
Ultra-Fast Vitrification: Minimizing the Toxicity of Cryoprotective Agents and Osmotic Stress in Mouse Oocyte Cryopreservation

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Survival Rates of Oocytes
2.2. Distribution and Fluorescence Intensity of ER
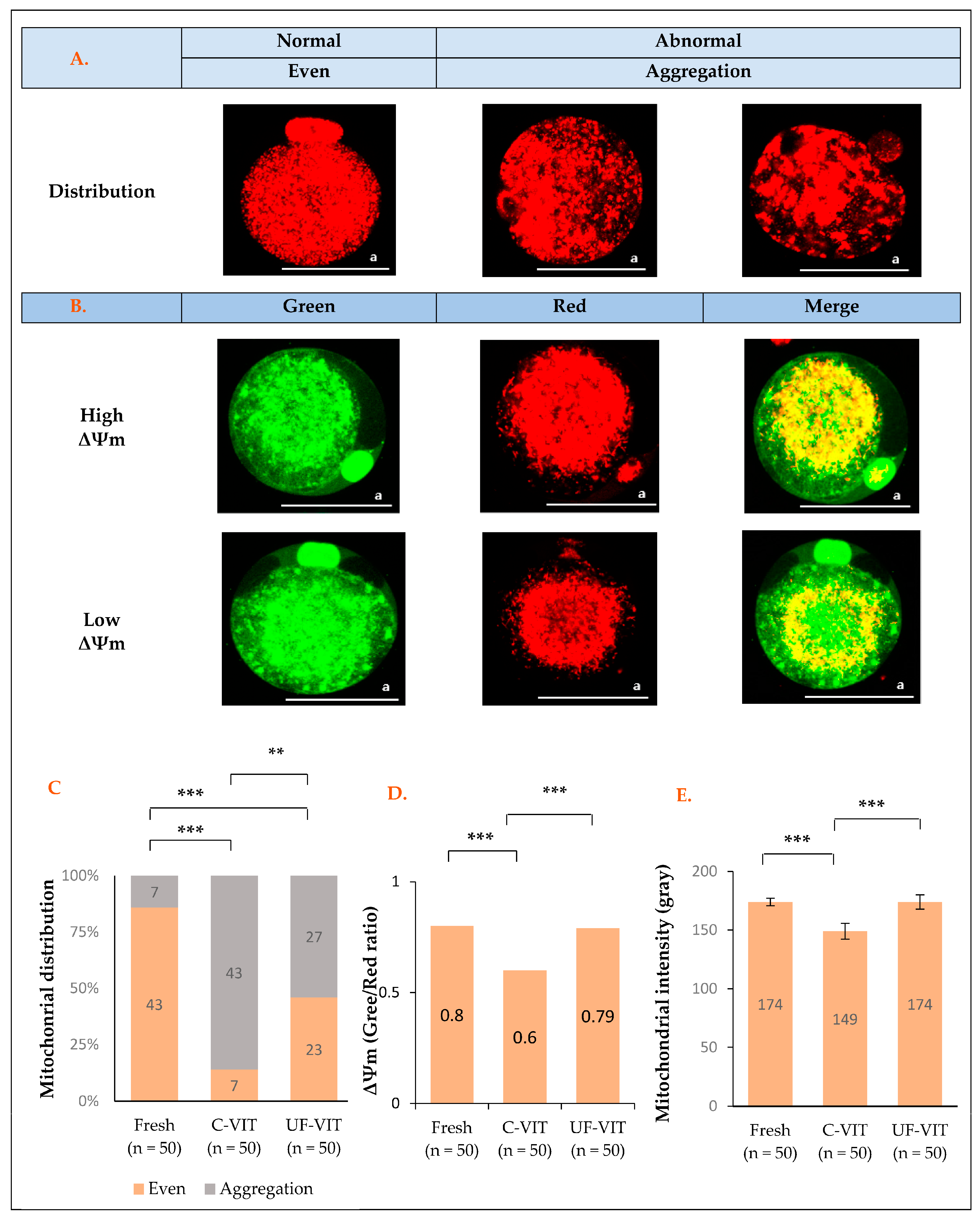
2.3. Distribution and Fluorescence Intensity of MT and Mitochondrial Membrane Potential Analysis
2.4. Meiotic Spindle in and Chromosomal Morphology of Oocytes
2.5. Comparison of Oocyte Volume Excursion Based on CPA Exposure Time
2.6. Comparing Embryonic Development Rates
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Oocyte Collection
4.3. Vitrification and Warming Protocols for C-VIT and UF-VIT
4.4. Survival Rates of Oocytes
4.5. Live Imaging of ER and MT in Mouse Oocytes Using a Confocal Microscope
4.6. Measurement of MT Membrane Potential Using JC-1 Staining
4.7. Immunofluorescence Staining of Oocytes
4.8. Image Acquisition and Relative Volume Excursion of Oocytes
4.9. Intracytoplasmic Sperm Injection
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ART | Assisted Reproductive Technology. |
| Ca2+ | Calcium ions. |
| [Ca2+]i | Intracellular calcium. |
| C-VIT | Conventional Vitrification. |
| CO2 | Carbon Dioxide. |
| CPA | Cryoprotective Agents. |
| DAPI | 6-Diamidino-2Phenylindole. |
| ΔΨm | Mitochondrial Membrane Potential. |
| DPBS | Dulbecco’s Phosphate-Buffered Saline. |
| DMSO | Dimethyl Sulfoxide. |
| EG | Ethylene Glycol. |
| ER | Endoplasmic Reticulum. |
| ES | Equilibration Solution. |
| hCG | Human Chorionic Gonadotropin. |
| ICSI | Intracytoplasmic Sperm Injection. |
| IP3 | Inositol 1,4,5-Trisphosphate. |
| IP3R1 | Inositol 1,4,5-Trisphosphate Receptor. |
| LSCM | Laser-Scanning Confocal Microscope. |
| MT | Mitochondrial. |
| MII | Metaphase II. |
| OC | Oocyte Cryopreservation. |
| PBS | Phosphate-Buffered Saline. |
| PVA | Polyvinyl Alcohol. |
| RT | Room Temperature. |
| SSS | Serum Substitute Supplement. |
| UF-VIT | Ultra-Fast Vitrification. |
| VS | Vitrification Solution. |
| VW | Vitrification/Warming. |
References
- Rienzi, L.; Martinez, F.; Ubaldi, F.; Minasi, M.G.; Iacobelli, M.; Tesarik, J.; Greco, E. Polscope analysis of meiotic spindle changes in living metaphase II human oocytes during the freezing and thawing procedures. Hum. Reprod. 2004, 19, 655–659. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Guo, N.; Tan, M.H.; Li, Y.F. Effect of mouse oocyte vitrification on mitochondrial membrane potential and distribution. J. Huazhong. Univ. Sci. Technol. 2014, 34, 4. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.U.; Lien, Y.R.; Chao, K.H.; Ho, H.N.; Yang, Y.S.; Lee, T.Y. Effects of cryopreservation on meiotic spindles of oocytes and its dynamics after thawing: Clinical implications in oocyte freezing—A review article. Mol. Cell. Endocrinol. 