [CrossRef]
- Yang, J.; Yang, X.; Yang, J.; Li, M. Hydroxychloroquine Inhibits the Differentiation of Th17 Cells in Systemic Lupus Erythematosus. J. Rheumatol. 2018, 45, 818–826. [Google Scholar] [CrossRef] [PubMed]
- Zisman, D.; Safieh, M.; Simanovich, E.; Feld, J.; Kinarty, A.; Zisman, L.; Gazitt, T.; Haddad, A.; Elias, M.; Rosner, I.; et al. Tocilizumab (TCZ) Decreases Angiogenesis in Rheumatoid Arthritis through Its Regulatory Effect on miR-146a-5p and EMMPRIN/CD147. Front. Immunol. 2021, 12, 739592. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Jiang, H.; Yang, J.; Mao, J.; Wei, G.; Meng, X.; Zang, H. LncRNA MIR31HG is induced by tocilizumab and ameliorates rheumatoid arthritis fibroblast-like synoviocyte-mediated inflammation via miR-214-PTEN-AKT signaling pathway. Aging 2021, 13, 24071–24085. [Google Scholar] [CrossRef] [PubMed]
- Sode, J.; Krintel, S.B.; Carlsen, A.L.; Hetland, M.L.; Johansen, J.S.; Horslev-Petersen, K.; Stengaard-Pedersen, K.; Ellingsen, T.; Burton, M.; Junker, P.; et al. Plasma MicroRNA Profiles in Patients with Early Rheumatoid Arthritis Responding to Adalimumab plus Methotrexate vs Methotrexate Alone: A Placebo-controlled Clinical Trial. J. Rheumatol. 2018, 45, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Lu, J.; Jing, S.; Xi, J.; Yan, C.; Song, J.; Luo, S.; Zhao, C. Low-dose rituximab lowers serum Exosomal miR-150-5p in AChR-positive refractory myasthenia gravis patients. J. Neuroimmunol. 2020, 348, 577383. [Google Scholar] [CrossRef] [PubMed]
- Kotewitsch, M.; Heimer, M.; Schmitz, B.; Mooren, F.C. Non-coding RNAs in exercise immunology: A systematic review. J. Sport. Health Sci. 2023, in press. [CrossRef] [PubMed]
- Goldberg, Y.; Segal, S.; Hamdi, L.; Nabat, H.; Fainstein, N.; Mediouni, E.; Asis, Y.; Theotokis, P.; Salamotas, I.; Grigoriadis, N.; et al. High-intensity interval training attenuates development of autoimmune encephalomyelitis solely by systemic immunomodulation. Sci. Rep. 2023, 13, 16513. [Google Scholar] [CrossRef]
- Schmitz, B.; Niehues, H.; Lenders, M.; Thorwesten, L.; Klose, A.; Kruger, M.; Brand, E.; Brand, S.M. Effects of high-intensity interval training on microvascular glycocalyx and associated microRNAs. Am. J. Physiol. Heart Circ. Physiol. 2019, 316, H1538–H1551. [Google Scholar] [CrossRef]
- Castano, C.; Mirasierra, M.; Vallejo, M.; Novials, A.; Parrizas, M. Delivery of muscle-derived exosomal miRNAs induced by HIIT improves insulin sensitivity through down-regulation of hepatic FoxO1 in mice. Proc. Natl. Acad. Sci. USA 2020, 117, 30335–30343. [Google Scholar] [CrossRef]
- Gu, Y.; Zhou, H.; Yu, H.; Yang, W.; Wang, B.; Qian, F.; Cheng, Y.; He, S.; Zhao, X.; Zhu, L.; et al. miR-99a regulates CD4+ T cell differentiation and attenuates experimental autoimmune encephalomyelitis by mTOR-mediated glycolysis. Mol. Ther. Nucleic Acids 2021, 26, 1173–1185. [Google Scholar] [CrossRef]
