
Design, Synthesis and Gene Modulation Insights into Pigments Derived from Tryptophan-Betaxanthin, Which Act against Tumor Development in Caenorhabditis elegans

 , , and
, , and
Abstract
:1. Introduction
2. Results
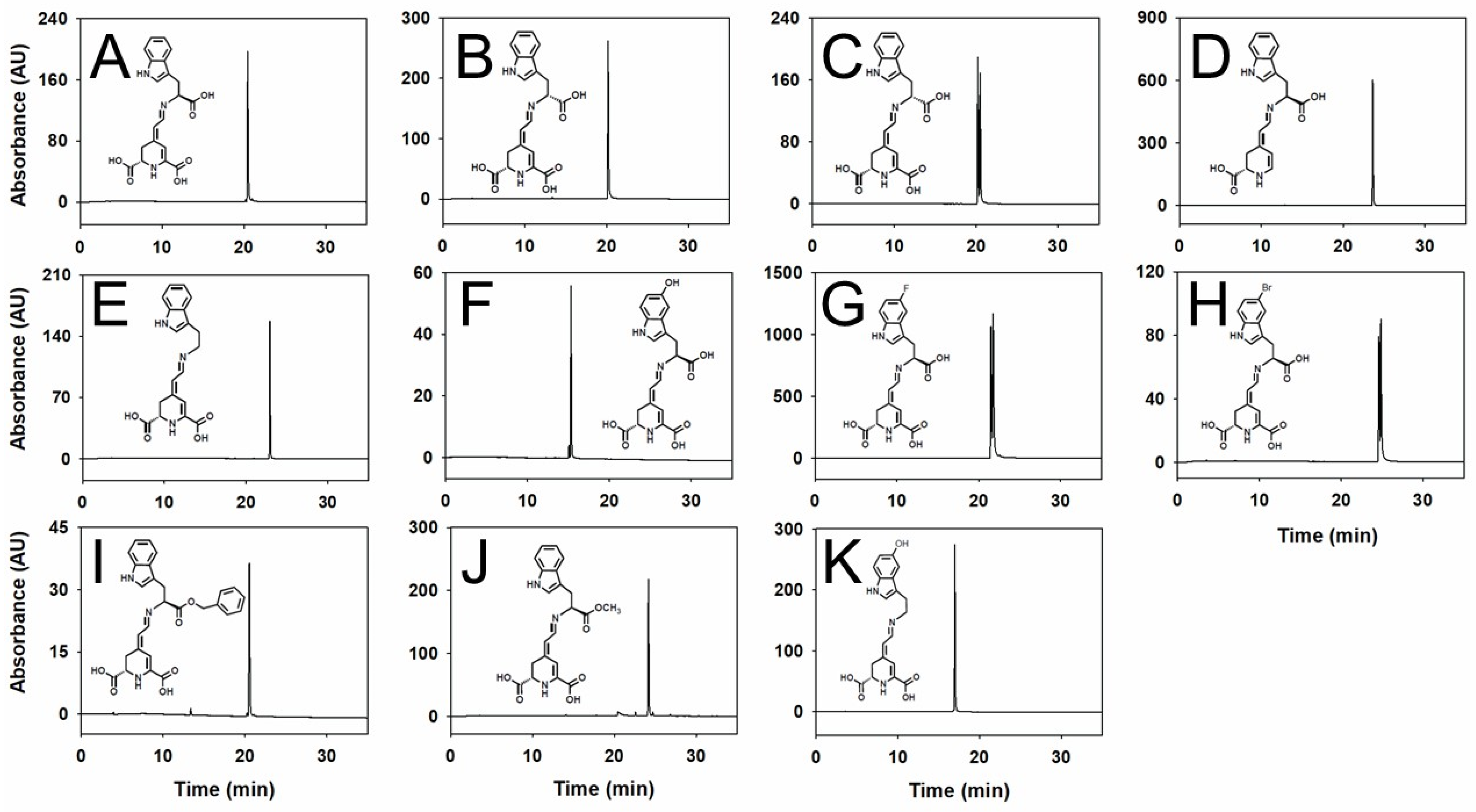
2.1. Production and Purification of Tryptophan-Derived Betaxanthins
2.2. Color of Tryptophan-Derived Betaxanthins
2.3. Effect of Tryptophan-Derived Betaxanthins on Tumor Growth in C. elegans
2.4. Microarray Analysis
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Production of Betalains by Cellular Factories
4.3. Purification of Betalains
4.4. Absorbance Spectroscopy
4.5. HPLC Analysis
4.6. Analysis by Mass Spectrometry with Electrospray Ionization
4.7. C. elegans Culture Conditions and Strains
4.8. Antitumoral Effect of Tryptophan-Derived Betaxanthins In Vivo
4.9. Tumor Size Evaluation
4.10. RNA Extraction and Microarray Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization International Agency for Research on Cancer The Global Cancer Observatory—All cancers. Int. Agency Res. Cancer—WHO 2020, 419, 199–200.