2003, 202, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Varlas, V.N.; Bors, R.G.; Albu, D.; Penes, O.N.; Nasui, B.A.; Mehedintu, C.; Pop, A.L. Social freezing: Pressing pause on fertility. Int. J. Environ. Res. Public. Health 2021, 18, 8088. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, Z.; Chen, Y.; He, F. A modified cryoloop vitrification protocol in the cryopreservation of mature mouse oocytes. Zygote 2009, 17, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Vanderzwalmen, P.; Ectors, F.; Panagiotidis, Y.; Schuff, M.; Murtinger, M.; Wirleitner, B. The evolution of the cryopreservation techniques in reproductive medicine—Exploring the character of the vitrified state intra- and extracellularly to better understand cell survival after cryopreservation. Reprod. Med. 2020, 1, 142–157. [Google Scholar] [CrossRef]
- Camargos, M.; Rodrigues, J.K.; Lobach, V.N.; El Cury-Silva, T.; Nunes, M.E.G.; Camargos, A.F.; Reis, F.M. Human oocyte morphometry before and after cryopreservation: A prospective cohort study. Cryobiology 2019, 88, 81–86. [Google Scholar] [CrossRef]
- Gala, A.; Ferrières-Hoa, A.; Loup-Cabaniols, V.; Fournier, A.; Anav, M.; Brunet, C.; Bringer-Deutsch, S.; Ranisavljevic, N.; Brouillet, S.; Hamamah, S. Closed vitrification system and egg donation: Predictive factors of oocyte survival and pregnancy. J. Gynecol. Obstet. Hum. Reprod. 2020, 49, 101687. [Google Scholar] [CrossRef]
- Lowther, K.M.; Weitzman, V.N.; Maier, D.; Mehlmann, L.M. Maturation, fertilization, and the structure and function of the endoplasmic reticulum in cryopreserved mouse oocytes. Biol. Reprod. 2009, 81, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Konc, J.; Kanyo, K.; Kriston, R.; Somoskoi, B.; Cseh, S. Cryopreservation of embryos and oocytes in human assisted reproduction. Biomed. Res. Int. 2014, 2014, 307268. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.D.; Motta, E.E.; Serafini, P. Theoretical and experimental basis of oocyte vitrification. Reprod. Biomed. Online 2011, 23, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.Y.; Yoon, S.Y.; Cha, S.K.; Kwak, K.H.; Fissore, R.A.; Parys, J.B.; Yoon, T.K.; Lee, D.R. Alterations in calcium oscillatory activity in vitrified mouse eggs impact on egg quality and subsequent embryonic development. Pflugers. Arch. 2011, 461, 515–526. [Google Scholar] [CrossRef]
- Tamura, A.N.; Huang, T.T.F.; Marikawa, Y. Impact of vitrification on the meiotic spindle and components of the microtubule -mouse. Biol. Reprod. 2013, 89, 112. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Han, S.; Liu, W.; Wang, Y.; Huang, G. Effect of vitrification on mitochondrial membrane potential in human metaphase II oocytes. J. Assist. Reprod. Genet. 2012, 29, 1045–1050. [Google Scholar] [CrossRef]
