

Supplementary Low Far-Red Light Promotes Proliferation and Photosynthetic Capacity of Blueberry In Vitro Plantlets
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of Supplementing FR Light on Morphology
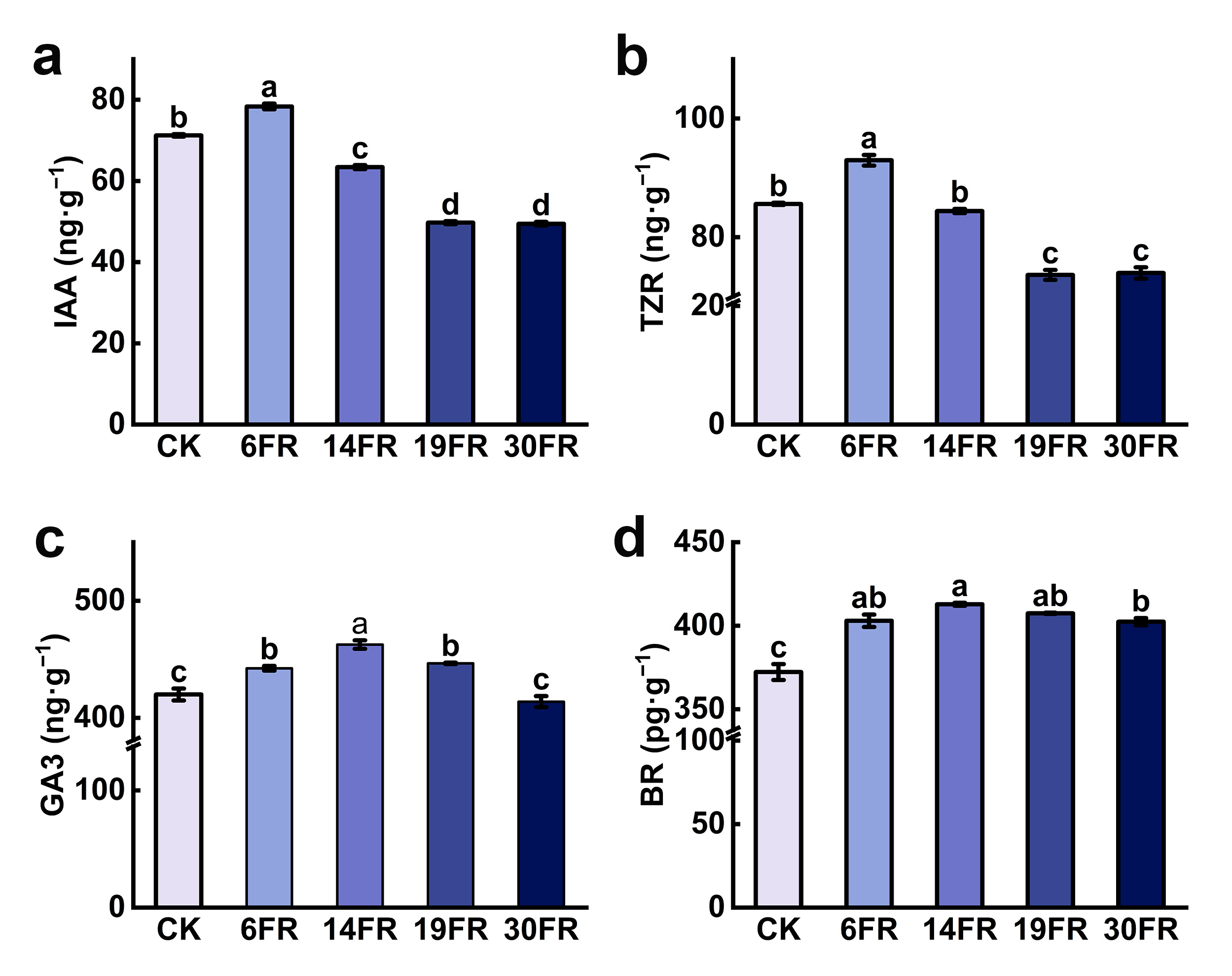
2.2. Effects of Supplementing FR Light on Endogenous Hormone Levels
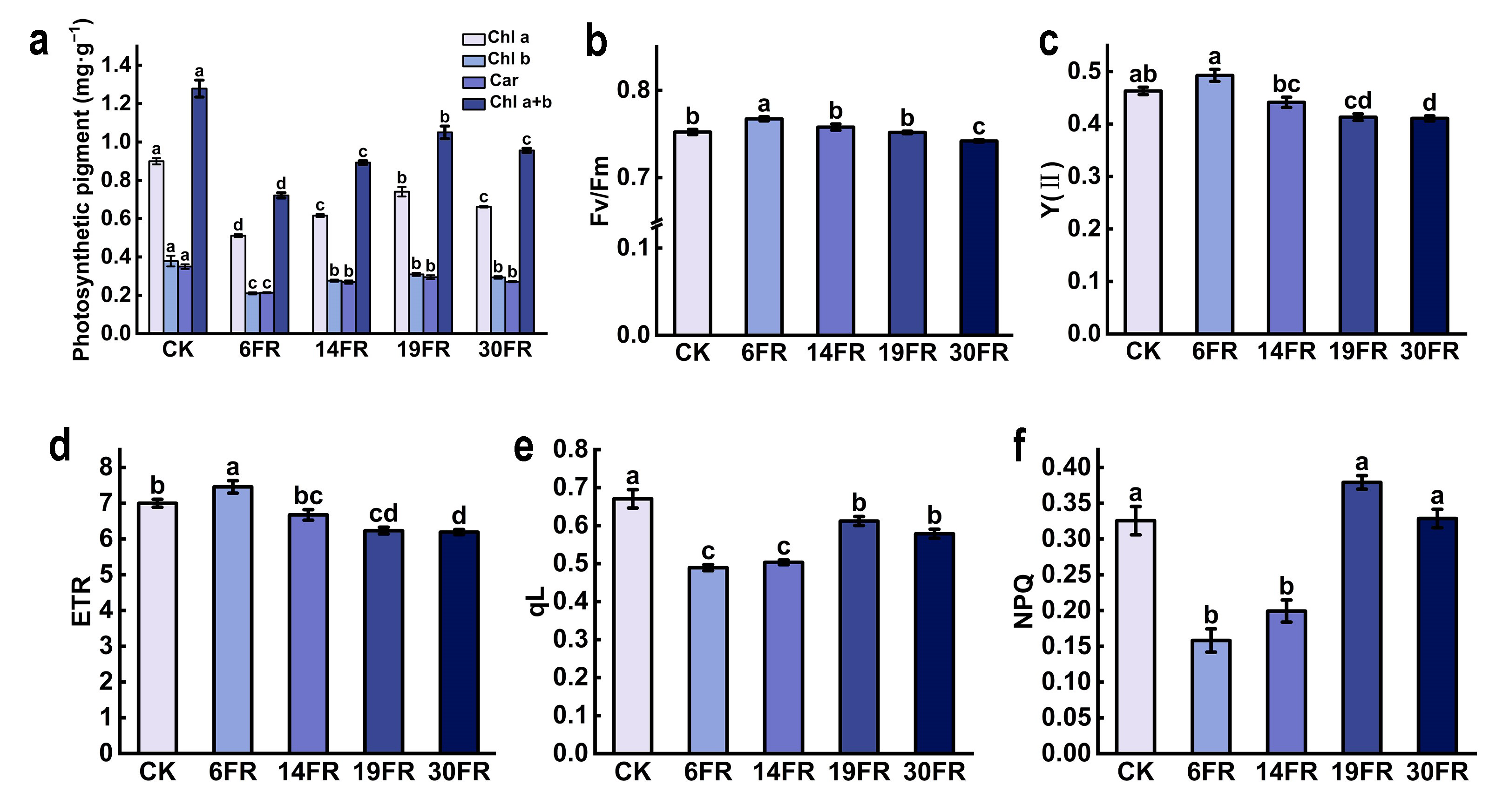
2.3. Effects of Supplementing FR Light on Photosynthetic Traits
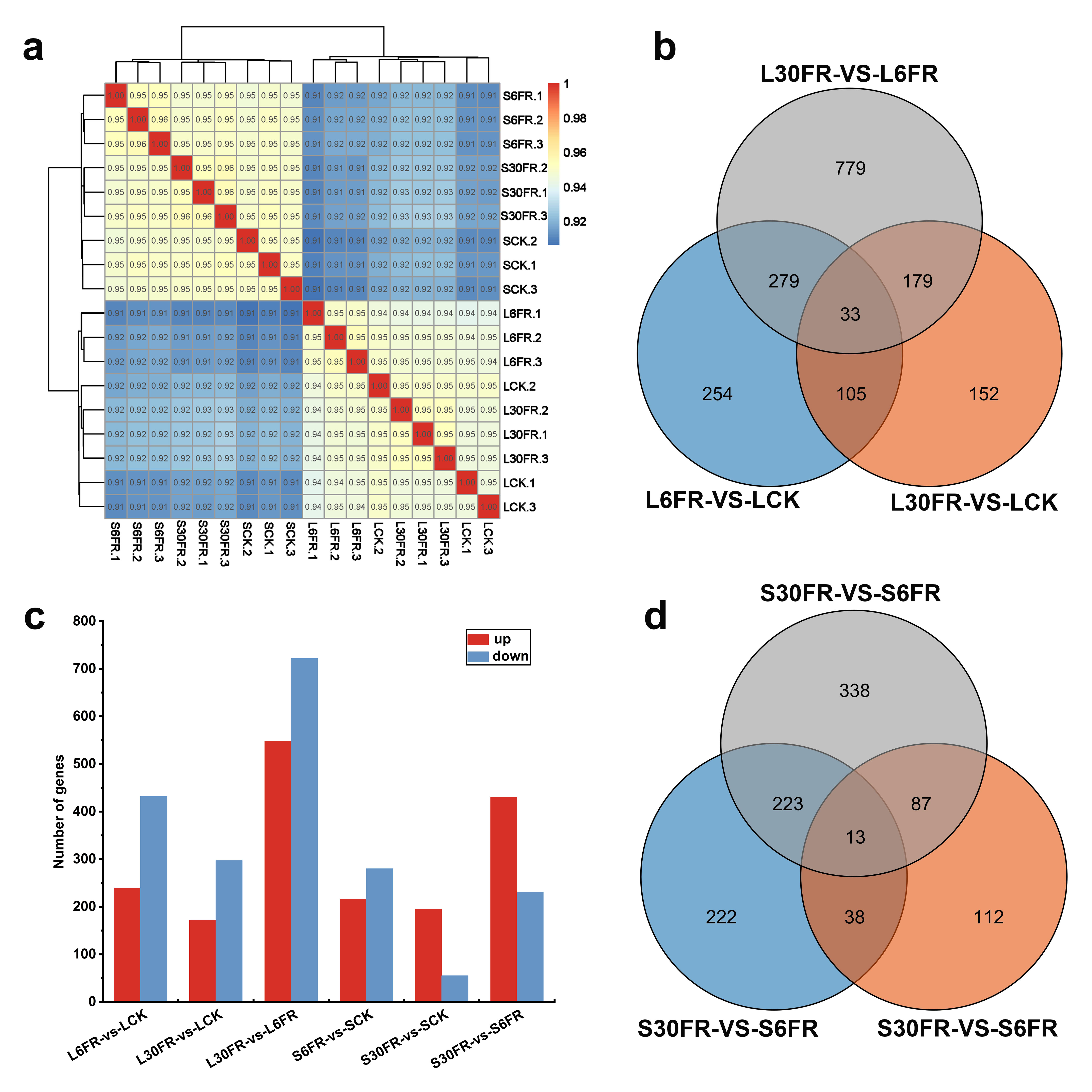
2.4. Transcriptome Sequencing
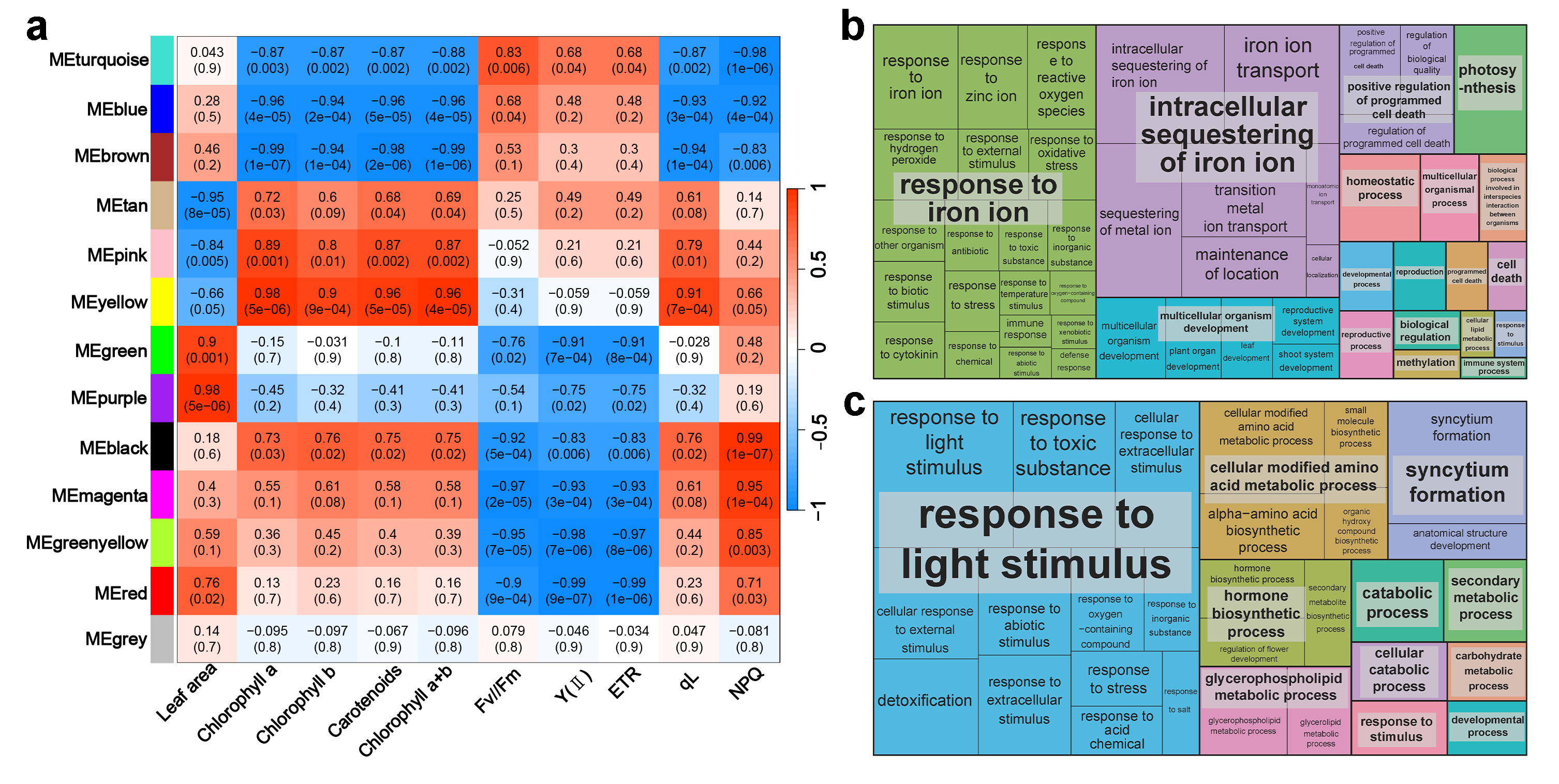
2.5. Co-Expression Network Analysis and Gene Ontology (GO) Analysis Shows the Impacts of FR Light on Proliferation
2.6. Co-Expression Network Analysis and GO Analysis Shows the Impacts of FR Light on Photosynthetic Traits
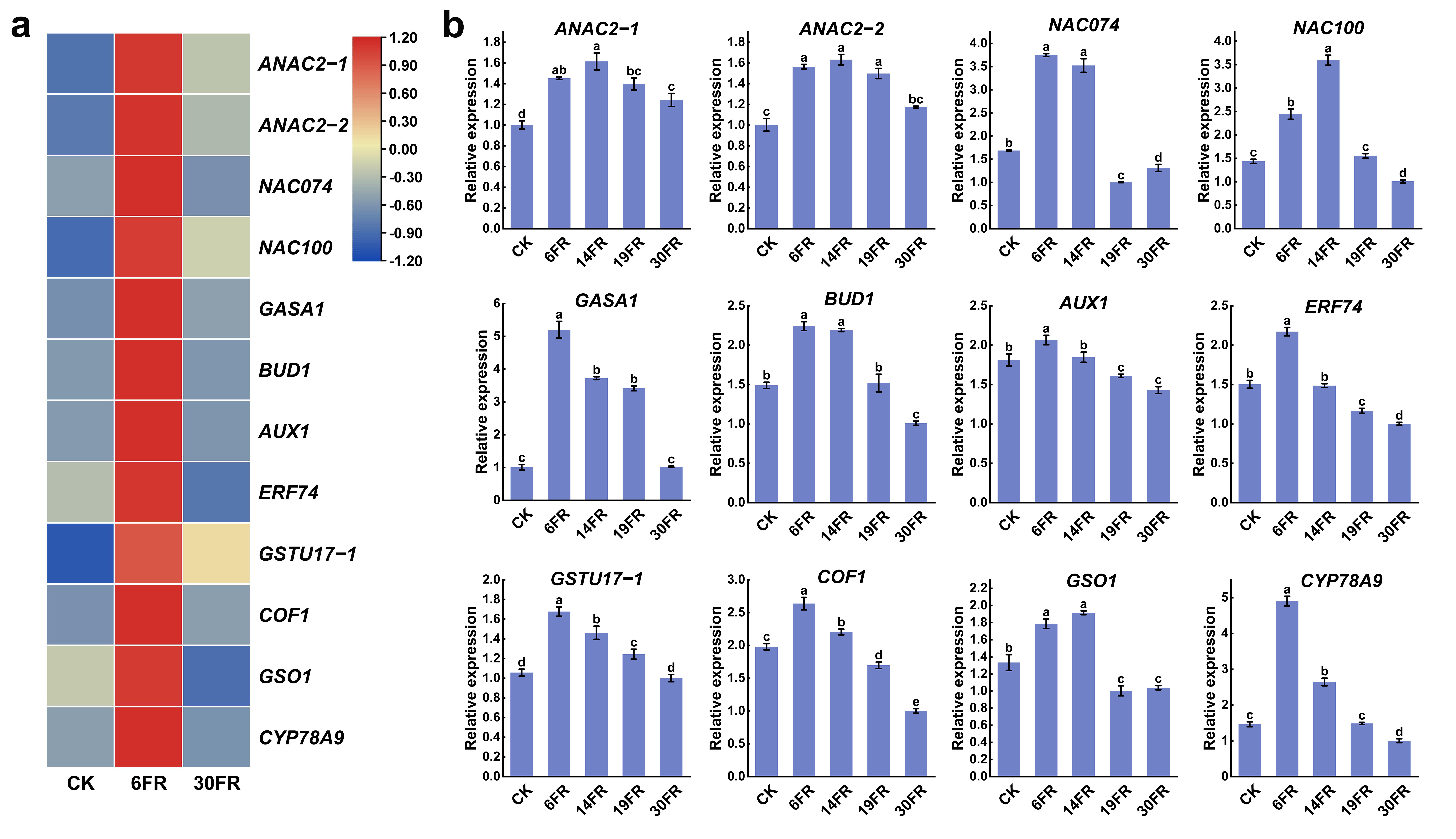
2.7. Analysis of Key Gene Expression Levels in Leaves and RT-qPCR Validation
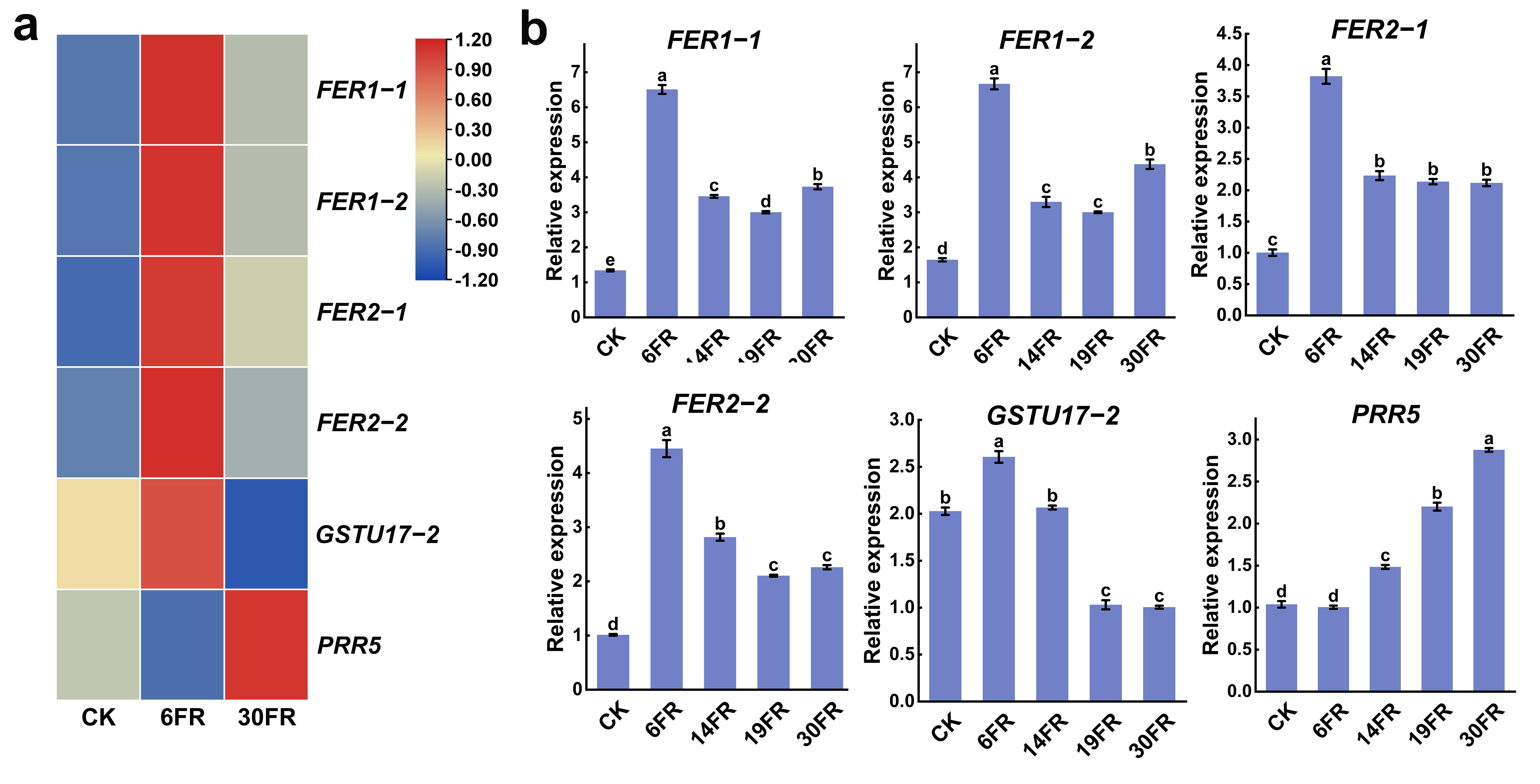
2.8. Analysis of Key Gene Expression Levels in Stems and RT-qPCR Validation
3. Discussion
3.1. Impacts of Supplemental FR Light on the Photosynthetic Characteristics and Proliferation of Blueberry Tissue Cultured Seedlings
3.2. Transcriptome Analysis Reveals Potential Regulatory Mechanisms of FR Light on Blueberry Proliferation
3.3. Transcriptome Analysis Reveals Potential Rregulatory Mechanisms of FR Light on the Photosynthetic Capacity of Blueberries
4. Materials and Methods
4.1. Plant Materials, Growth Conditions, and Light Experimental Design
4.2. Chlorophyll Content and Chlorophyll Fluorescence Measurements
4.3. Endogenous IAA, BR, GA3, and TZR Determination
4.4. RNA Extraction, cDNA Library Construction, and RNA-seq Analysis
4.5. WGCNA and GO Enrichment Analysis
4.6. Real-Time Quantitative PCR (RT-qPCR)
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shafiq, I.; Hussain, S.; Raza, M.A.; Iqbal, N.; Asghar, M.A.; Raza, A.; Fan, Y.; Mumtaz, M.; Shoaib, M.; Ansar, M.; et al. Crop Photosynthetic Response to Light Quality and Light Intensity. J. Integr. Agric. 2021, 20, 4–23. [Google Scholar] [CrossRef]
- Sullivan, J.A.; Deng, X.W. From Seed to Seed: The Role of Photoreceptors in Arabidopsis Development. Dev. Biol. 2003, 260, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Lau, O.S.; Deng, X.W. Light-Regulated Transcriptional Networks in Higher Plants. Nat. Rev. Genet. 2007, 8, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Zhen, S.; Bugbee, B. Far-red Photons Have Equivalent Efficiency to Traditional Photosynthetic Photons: Implications for Redefining Photosynthetically Active Radiation. Plant Cell Environ. 2020, 43, 1259–1272. [Google Scholar] [CrossRef]
- Zhen, S.; Van Iersel, M.; Bugbee, B. Why Far-Red Photons Should Be Included in the Definition of Photosynthetic Photons and the Measurement of Horticultural Fixture Efficacy. Front. Plant Sci. 2021, 12, 693445. [Google Scholar] [CrossRef]