- Hassanpour, S.H.; Dehghani, M. Review of cancer from perspective of molecular. J. Cancer Res. Pract. 2017, 4, 127–129. [Google Scholar] [CrossRef]
- Hoenerhoff, M.J.; Boyle, M.; Minocherhomji, S.; Pandiri, A.R. Carcinogenesis: Mechanisms and Evaluation. In Haschek and Rousseaux’s Handbook of Toxicologic Pathology Volume 1: Principles and Practice of Toxicologic Pathology; BioMed Research International; Academic Press: Cambridge, MA, USA, 2022; pp. 205–254. [Google Scholar] [CrossRef]
- Fisher, R.; Pusztai, L.; Swanton, C. Cancer heterogeneity: Implications for targeted therapeutics. Br. J. Cancer 2013, 108, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Yang, Y.; Guo, T.; Veeraraghavan, V.P.; Wang, X. Potential chemotherapeutic effect of betalain against human non-small cell lung cancer through PI3K/Akt/mTOR signaling pathway. Environ. Toxicol. 2021, 36, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Allegra, M.; De Cicco, P.; Ercolano, G.; Attanzio, A.; Busà, R.; Cirino, G.; Tesoriere, L.; Livrea, M.A.; Ianaro, A. Indicaxanthin from Opuntia Ficus Indica (L. Mill) impairs melanoma cell proliferation, invasiveness, and tumor progression. Phytomedicine 2018, 50, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Rodríguez, P.; Guerrero-Rubio, M.A.; Henarejos-Escudero, P.; García-Carmona, F.; Gandía-Herrero, F. Health-promoting potential of betalains in vivo and their relevance as functional ingredients: A review. Trends Food Sci. Technol. 2022, 122, 66–82. [Google Scholar] [CrossRef]
- Da Silva, D.V.T.; Pereira, A.D.; Boaventura, G.T.; Ribeiro, R.S.D.A.; Verícimo, M.A.; De Carvalho-Pinto, C.E.; Baião, D.D.S.; Del Aguila, E.M.; Paschoalin, V.M.F. Short-term betanin intake reduces oxidative stress in wistar rats. Nutrients 2019, 11, 1978. [Google Scholar] [CrossRef]
- Han, J.; Ma, D.; Zhang, M.; Yang, X.; Tan, D. Natural antioxidant betanin protects rats from paraquat-induced acute lung injury interstitial pneumonia. Biomed Res. Int. 2015, 2015, 1–9. [Google Scholar] [CrossRef]
- Han, J.; Tan, C.; Wang, Y.; Yang, S.; Tan, D. Betanin reduces the accumulation and cross-links of collagen in high-fructose-fed rat heart through inhibiting non-enzymatic glycation. Chem. Biol. Interact. 2015, 227, 37–44. [Google Scholar] [CrossRef]
- Vulić, J.J.; Ćebović, T.N.; Čanadanović, V.M.; Ćetković, G.S.; Djilas, S.M.; Čanadanović-Brunet, J.M.; Velićanski, A.S.; Cvetković, D.D.; Tumbas, V.T. Antiradical, antimicrobial and cytotoxic activities of commercial beetroot pomace. Food Funct. 2013, 4, 713–721. [Google Scholar] [CrossRef]
- Clifford, T.; Allerton, D.M.; Brown, M.A.; Harper, L.; Horsburgh, S.; Keane, K.M.; Stevenson, E.J.; Howatson, G. Minimal muscle damage after a marathon and no influence of beetroot juice on inflammation and recovery. Appl. Physiol. Nutr. Metab. 2017, 42, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Macias-Ceja, D.C.; Cosín-Roger, J.; Ortiz-Masiá, D.; Salvador, P.; Hernández, C.; Esplugues, J.V.; Calatayud, S.; Barrachina, M.D. Stimulation of autophagy prevents intestinal mucosal inflammation and ameliorates murine colitis. Br. J. Pharmacol. 2017, 174, 2501–2511. [Google Scholar] [CrossRef] [PubMed]
