

The Specificities of Lysophosphatidic Acid Acyltransferase and Fatty Acid Desaturase Determine the High Content of Myristic and Myristoleic Acids in Cyanobacterium sp. IPPAS B-1200


Abstract
:1. Introduction
2. Results
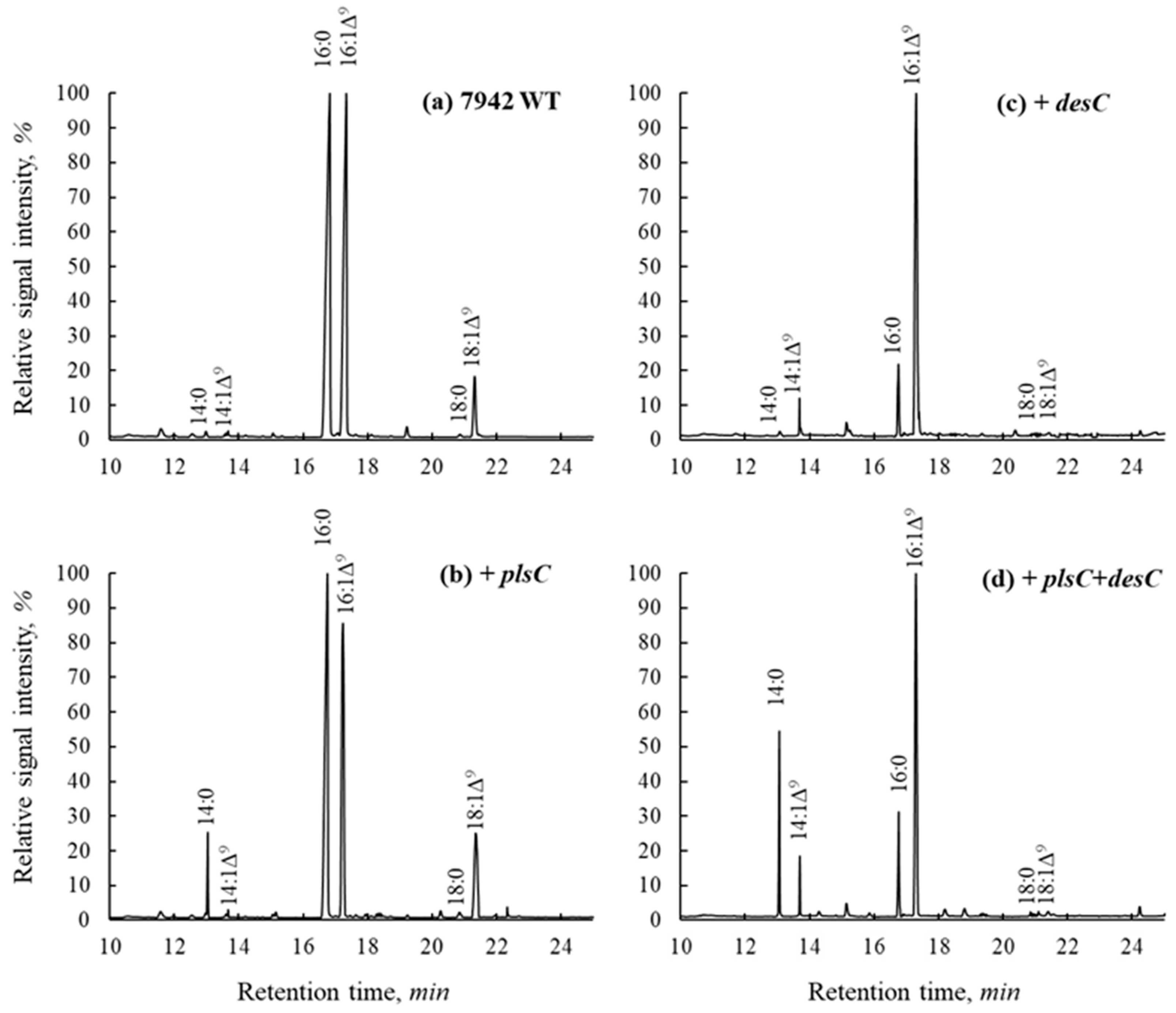
2.1. Changes in FA Composition of Total Lipids in S. elongatus Transformants
2.2. Stereospecific Positioning of C14 FAs
2.3. Analysis of FA Composition in Individual Lipid Classes
3. Discussion
4. Materials and Methods
4.1. Cyanobacterial Strains and Growth Conditions
4.2. Cloning and Expression of the desC and plsC Genes
4.3. FA Analysis of Total Lipids
4.4. GC-MS
4.5. Analysis of Lysophosphatidic and Phosphatidic Acids
4.6. Analysis of Fatty Compositions of Individual Classes of Glycerolipids
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boudière, L.; Michaud, M.; Petroutsos, D.; Rébeillé, F.; Falconet, D.; Bastien, O.; Roy, S.; Finazzi, G.; Rolland, N.; Jouhet, J.; et al. Glycerolipids in photosynthesis: Composition, synthesis and trafficking. Biochim. Biophys. Acta Bioenerg. 2014, 1837, 470–480. [Google Scholar] [CrossRef]
- Sato, N.; Wada, H. Lipid biosynthesis and its regulation in cyanobacteria. In Lipids in Photosynthesis; Advances in Photosynthesis and Respiration; Wada, H., Murata, N., Eds.; Springer: Dordrecht, The Netherlands, 2009; Volume 30, pp. 157–177. [Google Scholar] [CrossRef]
- Saito, M.; Endo, K.; Kobayashi, K.; Watanabe, M.; Ikeuchi, M.; Murakami, A.; Murata, N.; Wada, H. High myristic acid content in the cyanobacterium Cyanothece sp. PCC 8801 results from substrate specificity of lysophosphatidic acid acyltransferase. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 939–947. [Google Scholar] [CrossRef]
- Gombos, Z.; Murata, N. Lipids and fatty acids of Prochlorothrix hollandica. Plant Cell Physiol. 1991, 32, 73–77. [Google Scholar] [CrossRef]
- Sarsekeyeva, F.K.; Usserbaeva, A.A.; Zayadan, B.K.; Mironov, K.S.; Sidorov, R.A.; Kozlova, A.Y.; Kupriyanova, E.V.; Sinetova, M.A.; Los, D.A. Isolation and characterization of a new cyanobacterial strain with a unique fatty acid composition. Adv. Microbiol. 2014, 4, 1033–1043. [Google Scholar] [CrossRef]
- Shih, P.M.; Wu, D.; Latifi, A.; Axen, S.D.; Fewer, D.P.; Talla, E.; Calteau, A.; Cai, F.; Tandeau de Marsac, N.; Rippka, R.; et al. Improving the coverage of the cyanobacterial phylum using diversity-driven genome sequencing. Proc. Natl. Acad. Sci. USA 2013, 110, 1053–1058. [Google Scholar] [CrossRef]