- Liu, J.; Van Iersel, M.W. Far-Red Photons Increase Light Capture but Have Lower Photosynthetic Capacity Than Red Photons. J. Am. Soc. Hortic. 2023, 148, 253–265. [Google Scholar] [CrossRef]
- Laisk, A.; Oja, V.; Eichelmann, H.; Dall’Osto, L. Action Spectra of Photosystems II and I and Quantum Yield of Photosynthesis in Leaves in State 1. Biochim. Biophys. Acta 2014, 1837, 315–325. [Google Scholar] [CrossRef]
- Hogewoning, S.W.; Wientjes, E.; Douwstra, P.; Trouwborst, G.; Van Ieperen, W.; Croce, R.; Harbinson, J. Photosynthetic Quantum Yield Dynamics: From Photosystems to Leaves. Plant Cell 2012, 24, 1921–1935. [Google Scholar] [CrossRef]
- Franklin, K.A.; Quail, P.H. Phytochrome Functions in Arabidopsis Development. J. Exp. Bot. 2010, 61, 11–24. [Google Scholar] [CrossRef]
- Casal, J.J. Shade Avoidance. Arab. Book 2012, 10, e0157. [Google Scholar] [CrossRef]
- Simkin, A.J.; Kapoor, L.; Doss, C.G.P.; Hofmann, T.A.; Lawson, T.; Ramamoorthy, S. The Role of Photosynthesis Related Pigments in Light Harvesting, Photoprotection and Enhancement of Photosynthetic Yield in Planta. Photosynth. Res. 2022, 152, 23–42. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.R. Chlorophyll Fluorescence: A Probe of Photosynthesis In Vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ning, Z.; Awan, S.A.; Gao, J.; Chen, J.; Lei, Y.; Tan, X.; Wu, X.; Wu, Y.; Liu, C.; et al. Far-Red Light Mediates Light Energy Capture and Distribution in Soybeans (Glycine Max L.) under the Shade. Plant Physiol. Biochem. 2023, 204, 108130. [Google Scholar] [CrossRef]
- Kim, H.-J.; Lin, M.-Y.; Mitchell, C.A. Light Spectral and Thermal Properties Govern Biomass Allocation in Tomato through Morphological and Physiological Changes. Environ. Exp. Bot. 2019, 157, 228–240. [Google Scholar] [CrossRef]
- Zou, J.; Zhang, Y.; Zhang, Y.; Bian, Z.; Fanourakis, D.; Yang, Q.; Li, T. Morphological and Physiological Properties of Indoor Cultivated Lettuce in Response to Additional Far-Red Light. Sci. Hortic. 2019, 257, 108725. [Google Scholar] [CrossRef]
- Laisk, A.; Talts, E.; Oja, V.; Eichelmann, H.; Peterson, R.B. Fast Cyclic Electron Transport around Photosystem I in Leaves under Far-Red Light: A Proton-Uncoupled Pathway? Photosyn. Res. 2010, 103, 79–95. [Google Scholar] [CrossRef]
- Kono, M.; Yamori, W.; Suzuki, Y.; Terashima, I. Photoprotection of PSI by Far-Red Light Against the Fluctuating Light-Induced Photoinhibition in Arabidopsis Thaliana and Field-Grown Plants. Plant Cell Physiol. 2017, 58, 35–45. [Google Scholar] [CrossRef]
- Zhen, S.; Van Iersel, M.W. Far-Red Light Is Needed for Efficient Photochemistry and Photosynthesis. J. Plant Physiol. 2017, 209, 115–122. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, H.; Gao, M.; He, R.; Liu, X.; Su, W.; Liu, H. Far-Red-Light-Induced Morphology Changes, Phytohormone, and Transcriptome Reprogramming of Chinese Kale (Brassica Alboglabra Bailey). Int. J. Mol. Sci. 2023, 24, 5563. [Google Scholar] [CrossRef]