- Amidi, F.; Khodabandeh, Z.; Nori Mogahi, M.H. Comparison of the effects of vitrification on gene expression of mature mouse oocytes using Cryotop and open pulled straw. Int. J. Fertil. Steril. 2018, 12, 61–67. [Google Scholar] [PubMed]
- Lee, J.H.; Park, J.K.; Yoon, S.Y.; Park, E.A.; Jun, J.H.; Lim, H.J.; Kim, J.; Song, H. Advanced maternal age deteriorates the developmental competence of vitrified oocytes in mice. Cells 2021, 10, 1563. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Hao, H.S.; Li, C.Y.; Zhao, Y.H.; Wang, H.Y.; Yan, C.L.; Du, W.H.; Wang, D.; Liu, Y.; Pang, Y.W.; et al. Calcium ion regulation by BAPTA-AM and ruthenium red improved the fertilisation capacity and developmental ability of vitrified bovine oocytes. Sci. Rep. 2017, 7, 10652. [Google Scholar] [CrossRef] [PubMed]
- Jegal, H.-G.; Park, H.-J.; Kim, J.-W.; Koo, D.-B. Confirmation of mitochondrial specific Rhod-2 expression patterns in matured and fertilized porcine oocytes in vitro. Ann. Anim. Resour. Sci. 2019, 30, 8–17. [Google Scholar] [CrossRef]
- Stigliani, S.; Moretti, S.; Casciano, I.; Canepa, P.; Remorgida, V.; Anserini, P.; Scaruffi, P. Presence of aggregates of smooth endoplasmic reticulum in MII oocytes affects oocyte competence: Molecular-based evidence. Mol. Hum. Reprod. 2018, 24, 310–317. [Google Scholar] [CrossRef]
- Jin, B.; Mazur, P. High survival of mouse oocytes/embryos after vitrification without permeating cryoprotectants followed by ultra-rapid warming with an IR laser pulse. Sci. Rep. 2015, 5, 9271. [Google Scholar] [CrossRef]
- Gallardo, M.; Saenz, J.; Risco, R. Human oocytes and zygotes are ready for ultra-fast vitrification after 2 minutes of exposure to standard CPA solutions. Sci. Rep. 2019, 9, 15986. [Google Scholar] [CrossRef]
- Wang, L.; Liu, J.; Zhou, G.B.; Hou, Y.P.; Li, J.J.; Zhu, S.E. Quantitative investigations on the effects of exposure durations to the combined cryoprotective agents on mouse oocyte vitrification procedures. Biol. Reprod. 2011, 85, 884–894. [Google Scholar] [CrossRef]
- Zhang, C.H.; Qian, W.P.; Qi, S.T.; Ge, Z.J.; Min, L.J.; Zhu, X.L.; Huang, X.; Liu, J.P.; Ouyang, Y.C.; Hou, Y.; et al. Maternal diabetes causes abnormal dynamic changes of endoplasmic reticulum during mouse oocyte maturation and early embryo development. Reprod. Biol. Endocrinol. 2013, 11, 31. [Google Scholar] [CrossRef]
- Wang, Q.; Stringer, J.M.; Liu, J.; Hutt, K.J. Evaluation of mitochondria in oocytes following gamma-irradiation. Sci. Rep. 2019, 9, 19941. [Google Scholar] [CrossRef] [PubMed]