- Starikov, A.Y.; Usserbaeva, A.A.; Sinetova, M.A.; Sarsekeyeva, F.K.; Zayadan, B.K.; Ustinova, V.V.; Kupriyanova, E.V.; Los, D.A.; Mironov, K.S. Draft genome sequence of Cyanobacterium sp. strain IPPAS B-1200 with a unique fatty acid composition. Genome Announc. 2016, 4, e01306-16. [Google Scholar] [CrossRef]
- Murata, N.; Wada, H.; Gombos, Z. Modes of fatty-acid desaturation in cyanobacteria. Plant Cell Physiol. 1992, 33, 933–941. [Google Scholar] [CrossRef]
- Santos-Merino, M.; Gargantilla-Becerra, Á.; de la Cruz, F.; Nogales, J. Highlighting the potential of Synechococcus elongatus PCC 7942 as platform to produce α-linolenic acid through an updated genome-scale metabolic modeling. Front. Microbiol. 2023, 14, 1126030. [Google Scholar] [CrossRef]
- Wada, H.; Gombos, Z.; Murata, N. Enhancement of chilling tolerance of a cyanobacterium by genetic manipulation of fatty acid desaturation. Nature 1990, 347, 200–203. [Google Scholar] [CrossRef]
- Starikov, A.Y.; Sidorov, R.A.; Mironov, K.S.; Goriainov, S.V.; Los, D.A. Delta or Omega? Δ12 (ω6) fatty acid desaturases count 3C after the pre-existing double bond. Biochimie 2020, 179, 46–53. [Google Scholar] [CrossRef]
- Starikov, A.Y.; Sidorov, R.A.; Goriainov, S.V.; Los, D.A. Acyl-lipid Δ6-desaturase may act as a first FAD in cyanobacteria. Biomolecules 2022, 12, 1795. [Google Scholar] [CrossRef]
- Brown, A.P.; Slabas, A.R.; Denton, H. Substrate selectivity of plant and microbial lysophosphatidic acid acyltransferases. Phytochemistry 2002, 61, 493–501. [Google Scholar] [CrossRef]
- Okazaki, K.; Sato, N.; Tsuji, N.; Tsuzuki, M.; Nishida, I. The significance of C16 fatty acids in the sn-2 positions of glycerolipids in the photosynthetic growth of Synechocystis sp. PCC 6803. Plant Physiol. 2006, 141, 546–556. [Google Scholar] [CrossRef]
- Ogawa, T.; Kuboshima, M.; Suwanawat, N.; Kawamoto, J.; Kurihara, T. Division of the role and physiological impact of multiple lysophosphatidic acid acyltransferase paralogs. BMC Microbiol. 2022, 22, 241. [Google Scholar] [CrossRef]
- Ogawa, T.; Suwanawat, N.; Toyotake, Y.; Watanabe, B.; Kawamoto, J.; Kurihara, T. Lysophosphatidic acid acyltransferase from the thermophilic bacterium Thermus thermophilus HB8 displays substrate promiscuity. Biosci. Biotechnol. Biochem. 2020, 84, 1831–1838. [Google Scholar] [CrossRef]
- Wada, H.; Murata, N. Temperature-induced changes in the fatty acid composition of the cyanobacterium, Synechocystis PCC6803. Plant Physiol. 1990, 92, 1062–1069. [Google Scholar] [CrossRef]
- Khan, M.U.; Mackenzie, S.L.; Williams, J.P. Modulation of fatty acids in the membranes of Anacystis nidulans (cyanobacteria): Incorporation of odd-numbered carbon fatty acids. J. Phycol. 1996, 32, 970–973. [Google Scholar] [CrossRef]