- Procko, C.; Crenshaw, C.M.; Ljung, K.; Noel, J.P.; Chory, J. Cotyledon-Generated Auxin Is Required for Shade-Induced Hypocotyl Growth in Brassica Rapa. Plant Physiol. 2014, 165, 1285–1301. [Google Scholar] [CrossRef]
- Lei, K.; Tan, Q.; Zhu, L.; Xu, L.; Yang, S.; Hu, J.; Gao, L.; Hou, P.; Shao, Y.; Jiang, D.; et al. Low Red/Far-Red Ratio Can Induce Cytokinin Degradation Resulting in the Inhibition of Tillering in Wheat (Triticum Aestivum L.). Front. Plant Sci. 2022, 13, 971003. [Google Scholar] [CrossRef] [PubMed]
- Ballaré, C.L.; Sánchez, R.A.; Scopel, A.L.; Casal, J.J.; Ghersa, C.M. Early Detection of Neighbour Plants by Phytochrome Perception of Spectral Changes in Reflected Sunlight. Plant Cell Environ. 1987, 10, 551–557. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass Allocation to Leaves, Stems and Roots: Meta-analyses of Interspecific Variation and Environmental Control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.K.; Holalu, S.V.; Casal, J.J.; Finlayson, S.A. Abscisic Acid Regulates Axillary Bud Outgrowth Responses to the Ratio of Red to Far-Red Light. Plant Physiol. 2013, 163, 1047–1058. [Google Scholar] [CrossRef]
- Yuan, C.; Ahmad, S.; Cheng, T.; Wang, J.; Pan, H.; Zhao, L.; Zhang, Q. Red to Far-Red Light Ratio Modulates Hormonal and Genetic Control of Axillary Bud Outgrowth in Chrysanthemum (Dendranthema Grandiflorum ‘Jinba’). Int. J. Mol. Sci. 2018, 19, 1590. [Google Scholar] [CrossRef]
- Li, W.; Liu, S.W.; Ma, J.J.; Liu, H.M.; Han, F.X.; Li, Y.; Niu, S.-H. Gibberellin Signaling Is Required for Far-Red Light-Induced Shoot Elongation in Pinus Tabuliformis Seedlings. Plant Physiol. 2020, 182, 658–668. [Google Scholar] [CrossRef]
- Tan, T.; Li, S.; Fan, Y.; Wang, Z.; Ali Raza, M.; Shafiq, I.; Wang, B.; Wu, X.; Yong, T.; Wang, X.; et al. Far-Red Light: A Regulator of Plant Morphology and Photosynthetic Capacity. Crop J. 2022, 10, 300–309. [Google Scholar] [CrossRef]
- Hornitschek, P.; Kohnen, M.V.; Lorrain, S.; Rougemont, J.; Ljung, K.; López-Vidriero, I.; Franco-Zorrilla, J.M.; Solano, R.; Trevisan, M.; Pradervand, S.; et al. Phytochrome Interacting Factors 4 and 5 Control Seedling Growth in Changing Light Conditions by Directly Controlling Auxin Signaling. Plant J. 2012, 71, 699–711. [Google Scholar] [CrossRef]
- Dünser, K.; Kleine-Vehn, J. Differential Growth Regulation in Plants—The Acid Growth Balloon Theory. Curr. Opin. Plant Biol. 2015, 28, 55–59. [Google Scholar] [CrossRef]
- Ballaré, C.L.; Pierik, R. The Shade-avoidance Syndrome: Multiple Signals and Ecological Consequences. Plant Cell Environ. 2017, 40, 2530–2543. [Google Scholar] [CrossRef]
- Hornitschek, P.; Lorrain, S.; Zoete, V.; Michielin, O.; Fankhauser, C. Inhibition of the Shade Avoidance Response by Formation of Non-DNA Binding bHLH Heterodimers. EMBO J. 2009, 28, 3893–3902. [Google Scholar] [CrossRef] [PubMed]