- Roth, Z. Symposium review: Reduction in oocyte developmental competence by stress is associated with alterations in mitochondrial function. J. Dairy. Sci. 2018, 101, 3642–3654. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.Y.; Xiang, D.C.; Zhang, B.; Quan, G.B.; Shao, Q.Y.; Hong, Q.H.; Wu, G.Q. Quality of vitrified porcine immature oocytes is improved by coculture with fresh oocytes during in vitro maturation. Mol. Reprod. Dev. 2019, 86, 1615–1627. [Google Scholar] [CrossRef]
- Schatten, H.; Sun, Q.Y.; Prather, R. The impact of mitochondrial function/dysfunction on IVF and new treatment possibilities for infertility. Reprod. Biol. Endocrinol. 2014, 12, 111. [Google Scholar] [CrossRef]
- Wilding, M.; Dale, B.; Marino, M.; di Matteo, L.; Alviggi, C.; Pisaturo, M.L.; Lombardi, L.; Placido, G.D. Mitochondrial aggregation patterns and activity in human oocytes and preimplantation embryos. Hum. Reprod. 2001, 16, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Hou, H.Y.; Wang, X.; Yu, Q.; Li, H.Y.; Li, S.J.; Tang, R.Y.; Guo, Z.X.; Chen, Y.Q.; Hu, C.X.; Yang, Z.J.; et al. Evidence that growth hormone can improve mitochondrial function in oocytes from aged mice. Reproduction 2018, 157, 345–358. [Google Scholar] [CrossRef]
- Karlsson, J.O.; Szurek, E.A.; Higgins, A.Z.; Lee, S.R.; Eroglu, A. Optimization of cryoprotectant loading into murine and human oocytes. Cryobiology 2014, 68, 18–28. [Google Scholar] [CrossRef]
- Feitosa, W.B.; Lopes, E.; Visintin, J.A.; Assumpcao, M. Endoplasmic reticulum distribution during bovine oocyte activation is regulated by protein kinase C via actin filaments. J. Cell. Physiol. 2020, 235, 5823–5834. [Google Scholar] [CrossRef] [PubMed]
- Kline, D.; Mehlmann, L.; Fox, C.; Terasaki, M. The cortical endoplasmic reticulum (ER) of the mouse egg: Localization of ER clusters in relation to the generation of repetitive calcium waves. Dev. Biol. 1999, 215, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Wakai, T.; Fissore, R.A. Constitutive IP3R1-mediated Ca2+ release reduces Ca2+ store content and stimulates mitochondrial metabolism in mouse GV oocytes. J. Cell. Sci. 2019, 132, jcs225441. [Google Scholar] [CrossRef]
- Liu, J.; Lu, Q.; Liang, R.; Guo, J.; Wang, K.; Dong, F.; Wang, J.; Zhang, J.; Fang, J. Communicating with mouse oocytes via regulating calcium oscillation patterns by nanosecond pulsed electric fields. Phys. Rev. Appl. 2019, 11, 024001. [Google Scholar] [CrossRef]
- Bennabi, I.; Terret, M.E.; Verlhac, M.H. Meiotic spindle assembly and chromosome segregation in oocytes. J. Cell. Biol. 2016, 215, 611–619. [Google Scholar] [CrossRef]