- Wang, H.; Klein, M.G.; Zou, H.; Lane, W.; Snell, G.; Levin, I.; Li, K.; Sang, B.C. Crystal structure of human stearoyl-coenzyme A desaturase in complex with substrate. Nat. Struct. Mol. Biol. 2015, 22, 581–585. [Google Scholar] [CrossRef]
- Bai, Y.; McCoy, J.G.; Levin, E.J.; Sobrado, P.; Rajashankar, K.R.; Fox, B.G.; Zhou, M. X-ray structure of a mammalian stearoyl-CoA desaturase. Nature 2015, 524, 252–256. [Google Scholar] [CrossRef]
- Li, D.; Moorman, R.; Vanhercke, T.; Petrie, J.; Singh, S.; Jackson, C.J. Classification and substrate head-group specificity of membrane fatty acid desaturases. Comput. Struct. Biotechnol. J. 2016, 14, 341–349. [Google Scholar] [CrossRef]
- Maslova, I.P.; Mouradyan, E.A.; Lapina, S.S.; Klyachko-Gurvich, G.L.; Los, D.A. Lipid fatty acid composition and thermophilicity of cyanobacteria. Russ. J. Plant Physiol. 2004, 51, 353–360. [Google Scholar] [CrossRef]
- Meinnel, T.; Dian, C.; Giglione, C. Myristoylation, an ancient protein modification mirroring eukaryogenesis and evolution. Trends Biochem. Sci. 2020, 45, 619–632. [Google Scholar] [CrossRef] [PubMed]
- Ameye, L.G.; Chee, W.S. Osteoarthritis and nutrition. From nutraceuticals to functional foods: A systematic review of the scientific evidence. Arthritis Res. Ther. 2006, 8, R127. [Google Scholar] [CrossRef] [PubMed]
- Obranović, M.; Bryś, J.; Repajić, M.; Balbino, S.; Škevin, D.; Bryś, A.; Tonković, P.; Medved, A.M.; Uzelac, V.D.; Kraljić, K. Fatty acid and sterol profile of nutmeg (Myristica fragrans) and star anise (Illicium verum) extracted using three different methods. Proceedings 2021, 70, 33. [Google Scholar] [CrossRef]
- Melis, A.; Hidalgo Martinez, D.A.; Betterle, N. Perspectives of cyanobacterial cell factories. Photosynth. Res. 2023. [Google Scholar] [CrossRef]
- Li, Z.; Li, S.; Chen, L.; Sun, T.; Zhang, W. Fast-growing cyanobacterial chassis for synthetic biology application. Crit. Rev. Biotechnol. 2023. [Google Scholar] [CrossRef]
- Rippka, R. Isolation and purification of cyanobacteria. Methods Enzymol. 1988, 167, 3–27. [Google Scholar] [CrossRef]
- Williams, J.G.K. Construction of specific mutations in photosystem II photosynthetic reaction center by genetic engineering methods in Synechocystis 6803. Methods Enzymol. 1988, 167, 766–778. [Google Scholar] [CrossRef]
- Andersson, C.R.; Tsinoremas, N.F.; Shelton, J.; Lebedeva, N.V.; Yarrow, J.; Min, H.; Golden, S.S. Application of bioluminescence to the study of circadian rhythms in cyanobacteria. Methods Enzymol. 2000, 305, 527–542. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Pinault, M.; Guimaraes, C.; Dumas, J.; Servais, S.; Chevalier, S.; Besson, P.; Goupille, C. A 1D high performance thin layer chromatography method validated to quantify phospholipids including cardiolipin and monolysocardiolipin from biological samples. Eur. J. Lipid Sci. Technol. 2020, 122, 1900240. [Google Scholar] [CrossRef]