- Rose, J.K.C.; Braam, J.; Fry, S.C.; Nishitani, K. The XTH Family of Enzymes Involved in Xyloglucan Endotransglucosylation and Endohydrolysis: Current Perspectives and a New Unifying Nomenclature. Plant Cell Physiol. 2002, 43, 1421–1435. [Google Scholar] [CrossRef] [PubMed]
- De Wit, M.; Kegge, W.; Evers, J.B.; Vergeer-van Eijk, M.H.; Gankema, P.; Voesenek, L.A.C.J.; Pierik, R. Plant Neighbor Detection through Touching Leaf Tips Precedes Phytochrome Signals. Proc. Natl. Acad. Sci. USA 2012, 109, 14705–14710. [Google Scholar] [CrossRef]
- Sasidharan, R.; Keuskamp, D.H.; Kooke, R.; Voesenek, L.A.C.J.; Pierik, R. Interactions between Auxin, Microtubules and XTHs Mediate Green Shade- Induced Petiole Elongation in Arabidopsis. PLoS ONE 2014, 9, e90587. [Google Scholar] [CrossRef]
- Fang, Y.; Nunez, G.H.; Silva, M.N.D.; Phillips, D.A.; Munoz, P.R. A Review for Southern Highbush Blueberry Alternative Production Systems. Agronomy 2020, 10, 1531. [Google Scholar] [CrossRef]
- Meiners, J.; Schwab, M.; Szankowski, I. Efficient in Vitro Regeneration Systems for Vaccinium Species. Plant Cell Tissue Organ Cult. 2007, 89, 169–176. [Google Scholar] [CrossRef]
- Fan, S.; Jian, D.; Wei, X.; Chen, J.; Beeson, R.C.; Zhou, Z.; Wang, X. Micropropagation of Blueberry ‘Bluejay’ and ‘Pink Lemonade’ through In Vitro Shoot Culture. Sci. Hortic. 2017, 226, 277–284. [Google Scholar] [CrossRef]
- Paradiso, R.; Proietti, S. Light-Quality Manipulation to Control Plant Growth and Photomorphogenesis in Greenhouse Horticulture: The State of the Art and the Opportunities of Modern LED Systems. J. Plant Growth Regul. 2022, 41, 742–780. [Google Scholar] [CrossRef]
- Zhao, J.; Thi, L.T.; Park, Y.G.; Jeong, B.R. Light Quality Affects Growth and Physiology of Carpesium Triste Maxim. Cultured In Vitro. Agriculture 2020, 10, 258. [Google Scholar] [CrossRef]
- Zhang, D.; Liu, Y.; Ni, C.; Chen, J. Effect of LED Light on the Growth and Physiological Indices of Blueberry. Agron. J. 2022, 114, 2105–2112. [Google Scholar] [CrossRef]
- An, H.; Zhang, J.; Zhang, L.; Li, S.; Zhou, B.; Zhang, X. Effects of Nutrition and Light Quality on the Growth of Southern Highbush Blueberry (Vaccinium Corymbosum L.) in an Advanced Plant Factory with Artificial Lighting (PFAL). Horticulturae 2023, 9, 287. [Google Scholar] [CrossRef]
- Zhang, J.; Li, S.; An, H.; Zhang, X.; Zhou, B. Integrated Transcriptome and Metabolome Analysis Reveals the Anthocyanin Biosynthesis Mechanisms in Blueberry (Vaccinium Corymbosum L.) Leaves under Different Light Qualities. Front. Plant Sci. 2022, 13, 1073332. [Google Scholar] [CrossRef]
- Gu, J.; Zhou, Z.; Li, Z.; Chen, Y.; Wang, Z.; Zhang, H.R.R. Rice (Oryza Sativa L.) with Reduced Chlorophyll Content Exhibit Higher Photosynthetic Rate and Efficiency, Improved Canopy Light Distribution, and Greater Yields than Normally Pigmented Plants. Field Crops Res. 2017, 200, 58–70. [Google Scholar] [CrossRef]