- Martinez, R.M.; Longhi-Balbinot, D.T.; Zarpelon, A.C.; Staurengo-Ferrari, L.; Baracat, M.M.; Georgetti, S.R.; Sassonia, R.C.; Verri, W.A.; Casagrande, R. Anti-inflammatory activity of betalain-rich dye of Beta vulgaris: Effect on edema, leukocyte recruitment, superoxide anion and cytokine production. Arch. Pharm. Res. 2015, 38, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.; Wang, Y.; Bai, B.; Yang, X.; Han, J. Betanin attenuates oxidative stress and inflammatory reaction in kidney of paraquat-treated rat. Food Chem. Toxicol. 2015, 78, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Schliemann, W.; Cai, Y.; Degenkolb, T.; Schmidt, J.; Corke, H. Betalains of Celosia argentea. Phytochemistry 2001, 58, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Kugler, F.; Stintzing, F.C.; Carle, R. Characterisation of betalain patterns of differently coloured inflorescences from Gomphrena globosa L. and Bougainvillea sp. by HPLC–DAD–ESI–MS n. Anal. Bioanal. Chem. 2007, 387, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Kugler, F.; Stintzing, F.C.; Carle, R. Identification of betalains from petioles of differently colored Swiss chard (Beta vulgaris L. ssp. cicla [L.] Alef. Cv. Bright Lights) by high-performance liquid chromatography-electrospray ionization mass spectrometry. J. Agric. Food Chem. 2004, 52, 2975–2981. [Google Scholar] [CrossRef] [PubMed]
- García-Cayuela, T.; Gómez-Maqueo, A.; Guajardo-Flores, D.; Welti-Chanes, J.; Cano, M.P. Characterization and quantification of individual betalain and phenolic compounds in Mexican and Spanish prickly pear (Opuntia ficus-indica L. Mill) tissues: A comparative study. J. Food Compos. Anal. 2019, 76, 1–13. [Google Scholar] [CrossRef]
- Betancourt, C.; Cejudo-Bastante, M.J.; Heredia, F.J.; Hurtado, N. Pigment composition and antioxidant capacity of betacyanins and betaxanthins fractions of Opuntia dillenii (Ker Gawl) Haw cactus fruit. Food Res. Int. 2017, 101, 173–179. [Google Scholar] [CrossRef]
- Sawicki, T.; Bączek, N.; Wiczkowski, W. Betalain profile, content and antioxidant capacity of red beetroot dependent on the genotype and root part. J. Funct. Foods 2016, 27, 249–261. [Google Scholar] [CrossRef]
- Chen, K.C.; Jian, Y.R.; Sun, M.F.; Chang, T.T.; Lee, C.C.; Chen, C.Y.C. Investigation of silent information regulator 1 (Sirt1) agonists from Traditional Chinese Medicine. J. Biomol. Struct. Dyn. 2013, 31, 1207–1218. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.-C.; Wang, J.-D.; Lee, M.-M.; Chang, S.-S.; Tsai, T.-Y.; Chang, K.-W.; Tsai, F.-J.; Chen, C.Y.-C. Lose Weight with Traditional Chinese Medicine? Potential Suppression of Fat Mass and Obesity-Associated Protein. J. Biomol. Struct. Dyn. 2011, 29, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Henarejos-Escudero, P.; Hernández-García, S.; Guerrero-Rubio, M.A.; García-Carmona, F.; Gandía-Herrero, F. Antitumoral Drug Potential of Tryptophan-Betaxanthin and Related Plant Betalains in the Caenorhabditis elegans Tumoral Model. Antioxidants 2020, 9, 646. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.; Zarowiecki, M.; Arnaboldi, V.; Becerra, A.; Cain, S.; Chan, J.; Chen, W.J.; Cho, J.; da Veiga Beltrame, E.; Diamantakis, S.; et al. WormBase in 2022—Data, processes, and tools for analyzing Caenorhabditis elegans. Genetics 2022, 220, iyac003. [Google Scholar] [CrossRef] [PubMed]