- Sumikawa, M.H.; Iwata, S.; Zhang, M.; Miyata, H.; Ueno, M.; Todoroki, Y.; Nagayasu, A.; Kanda, R.; Sonomoto, K.; Torimoto, K.; et al. An enhanced mitochondrial function through glutamine metabolism in plasmablast differentiation in systemic lupus erythematosus. Rheumatology 2022, 61, 3049–3059. [Google Scholar] [CrossRef]
- Leone, R.D.; Zhao, L.; Englert, J.M.; Sun, I.M.; Oh, M.H.; Sun, I.H.; Arwood, M.L.; Bettencourt, I.A.; Patel, C.H.; Wen, J.; et al. Glutamine blockade induces divergent metabolic programs to overcome tumor immune evasion. Science 2019, 366, 1013–1021. [Google Scholar] [CrossRef]
- Fan, W.H.; Wang, F.C.; Jin, Z.; Zhu, L.; Zhang, J.X. Curcumin Synergizes with Cisplatin to Inhibit Colon Cancer through Targeting the MicroRNA-137-Glutaminase Axis. Curr. Med. Sci. 2022, 42, 108–117. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, J.; Li, Y.; Fan, J.; Chen, L.; Xu, R. MicroRNA-153 regulates glutamine metabolism in glioblastoma through targeting glutaminase. Tumour Biol. 2017, 39, 1010428317691429. [Google Scholar] [CrossRef]
- Lv, R.; Pan, X.; Song, L.; Sun, Q.; Guo, C.; Zou, S.; Zhou, Q. MicroRNA-200a-3p accelerates the progression of osteoporosis by targeting glutaminase to inhibit osteogenic differentiation of bone marrow mesenchymal stem cells. Biomed. Pharmacother. 2019, 116, 108960. [Google Scholar] [CrossRef]
- Sassi, F.; Tamone, C.; D’Amelio, P. Vitamin D: Nutrient, Hormone, and Immunomodulator. Nutrients 2018, 10, 1656. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Chen, Y.; Liu, W.; Thadhani, R. MicroRNA-mediated mechanism of vitamin D regulation of innate immune response. J. Steroid Biochem. Mol. Biol. 2014, 144 Pt A, 81–86. [Google Scholar] [CrossRef]
- Mohammadi Kordkhayli, M.; Mansouri, F.; Talebi, F.; Noorbakhsh, F.; Saboor-Yaraghi, A.A. Influence of Vitamins A and D on the Expression of MicroRNA27-3p Isoforms and GATA3 in Experimental Autoimmune Encephalomyelitis. Iran. J. Allergy Asthma Immunol. 2022, 21, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Alehagen, U.; Johansson, P.; Aaseth, J.; Alexander, J.; Wagsater, D. Significant changes in circulating microRNA by dietary supplementation of selenium and coenzyme Q10 in healthy elderly males. A subgroup analysis of a prospective randomized double-blind placebo-controlled trial among elderly Swedish citizens. PLoS ONE 2017, 12, e0174880. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.Y.; Luo, Q.; Sui, H.; Du, X.N.; Zhao, Y.J.; Liu, L.; Guan, Q.; Zhou, Y.; Wen, Q.S.; Shi, Y.; et al. Recent advances in the involvement of epigenetics in the pathogenesis of systemic lupus erythematosus. Clin. Immunol. 2023, 258, 109857. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, A.D.; Allis, C.D.; Bernstein, E. Epigenetics: A landscape takes shape. Cell 2007, 128, 635–638. [Google Scholar] [CrossRef] [PubMed]