- Liu, Y.; Jafari, F.; Wang, H. Integration of Light and Hormone Signaling Pathways in the Regulation of Plant Shade Avoidance Syndrome. aBIOTECH 2021, 2, 131–145. [Google Scholar] [CrossRef]
- Liu, H.; Yang, C.; Li, L. Shade-induced Stem Elongation in Rice Seedlings: Implication of Tissue-specific Phytohormone Regulation. J. Integr. Plant Biol. 2016, 58, 614–617. [Google Scholar] [CrossRef]
- Shin, J.; Park, P.B. Shade Avoidance and the Regulation of Leaf Inclination in Rice. Rapid Commun. Photosci. 2014, 3, 53–55. [Google Scholar] [CrossRef]
- Zhu, Y.; Nomura, T.; Xu, Y.; Zhang, Y.; Peng, Y.; Mao, B.; Hanada, A.; Zhou, H.; Wang, R.; Li, P.; et al. Elongated Uppermost Internode Encodes a Cytochrome P450 Monooxygenase that Epoxidizes Gibberellins in a Novel Deactivation Reaction in Rice. Plant Cell 2006, 18, 442–456. [Google Scholar] [CrossRef]
- Yamamuro, C.; Ihara, Y.; Wu, X.; Noguchi, T.; Fujioka, S.; Takatsuto, S.; Ashikari, M.; Kitano, H.; Matsuoka, M. Loss of Function of a Rice Brassinosteroid Insensitive1 Homolog Prevents Internode Elongation and Bending of the Lamina Joint. Plant Cell 2000, 12, 1591–1606. [Google Scholar] [CrossRef]
- Song, X.; Gu, X.; Chen, S.; Qi, Z.; Yu, J.; Zhou, Y.; Xia, X. Far-red Light Inhibits Lateral Bud Growth Mainly through Enhancing Apical Dominance Independently of Strigolactone Synthesis in Tomato. Plant Cell Environ. 2023; online ahead of print. [Google Scholar] [CrossRef]
- Feng, G.; Wu, J.; Xu, Y.; Lu, L.; Yi, H. High-spatiotemporal-resolution Transcriptomes Provide Insights into Fruit Development and Ripening in Citrus Sinensis. Plant Biotechnol. J. 2021, 19, 1337–1353. [Google Scholar] [CrossRef]
- Wang, P.; Chen, S.; Gu, M.; Chen, X.; Chen, X.; Yang, J.; Zhao, F.; Ye, N. Exploration of the Effects of Different Blue LED Light Intensities on Flavonoid and Lipid Metabolism in Tea Plants via Transcriptomics and Metabolomics. Int. J. Mol. Sci. 2020, 21, 4606. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yang, H.; Ma, Y.; Jiang, G.; Mei, X.; Li, X.; Yang, Q.; Kan, J.; Xu, Y.; Yang, T.; et al. WGCNA Analysis Revealing Molecular Mechanism That Bio-Organic Fertilizer Improves Pear Fruit Quality by Increasing Sucrose Accumulation and Reducing Citric Acid Metabolism. Front. Plant Sci. 2022, 13, 1039671. [Google Scholar] [CrossRef] [PubMed]
- Taylor-Teeples, M.; Lin, L.; De Lucas, M.; Turco, G.; Toal, T.W.; Gaudinier, A.; Young, N.F.; Trabucco, G.M.; Veling, M.T.; Lamothe, R.; et al. An Arabidopsis Gene Regulatory Network for Secondary Cell Wall Synthesis. Nature 2015, 517, 571–575. [Google Scholar] [CrossRef]