- Sallal, A.K.; Nimer, N.A.; Radwan, S.S. Lipid and fatty acid composition of freshwater cyanobacteria. J. Gen. Microbiol. 1990, 136, 2043–2048. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| FAs | 7942 | plsC | desC | plsC + desC |
|---|---|---|---|---|
| 14:0 | 0.5 | 10.2 | 0.6 | 26.5 |
| 14:1∆9 | 0.5 | 0.8 | 9.0 | 9.0 |
| 16:0 | 50.3 | 40.4 | 16.0 | 15.2 |
| 16:1∆9 | 31.7 | 34.6 | 73.1 | 48.5 |
| 18:0 | 1.2 | 2.6 | 0.6 | 0.2 |
| 18:1∆9 | 13.0 | 9.4 | 0.5 | 0.5 |
| 18:1∆11 | 2.8 | 2.0 | 0.2 | 0.1 |
| WT | PlsC | |||
|---|---|---|---|---|
| FAs | LPA | PA | LPA | PA |
| 14:0 | 0.8 | 0.6 | 3 | 12.3 |
| 14:1Δ9 | * | 1 | 0.5 | 1.2 |
| 16:0 | 57.4 | 49.4 | 53.6 | 38 |
| 16:1Δ9 | 4 | 32.3 | 4.8 | 35.2 |
| 18:0 | 37.1 | 9 | 37.5 | 8 |
| 18:1Δ9 | 0.7 | 7.7 | 0.6 | 5.3 |
| WT | PlsC | |||||||
|---|---|---|---|---|---|---|---|---|
| FAs | MGDG | DGDG | SQDG | PG | MGDG | DGDG | SQDG | PG |
| 14:0 | 1.2 | 2.3 | 1.1 | 0.9 | 14.6 | 19.7 | 7.6 | 7.9 |
| 14:1Δ9 | 0.1 | 1.1 | 0.8 | 1.0 | 1.2 | 0.8 | 0.6 | 1.7 |
| 16:0 | 48.1 | 40.3 | 56.2 | 49.5 | 31.2 | 30.9 | 48.7 | 41.9 |
| 16:1Δ9 | 46.2 | 42.9 | 34.3 | 32.3 | 48.1 | 43.4 | 35.5 | 33.0 |
| 18:0 | 0.8 | 10.7 | 6.0 | 5.2 | 1.1 | 2.1 | 3.2 | 4.7 |
| 18:1Δ9 | 3.6 | 2.7 | 1.6 | 11.1 | 3.8 | 3.1 | 4.4 | 10.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Starikov, A.Y.; Sidorov, R.A.; Mironov, K.S.; Los, D.A. The Specificities of Lysophosphatidic Acid Acyltransferase and Fatty Acid Desaturase Determine the High Content of Myristic and Myristoleic Acids in Cyanobacterium sp. IPPAS B-1200. Int. J. Mol. Sci. 2024, 25, 774. https://doi.org/10.3390/ijms25020774
Starikov AY, Sidorov RA, Mironov KS, Los DA. The Specificities of Lysophosphatidic Acid Acyltransferase and Fatty Acid Desaturase Determine the High Content of Myristic and Myristoleic Acids in Cyanobacterium sp. IPPAS B-1200. International Journal of Molecular Sciences. 2024; 25(2):774. https://doi.org/10.3390/ijms25020774
Chicago/Turabian StyleStarikov, Alexander Y., Roman A. Sidorov, Kirill S. Mironov, and Dmitry A. Los. 2024. "The Specificities of Lysophosphatidic Acid Acyltransferase and Fatty Acid Desaturase Determine the High Content of Myristic and Myristoleic Acids in Cyanobacterium sp. IPPAS B-1200" International Journal of Molecular Sciences 25, no. 2: 774. https://doi.org/10.3390/ijms25020774