- Mi, S.; Zhang, J.; Zhang, W.; Huang, R.S. Circulating microRNAs as biomarkers for inflammatory diseases. Microrna 2013, 2, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Weidhaas, J.B. Identifying MicroRNA Pathway Variants as Biomarkers of Patient Selection for Immune Therapy. Methods Mol. Biol. 2020, 2055, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.D.; Wei, Z.; Silva, M.C.; Barberan-Soler, S.; Zhang, J.; Rabinovsky, R.; Muratore, C.R.; Stricker, J.M.S.; Hortman, C.; Young-Pearse, T.L.; et al. Small molecule regulators of microRNAs identified by high-throughput screen coupled with high-throughput sequencing. Nat. Commun. 2023, 14, 7575. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, M.L.; Zamecnik, P.C. Inhibition of Rous sarcoma viral RNA translation by a specific oligodeoxyribonucleotide. Proc. Natl. Acad. Sci. USA 1978, 75, 285–288. [Google Scholar] [CrossRef]
- Roehr, B. Fomivirsen approved for CMV retinitis. J. Int. Assoc. Physicians AIDS Care 1998, 4, 14–16. [Google Scholar]
- Moumne, L.; Marie, A.C.; Crouvezier, N. Oligonucleotide Therapeutics: From Discovery and Development to Patentability. Pharmaceutics 2022, 14, 260. [Google Scholar] [CrossRef]
- Ho, P.T.B.; Clark, I.M.; Le, L.T.T. MicroRNA-Based Diagnosis and Therapy. Int. J. Mol. Sci. 2022, 23, 7167. [Google Scholar] [CrossRef]
- Jeker, L.T.; Marone, R. Targeting microRNAs for immunomodulation. Curr. Opin. Pharmacol. 2015, 23, 25–31. [Google Scholar] [CrossRef]
- Jeker, L.T.; Bluestone, J.A. MicroRNA regulation of T-cell differentiation and function. Immunol. Rev. 2013, 253, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.J.; Bahal, R.; Babar, I.A.; Pincus, Z.; Barrera, F.; Liu, C.; Svoronos, A.; Braddock, D.T.; Glazer, P.M.; Engelman, D.M.; et al. MicroRNA silencing for cancer therapy targeted to the tumour microenvironment. Nature 2015, 518, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Berg, J.L.; Perfler, B.; Hatzl, S.; Mayer, M.C.; Wurm, S.; Uhl, B.; Reinisch, A.; Klymiuk, I.; Tierling, S.; Pregartner, G.; et al. Micro-RNA-125a mediates the effects of hypomethylating agents in chronic myelomonocytic leukemia. Clin. Epigenetics 2021, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, A.; Mohammadi, A.; Fallah, S. Epigenetic Modification of MicroRNA-219-1 and Its Association with Glioblastoma Multiforme. Biochemistry 2021, 86, 420–432. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.K.; Mattie, M.D.; Berger, C.E.; Benz, S.C.; Benz, C.C. Rapid alteration of microRNA levels by histone deacetylase inhibition. Cancer Res. 2006, 66, 1277–1281. [Google Scholar] [CrossRef] [PubMed]