- Han, K.; Zhao, Y.; Sun, Y.; Li, Y. NACs, Generalist in Plant Life. Plant Biotechnol. J. 2023, 21, 2433–2457. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Lee, C.; McCarthy, R.L.; Reeves, C.K.; Jones, E.G.; Ye, Z.H. Transcriptional Activation of Secondary Wall Biosynthesis by Rice and Maize NAC and MYB Transcription Factors. Plant Cell Physiol. 2011, 52, 1856–1871. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Lee, C.; Ye, Z.H. Functional Characterization of Poplar Wood-Associated NAC Domain Transcription Factors. Plant Physiol. 2010, 152, 1044–1055. [Google Scholar] [CrossRef]
- Zhong, R.; Richardson, E.A.; Ye, Z.H. The MYB46 Transcription Factor Is a Direct Target of SND1 and Regulates Secondary Wall Biosynthesis in Arabidopsis. Plant Cell 2007, 19, 2776–2792. [Google Scholar] [CrossRef]
- Zhong, R.; Lee, C.; Haghighat, M.; Ye, Z.H. Xylem Vessel-specific SND5 and Its Homologs Regulate Secondary Wall Biosynthesis through Activating Secondary Wall NAC Binding Elements. New Phytol. 2021, 231, 1496–1509. [Google Scholar] [CrossRef]
- Zhong, C.; Xu, H.; Ye, S.; Wang, S.; Li, L.; Zhang, S.; Wang, X. AtGASA6 Serves as an Integrator of Gibberellin-, Abscisic Acid- and Glucose-Signaling during Seed Germination in Arabidopsis. Plant Physiol. 2015, 169, 2288–2303. [Google Scholar] [CrossRef]
- Nahirñak, V.; Almasia, N.I.; Fernandez, P.V.; Hopp, H.E.; Estevez, J.M.; Carrari, F.; Vazquez-Rovere, C. Potato Snakin-1 Gene Silencing Affects Cell Division, Primary Metabolism, and Cell Wall Composition. Plant Physiol. 2012, 158, 252–263. [Google Scholar] [CrossRef]
- Han, M.; Jin, X.; Yao, W.; Kong, L.; Huang, G.; Tao, Y.; Li, L.; Wang, X.; Wang, Y. Mini Zinc-Finger Protein (MIF) from Gerbera Hybrida Activates the GASA Protein Family Gene, GEG, to Inhibit Ray Petal Elongation. Front. Plant Sci. 2017, 8, 1649. [Google Scholar] [CrossRef]
- Huang, G.; Han, M.; Jian, L.; Chen, Y.; Sun, S.; Wang, X.; Wang, Y. An ETHYLENE INSENSITIVE3-LIKE1 Protein Directly Targets the GEG Promoter and Mediates Ethylene-Induced Ray Petal Elongation in Gerbera Hybrida. Front. Plant Sci. 2019, 10, 1737. [Google Scholar] [CrossRef]
- Dai, Y.; Wang, H.; Li, B.; Huang, J.; Liu, X.; Zhou, Y.; Mou, Z.; Li, J. Increased Expression of MAP KINASE KINASE7 Causes Deficiency in Polar Auxin Transport and Leads to Plant Architectural Abnormality in Arabidopsis. Plant Cell 2006, 18, 308–320. [Google Scholar] [CrossRef]
- Swarup, R.; Bhosale, R. Developmental Roles of AUX1/LAX Auxin Influx Carriers in Plants. Front. Plant Sci. 2019, 10, 1306. [Google Scholar] [CrossRef]
- Vosolsobě, S.; Skokan, R.; Petrášek, J. The Evolutionary Origins of Auxin Transport: What We Know and What We Need to Know. J. Exp. Bot. 2020, 71, 3287–3295. [Google Scholar] [CrossRef]
- Ma, L.; Li, G. Auxin-Dependent Cell Elongation During the Shade Avoidance Response. Front. Plant Sci. 2019, 10, 914. [Google Scholar] [CrossRef]