- Li, Y.; Feng, H.L.; Cao, Y.J.; Zheng, G.J.; Yang, Y.; Mullen, S.; Critser, J.K.; Chen, Z.J. Confocal microscopic analysis of the spindle and chromosome configurations of human oocytes matured in vitro. Fertil. Steril. 2006, 85, 827–832. [Google Scholar] [CrossRef]
- Chen, S.U.; Yang, Y.S. Slow freezing or vitrification of oocytes: Their effects on survival and meiotic spindles, and the time schedule for clinical practice. Taiwan. J. Obstet. Gynecol. 2009, 48, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Yu, I. Effect of incubation time after cooling on the meiotic spindle and chromosomes of mouse oocytes. J. Emb. Trans. 2004, 19, 283–289. [Google Scholar]
- Chang, C.C.; Lin, C.J.; Sung, L.Y.; Kort, H.I.; Tian, X.C.; Nagy, Z.P. Impact of phase transition on the mouse oocyte spindle during vitrification. Reprod. Biomed. Online 2011, 22, 184–191. [Google Scholar] [CrossRef]
- Cho, I.Y.; Sheen, Y.Y. Effect of dioxin on the change of mitochondrial inner membrane potential and the induction of ROS. J. Environ. Toxicol. 2009, 24, 33–41. [Google Scholar]
- Uppangala, S.; Dhiman, S.; Salian, S.R.; Singh, V.J.; Kalthur, G.; Adiga, S.K. In vitro matured oocytes are more susceptible than in vivo matured oocytes to mock ICSI induced functional and genetic changes. PLoS ONE. 2015, 10, e0119735. [Google Scholar] [CrossRef] [PubMed]
- Mullen, S.F.; Li, M.; Li, Y.; Chen, Z.J.; Critser, J.K. Human oocyte vitrification: The permeability of metaphase II oocytes to water and ethylene glycol and the appliance toward vitrification. Fertil. Steril. 2008, 89, 1812–1825. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Q.F.; Deng, L.; Xue, S.G.; Cao, S.F.; Liu, X.Y.; Jin, W.; Wu, L.Q. New technique for mouse oocyte injection via a modified holding pipette. Reprod. Biomed. Online 2010, 21, 663–666. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, J.-R.; Yu, E.-H.; Lee, H.-J.; Kim, I.-H.; Jeong, J.-H.; Lee, D.-B.; Cho, S.-K.; Joo, J.-K. Ultra-Fast Vitrification: Minimizing the Toxicity of Cryoprotective Agents and Osmotic Stress in Mouse Oocyte Cryopreservation. Int. J. Mol. Sci. 2024, 25, 1884. https://doi.org/10.3390/ijms25031884
Cho J-R, Yu E-H, Lee H-J, Kim I-H, Jeong J-H, Lee D-B, Cho S-K, Joo J-K. Ultra-Fast Vitrification: Minimizing the Toxicity of Cryoprotective Agents and Osmotic Stress in Mouse Oocyte Cryopreservation. International Journal of Molecular Sciences. 2024; 25(3):1884. https://doi.org/10.3390/ijms25031884
Chicago/Turabian StyleCho, Jung-Ran, Eun-Hee Yu, Hyun-Joo Lee, In-Hye Kim, Ji-Hye Jeong, Dan-Bi Lee, Seong-Keun Cho, and Jong-Kil Joo. 2024. "Ultra-Fast Vitrification: Minimizing the Toxicity of Cryoprotective Agents and Osmotic Stress in Mouse Oocyte Cryopreservation" International Journal of Molecular Sciences 25, no. 3: 1884. https://doi.org/10.3390/ijms25031884