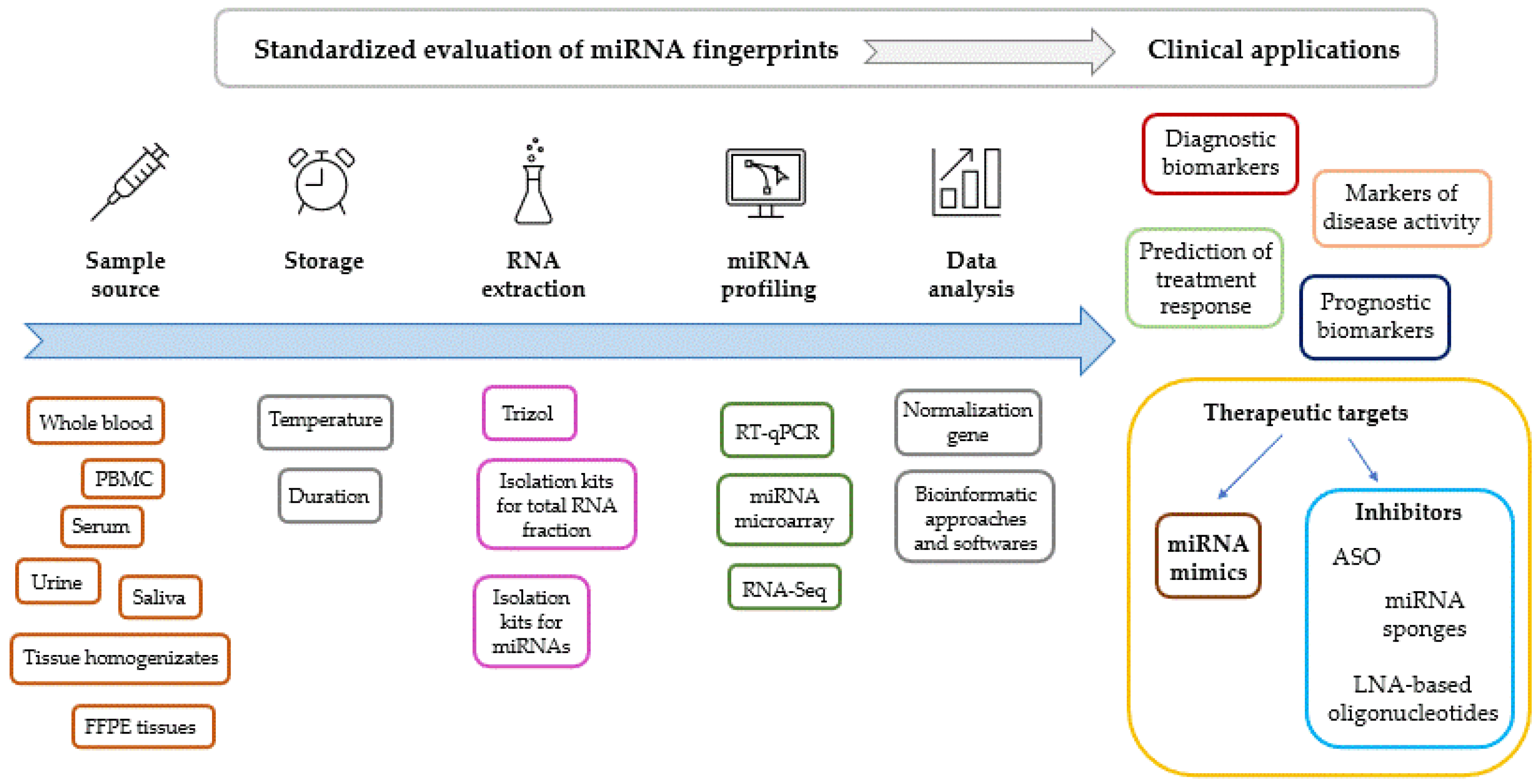
- Mraz, M.; Malinova, K.; Mayer, J.; Pospisilova, S. MicroRNA isolation and stability in stored RNA samples. Biochem. Biophys. Res. Commun. 2009, 390, 1–4. [Google Scholar] [CrossRef]
- Lekchnov, E.A.; Zaporozhchenko, I.A.; Morozkin, E.S.; Bryzgunova, O.E.; Vlassov, V.V.; Laktionov, P.P. Protocol for miRNA isolation from biofluids. Anal. Biochem. 2016, 499, 78–84. [Google Scholar] [CrossRef]
- Moret, I.; Sanchez-Izquierdo, D.; Iborra, M.; Tortosa, L.; Navarro-Puche, A.; Nos, P.; Cervera, J.; Beltran, B. Assessing an improved protocol for plasma microRNA extraction. PLoS ONE 2013, 8, e82753. [Google Scholar] [CrossRef]
- Ahlberg, E.; Jenmalm, M.C.; Tingo, L. Evaluation of five column-based isolation kits and their ability to extract miRNA from human milk. J. Cell Mol. Med. 2021, 25, 7973–7979. [Google Scholar] [CrossRef]
- McAlexander, M.A.; Phillips, M.J.; Witwer, K.W. Comparison of Methods for miRNA Extraction from Plasma and Quantitative Recovery of RNA from Cerebrospinal Fluid. Front. Genet. 2013, 4, 83. [Google Scholar] [CrossRef]
- Roest, H.P.; JNM, I.J.; van der Laan, L.J.W. Evaluation of RNA isolation methods for microRNA quantification in a range of clinical biofluids. BMC Biotechnol. 2021, 21, 48. [Google Scholar] [CrossRef]
- Tajbakhsh, A.; Bianconi, V.; Pirro, M.; Gheibi Hayat, S.M.; Johnston, T.P.; Sahebkar, A. Efferocytosis and Atherosclerosis: Regulation of Phagocyte Function by MicroRNAs. Trends Endocrinol. Metab. 2019, 30, 672–683. [Google Scholar] [CrossRef]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef]
- Leber, M.F.; Bossow, S.; Leonard, V.H.; Zaoui, K.; Grossardt, C.; Frenzke, M.; Miest, T.; Sawall, S.; Cattaneo, R.; von Kalle, C.; et al. MicroRNA-sensitive oncolytic measles viruses for cancer-specific vector tropism. Mol. Ther. 2011, 19, 1097–1106. [Google Scholar] [CrossRef] [PubMed]
- Bramsen, J.B.; Laursen, M.B.; Nielsen, A.F.; Hansen, T.B.; Bus, C.; Langkjaer, N.; Babu, B.R.; Hojland, T.; Abramov, M.; Van Aerschot, A.; et al. A large-scale chemical modification screen identifies design rules to generate siRNAs with high activity, high stability and low toxicity. Nucleic Acids Res. 2009, 37, 2867–2881. [Google Scholar] [CrossRef]
- Chiou, G.Y.; Cherng, J.Y.; Hsu, H.S.; Wang, M.L.; Tsai, C.M.; Lu, K.H.; Chien, Y.; Hung, S.C.; Chen, Y.W.; Wong, C.I.; et al. Cationic polyurethanes-short branch PEI-mediated delivery of Mir145 inhibited epithelial-mesenchymal transdifferentiation and cancer stem-like properties and in lung adenocarcinoma. J. Control Release 2012, 159, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Kasina, V.; Wahane, A.; Liu, C.H.; Yang, L.; Nieh, M.P.; Slack, F.J.; Bahal, R. Next-generation poly-L-histidine formulations for miRNA mimic delivery. Mol. Ther. Methods Clin. Dev. 2023, 29, 271–283. [Google Scholar] [CrossRef]