- Ruberti, I.; Sessa, G.; Ciolfi, A.; Possenti, M.; Carabelli, M.; Morelli, G. Plant Adaptation to Dynamically Changing Environment: The Shade Avoidance Response. Biotechnol. Adv. 2012, 30, 1047–1058. [Google Scholar] [CrossRef]
- Keuskamp, D.H.; Pollmann, S.; Voesenek, L.A.C.J.; Peeters, A.J.M.; Pierik, R. Auxin Transport through PIN-FORMED 3 (PIN3) Controls Shade Avoidance and Fitness during Competition. Proc. Natl. Acad. Sci. USA 2010, 107, 22740–22744. [Google Scholar] [CrossRef]
- Yao, Y.; He, R.J.; Xie, Q.L.; Zhao, X.H.; Deng, X.M.; He, J.B.; Song, L.; He, J.; Marchant, A.; Chen, X.Y.; et al. ETHYLENE RESPONSE FACTOR 74 (ERF74) Plays an Essential Role in Controlling a Respiratory Burst Oxidase Homolog D (RbohD)-dependent Mechanism in Response to Different Stresses in Arabidopsis. New Phytol. 2017, 213, 1667–1681. [Google Scholar] [CrossRef]
- Eberhard, S.; Finazzi, G.; Wollman, F.A. The Dynamics of Photosynthesis. Annu. Rev. Genet. 2008, 42, 463–515. [Google Scholar] [CrossRef]
- Briat, J.-F.; Dubos, C.; Gaymard, F. Iron Nutrition, Biomass Production, and Plant Product Quality. Trends Plant Sci. 2015, 20, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Abadía, J.; Vázquez, S.; Rellán-Álvarez, R.; El-Jendoubi, H.; Abadía, A.; Álvarez-Fernández, A.; López-Millán, A.F. Towards a Knowledge-Based Correction of Iron Chlorosis. Plant Physiol. Biochem. 2011, 49, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Briat, J.F.; Duc, C.; Ravet, K.; Gaymard, F. Ferritins and Iron Storage in Plants. Biochim. Biophys. Acta 2010, 1800, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Ravet, K.; Touraine, B.; Boucherez, J.; Briat, J.F.; Gaymard, F.; Cellier, F. Ferritins Control Interaction between Iron Homeostasis and Oxidative Stress in Arabidopsis. Plant J. 2009, 57, 400–412. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.Q.; Bauer, P.; Bereczky, Z.; Keller, B.; Ganal, M. The Tomato Fer Gene Encoding a bHLH Protein Controls Iron-Uptake Responses in Roots. Proc. Natl. Acad. Sci. USA 2002, 99, 13938–13943. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.W.; Liu, M.J.; Chen, I.C.; Huang, C.H.; Chao, L.Y.; Hsieh, H.L. A Glutathione S -Transferase Regulated by Light and Hormones Participates in the Modulation of Arabidopsis Seedling Development. Plant Physiol. 2010, 154, 1646–1658. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.C.; Huang, I.C.; Liu, M.J.; Wang, Z.G.; Chung, S.S.; Hsieh, H.L. Glutathione S -Transferase Interacting with Far-Red Insensitive 219 Is Involved in Phytochrome A-Mediated Signaling in Arabidopsis. Plant Physiol. 2007, 143, 1189–1202. [Google Scholar] [CrossRef]
- Hsieh, H.L.; Okamoto, H.; Wang, M.; Ang, L.H.; Matsui, M.; Goodman, H.; Deng, X.W. FIN219, an Auxin-Regulated Gene, Defines a Link between Phytochrome A and the Downstream Regulator COP1 in Light Control of Arabidopsis Development. Genes Dev. 2000, 14, 1958–1970. [Google Scholar] [CrossRef]