- Shaye, D.D.; Greenwald, I. OrthoList: A Compendium of C. elegans Genes with Human Orthologs. PLoS ONE 2011, 6, e20085. [Google Scholar] [CrossRef] [PubMed]
- Medina, P.M.; Ponce, J.M.; Cruz, C.A. Revealing the anticancer potential of candidate drugs in vivo using Caenorhabditis elegans mutant strains. Transl. Oncol. 2021, 14, 100940. [Google Scholar] [CrossRef]
- Kirienko, N.V.; Mani, K.; Fay, D.S. Cancer models in Caenorhabditis elegans. Dev. Dyn. 2010, 239, 1413–1448. [Google Scholar] [CrossRef]
- Jhanwar-Uniyal, M.; Wainwright, J.V.; Mohan, A.L.; Tobias, M.E.; Murali, R.; Gandhi, C.D.; Schmidt, M.H. Diverse signaling mechanisms of mTOR complexes: mTORC1 and mTORC2 in forming a formidable relationship. Adv. Biol. Regul. 2019, 72, 51–62. [Google Scholar] [CrossRef]
- Hou, G.; Zhao, Q.; Zhang, M.; Fan, T.; Liu, M.; Shi, X.; Ren, Y.; Wang, Y.; Zhou, J.; Lu, Z. Down-regulation of Rictor enhances cell sensitivity to PI3K inhibitor LY294002 by blocking mTORC2-medicated phosphorylation of Akt/PRAS40 in esophageal squamous cell carcinoma. Biomed. Pharmacother. 2018, 106, 1348–1356. [Google Scholar] [CrossRef]
- Shi, D.; Zhu, Y.; Niu, P.; Zhou, J.; Chen, H. Raptor mediates the antiproliferation of cardamonin by mTORC1 inhibition in SKOV3 cells. OncoTargets Ther. 2018, 11, 757–767. [Google Scholar] [CrossRef]
- Devarajan, E.; Sahin, A.A.; Chen, J.S.; Krishnamurthy, R.R.; Aggarwal, N.; Brun, A.M.; Sapino, A.; Zhang, F.; Sharma, D.; Yang, X.H.; et al. Down-regulation of caspase 3 in breast cancer: A possible mechanism for chemoresistance. Oncogene 2002, 21, 8843–8851. [Google Scholar] [CrossRef] [PubMed]
- Winter, R.N.; Kramer, A.; Borkowski, A.; Kyprianou, N. Loss of caspase-1 and caspase-3 protein expression in human prostate cancer. Cancer Res. 2001, 61, 1227–1232. [Google Scholar]
- Henarejos-Escudero, P.; Hernández-García, S.; Martínez-Rodríguez, P.; García-Carmona, F.; Gandía-Herrero, F. Bioactive potential and spectroscopical characterization of a novel family of plant pigments betalains derived from dopamine. Food Res. Int. 2022, 162, 111956. [Google Scholar] [CrossRef] [PubMed]
- Francis, R.; Barton, M.K.; Kimble, J.; Schedl, T. gld-1, a tumor suppressor gene required for oocyte development in Caenorhabditis elegans. Genetics 1995, 139, 579–606. [Google Scholar] [CrossRef]
- Chen, K.C.; Chang, S.S.; Huang, H.J.; Lin, T.L.; Wu, Y.J.; Chen, C.Y.C. Three-in-one agonists for PPAR-a, PPAR-γ, and PPAR-d from traditional Chinese medicine. J. Biomol. Struct. Dyn. 2012, 30, 662–683. [Google Scholar] [CrossRef]
- Ortega, E.; Ballester, F.J.; Hernández-García, A.; Hernández-García, S.; Guerrero-Rubio, M.A.; Bautista, D.; Santana, M.D.; Gandía-Herrero, F.; Ruiz, J. Novel organo-osmium(ii) proteosynthesis inhibitors active against human ovarian cancer cells reduce gonad tumor growth in: Caenorhabditis elegans. Inorg. Chem. Front. 2021, 8, 141–155. [Google Scholar] [CrossRef]
- Ortega-Forte, E.; Hernández-García, S.; Vigueras, G.; Henarejos-Escudero, P.; Cutillas, N.; Ruiz, J.; Gandía-Herrero, F. Potent anticancer activity of a novel iridium metallodrug via oncosis. Cell. Mol. Life Sci. 2022, 79. [Google Scholar] [CrossRef] [PubMed]