- Winkler, J. Nanomedicines based on recombinant fusion proteins for targeting therapeutic siRNA oligonucleotides. Ther. Deliv. 2011, 2, 891–905. [Google Scholar] [CrossRef]
- Baumann, V.; Winkler, J. miRNA-based therapies: Strategies and delivery platforms for oligonucleotide and non-oligonucleotide agents. Future Med. Chem. 2014, 6, 1967–1984. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, I.; Chatterjee, A. Recent Advances in miRNA Delivery Systems. Methods Protoc. 2021, 4, 10. [Google Scholar] [CrossRef]
- Baum, C.; Dullmann, J.; Li, Z.; Fehse, B.; Meyer, J.; Williams, D.A.; von Kalle, C. Side effects of retroviral gene transfer into hematopoietic stem cells. Blood 2003, 101, 2099–2114. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Sun, Y.; Zhao, R. Establishment of MicroRNA delivery system by PP7 bacteriophage-like particles carrying cell-penetrating peptide. J. Biosci. Bioeng. 2017, 124, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Daya, S.; Berns, K.I. Gene therapy using adeno-associated virus vectors. Clin. Microbiol. Rev. 2008, 21, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Z.; Gemeinhart, R.A. Progress in microRNA delivery. J. Control Release 2013, 172, 962–974. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Microorganism | miRNA | Target/Mechanism of Action |
|---|---|---|
| Virus | ||
| HBV | miR-1231 | HBV core mRNA [57] |
| HCV | miR-122 | viral translation [58] |
| Influenza A (H1N1) | miR-21, miR-99a | pulmonary repair and inflammatory processes [59] |
| HMPV | miR-4634 | IFN production [60] |
| HSV1 | miR-24 | STING [61] |
| EBV | miR-BART17-3p | RIG-I like receptor antiviral pathway [62] |
| ZIKV | miR-30c, miR-194, miR-302b, miR-302c | microcephaly and brain lesions [63] |
| HIV-1 | miR-155, miR-181-5p | TLR, DDX3X [64] |
| SARS-CoV2 | let-7, miR-17-5p, miR-18a-5p, miR-19a-3p, miR-20a-5p, miR-124, miR-155, miR-200c-3p, miR-223 | production of inflammatory cytokines [65,66], development of fibrotic lung lesions [67] |
| Bacteria | ||
| M. tuberculosis | miR-155, miR-215-5p | fusion of autophagosomes with lysosomes [68,69] |
| M. avium subsp. paratuberculosis | miR-150 | PDCD4 [70] |
| S. pneumoniae | miR-223 | pulmonary inflammation [71] |
| N. meningitidis | miR-146a | induction of endotoxin tolerance [72] |
| L. pneumophila | miR-125a-5p | NTAN1 [73] |
| K. pneumoniae | miR-181a-5p | alleviation of lung injury [74] |
| B. abortus | miR-125b-5p | TNFα production [75] |
| Fungi | ||