- Franklin, K.A. PRR Proteins of the Circadian Clock Call Time on Shade Avoidance. Proc. Natl. Acad. Sci. USA 2020, 117, 5095–5096. [Google Scholar] [CrossRef]
- Zhang, Y.; Pfeiffer, A.; Tepperman, J.M.; Dalton-Roesler, J.; Leivar, P.; Gonzalez Grandio, E.; Quail, P.H. Central Clock Components Modulate Plant Shade Avoidance by Directly Repressing Transcriptional Activation Activity of PIF Proteins. Proc. Natl. Acad. Sci. USA 2020, 117, 3261–3269. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Meth. Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-Based Genome Alignment and Genotyping with HISAT2 and HISAT-Genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Colle, M.; Leisner, C.P.; Wai, C.M.; Ou, S.; Bird, K.A.; Wang, J.; Wisecaver, J.H.; Yocca, A.E.; Alger, E.I.; Tang, H.; et al. Haplotype-Phased Genome and Evolution of Phytonutrient Pathways of Tetraploid Blueberry. GigaScience 2019, 8, giz012. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. WGCNA: An R Package for Weighted Correlation Network Analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Forslund, K.; Coelho, L.P.; Szklarczyk, D.; Jensen, L.J.; Von Mering, C.; Bork, P. Fast Genome-Wide Functional Annotation through Orthology Assignment by eggNOG-Mapper. Mol. Biol. Evol. 2017, 34, 2115–2122. [Google Scholar] [CrossRef]
- Chen, C.; Wu, Y.; Li, J.; Wang, X.; Zeng, Z.; Xu, J.; Liu, Y.; Feng, J.; Chen, H.; He, Y.; et al. TBtools-II: A ‘One for All, All for One‘ Bioinformatics Platform for Biological Big-Data Mining. Mol. Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO Summarizes and Visualizes Long Lists of Gene Ontology Terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef]
- Jiang, Z.; Wang, Y.; Li, W.; Wang, Y.; Liu, X.; Ou, X.; Su, W.; Song, S.; Chen, R. Genome-Wide Identification of the NF-Y Gene Family and Their Involvement in Bolting and Flowering in Flowering Chinese Cabbage. Int. J. Mol. Sci. 2023, 24, 11898. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Jiang, Z.; Li, W.; Yang, X.; Li, C.; Cai, D.; Pan, Y.; Su, W.; Chen, R. Supplementary Low Far-Red Light Promotes Proliferation and Photosynthetic Capacity of Blueberry In Vitro Plantlets. Int. J. Mol. Sci. 2024, 25, 688. https://doi.org/10.3390/ijms25020688
Wang Y, Jiang Z, Li W, Yang X, Li C, Cai D, Pan Y, Su W, Chen R. Supplementary Low Far-Red Light Promotes Proliferation and Photosynthetic Capacity of Blueberry In Vitro Plantlets. International Journal of Molecular Sciences. 2024; 25(2):688. https://doi.org/10.3390/ijms25020688
Chicago/Turabian StyleWang, Yuting, Zhehao Jiang, Wenxiang Li, Xiaolong Yang, Chengming Li, Dunxu Cai, Yunxue Pan, Wei Su, and Riyuan Chen. 2024. "Supplementary Low Far-Red Light Promotes Proliferation and Photosynthetic Capacity of Blueberry In Vitro Plantlets" International Journal of Molecular Sciences 25, no. 2: 688. https://doi.org/10.3390/ijms25020688