- Bristow, A.W.T.; Webb, K.S. Intercomparison study on accurate mass measurement of small molecules in mass spectrometry. J. Am. Soc. Mass Spectrom. 2003, 14, 1086–1098. [Google Scholar] [CrossRef]
- Stiernagle, T. Maintenance of C. elegans. WormBook 2006. [Google Scholar] [CrossRef]
- Ahringer, J. Reverse genetics. WormBook 2006. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Pigment | HPLC Analysis Rt (min) | Chemical Formula | TOF Exact Mass | ||
|---|---|---|---|---|---|
| Experimental (m/z) | Theoretical (m/z) | Δppm | |||
| L-tryptophan-betaxanthin | 20.47 | C20H19N3O6 | 398.1322 | 398.1347 | 6.2 |
| D-tryptophan-betaxanthin | 20.17 | C20H19N3O6 | 398.1323 | 398.1347 | 6.0 |
| L-tryptophan-6-decarboxy-betaxanthin | 23.63 | C19H19N3O4 | 354.1424 | 354.1448 | 6.7 |
| Tryptamine-betaxanthin | 22.95 | C19H19N3O4 | 354.1464 | 354.1448 | 4.5 |
| 5-hydroxy-L-tryptophan-betaxanthin | 15.31 | C20H19N3O7 | 414.1277 | 414.1296 | 4.5 |
| 5-fluoro-DL-tryptophan-betaxanthin | 21.53 21.78 | C20H18FN3O6 | 416.1228 416.1231 | 416.1252 416.1252 | 5.7 5.0 |
| 5-bromo-DL-tryptophan-betaxanthin | 24.63 24.88 | C20H18BrN3O6 | 476.0426 476.0429 | 476.0452 476.0452 | 5.4 4.8 |
| L-tryptophan-benzyl ester-betaxanthin | 20.53 | C27H25N3O6 | 488.1790 | 488.1816 | 5.3 |
| L-tryptophan-methyl ester-betaxanthin | 24.16 | C21H21N3O6 | 412.1500 | 412.1503 | 0.7 |
| Serotonin-betaxanthin | 16.97 | C19H19N3O6 | 370.1411 | 370.1397 | 3.7 |
| Pigment | Absorbance | ||
|---|---|---|---|
| λmax (nm) | ε (M−1 cm−1) | Width (nm) | |
| L-tryptophan-betaxanthin | 477 | 42,000 | 64 |
| D-tryptophan-betaxanthin | 477 | 42,000 | 65 |
| L-tryptophan-6-decarboxy-betaxanthin | 474 | 49,000 | 61 |
| Tryptamine-betaxanthin | 473 | 36,000 | 59 |
| 5-hydroxy-L-tryptophan-betaxanthin | 477 | 38,000 | 70 |
| 5-fluoro-DL-tryptophan-betaxanthin | 479 | 43,000 | 61 |
| 5-bromo-DL-tryptophan-betaxanthin | 479 | 55,000 | 62 |
| L-tryptophan-benzyl ester-betaxanthin | 475 | 45,000 | 68 |
| L-tryptophan-methyl ester-betaxanthin | 479 | 36,000 | 67 |
| Serotonin-betaxanthin | 474 | 43,000 | 68 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henarejos-Escudero, P.; Méndez-García, F.F.; Hernández-García, S.; Martínez-Rodríguez, P.; Gandía-Herrero, F. Design, Synthesis and Gene Modulation Insights into Pigments Derived from Tryptophan-Betaxanthin, Which Act against Tumor Development in Caenorhabditis elegans. Int. J. Mol. Sci. 2024, 25, 63. https://doi.org/10.3390/ijms25010063
Henarejos-Escudero P, Méndez-García FF, Hernández-García S, Martínez-Rodríguez P, Gandía-Herrero F. Design, Synthesis and Gene Modulation Insights into Pigments Derived from Tryptophan-Betaxanthin, Which Act against Tumor Development in Caenorhabditis elegans. International Journal of Molecular Sciences. 2024; 25(1):63. https://doi.org/10.3390/ijms25010063
Chicago/Turabian StyleHenarejos-Escudero, Paula, Fernando F. Méndez-García, Samanta Hernández-García, Pedro Martínez-Rodríguez, and Fernando Gandía-Herrero. 2024. "Design, Synthesis and Gene Modulation Insights into Pigments Derived from Tryptophan-Betaxanthin, Which Act against Tumor Development in Caenorhabditis elegans" International Journal of Molecular Sciences 25, no. 1: 63. https://doi.org/10.3390/ijms25010063