| C. albicans | miR-16-1, miR-17-3p, miR-155 | SOCS1 [76,77] |
| C. glabrata | miR-146a | production of inflammatory cytokines [78] |
| A. fumigatus | miR-21-5p, miR-26-5p, miR-142-3p, miR-142-5p | markers of invasive aspergillosis [79] |
| Parasite | ||
| B. malayi | miR-34 | CXCL10/CXCL11/CXCR3 [80] |
| T. cruzi | miR-155 | TNFα and IFN production, NK cell function [81] |
| L. donovani | miR-21, miR-146b-5p | increased expression of IL-6 and STAT3 [82] |
| S. japonicum | miR-146b, miR-155 | hepatic inflammation [83] |
| P. falciparum | miR-27a, miR-150, let-7i | markers of cerebral malaria [84] |
| Disease | Pathogenesis | Diagnosis/Disease Activity | Prognosis/Treatment Response | Therapy |
|---|---|---|---|---|
| Rheumatoid arthritis | miR-146a (rs2431697) [105], miR-200a-3p [106] | miR-21-5p [107], miR-25, miR-371b, miR-378d, miR-483, miR-642b [108] | miR-19b [109], miR-23, miR-223 [110] | miR-155 [111], miR-340-5p [112] |
| Systemic lupus erythematosus | miR-17-92 (rs4284505) [113], miR-146a [114], miR-365a-3p [115], miR-4689 [116] | miR-21 [117], miR-124-3p, miR-146, miR-320b, miR-377-3p [118,119] | miR-181a, miR-223 [120], miR-485-5p [121] | miR-7 [122], miR-30a [123], miR-181d-5p [124], miR-590-3p [125] |
| Antiphospholipid syndrome | miR-326, miR-483-3p [126] | miR-19b-3p, miR-20a-5p [127] | miR-146a-5p [128] | |
| Systemic sclerosis, scleroderma | miR-21 [129], miR-126 [130], miR-155-5p [131] | miR-21 [132], miR-27a [133], miR-29a, miR-138 [134] | miR-21-5p, miR-29a-3p, miR-143, miR-155 [135], miR-181b-5p, miR-210-3p, miR-223-3p [136] | miR-27a [137], miR-214 [138] |
| Sjögren’s syndrome | miR-181d-5p [139], miR-223-3p [140] | miR-30 [141], miR-181b-5p, miR-322-5p, miR-424-5p, miR-503-5p [142], miR-513c-3p [143], miR-1290, let-7b-5p [144] | miR-92a [145] | miR-16, miR-125b [146], miR-142, miR-223 [147], let-7 [148] |
| Autoimmune vasculitis | miR-424-5p [149] | miR-24-3p [150], miR-30a-5p, miR-33, miR-34 [151], miR-99a-5p, miR-106b-5p, miR-133a [152], miR-146a-5p [149], miR-148b-3p, miR-155-5p, miR-182-5p [153], miR-223-3p, miR-664-3p [154], let-7b [155] | miR-1287-5p [156] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaál, Z. Role of microRNAs in Immune Regulation with Translational and Clinical Applications. Int. J. Mol. Sci. 2024, 25, 1942. https://doi.org/10.3390/ijms25031942
Gaál Z. Role of microRNAs in Immune Regulation with Translational and Clinical Applications. International Journal of Molecular Sciences. 2024; 25(3):1942. https://doi.org/10.3390/ijms25031942
Chicago/Turabian StyleGaál, Zsuzsanna. 2024. "Role of microRNAs in Immune Regulation with Translational and Clinical Applications" International Journal of Molecular Sciences 25, no. 3: 1942. https://doi.org/10.3390/ijms25031942