
PCR Array Profiling of miRNA Expression Involved in the Differentiation of Amniotic Fluid Stem Cells toward Endothelial and Smooth Muscle Progenitor Cells

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
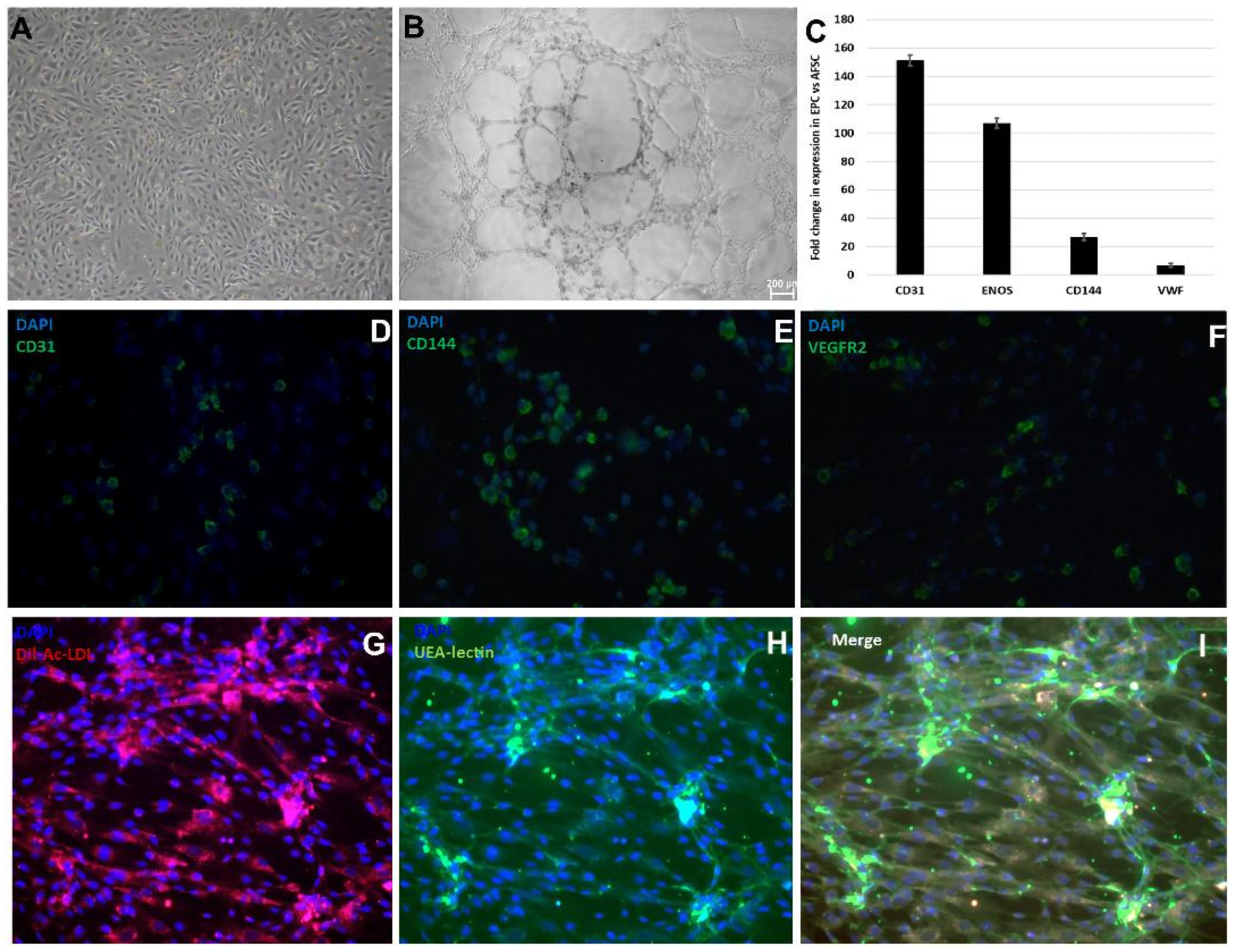
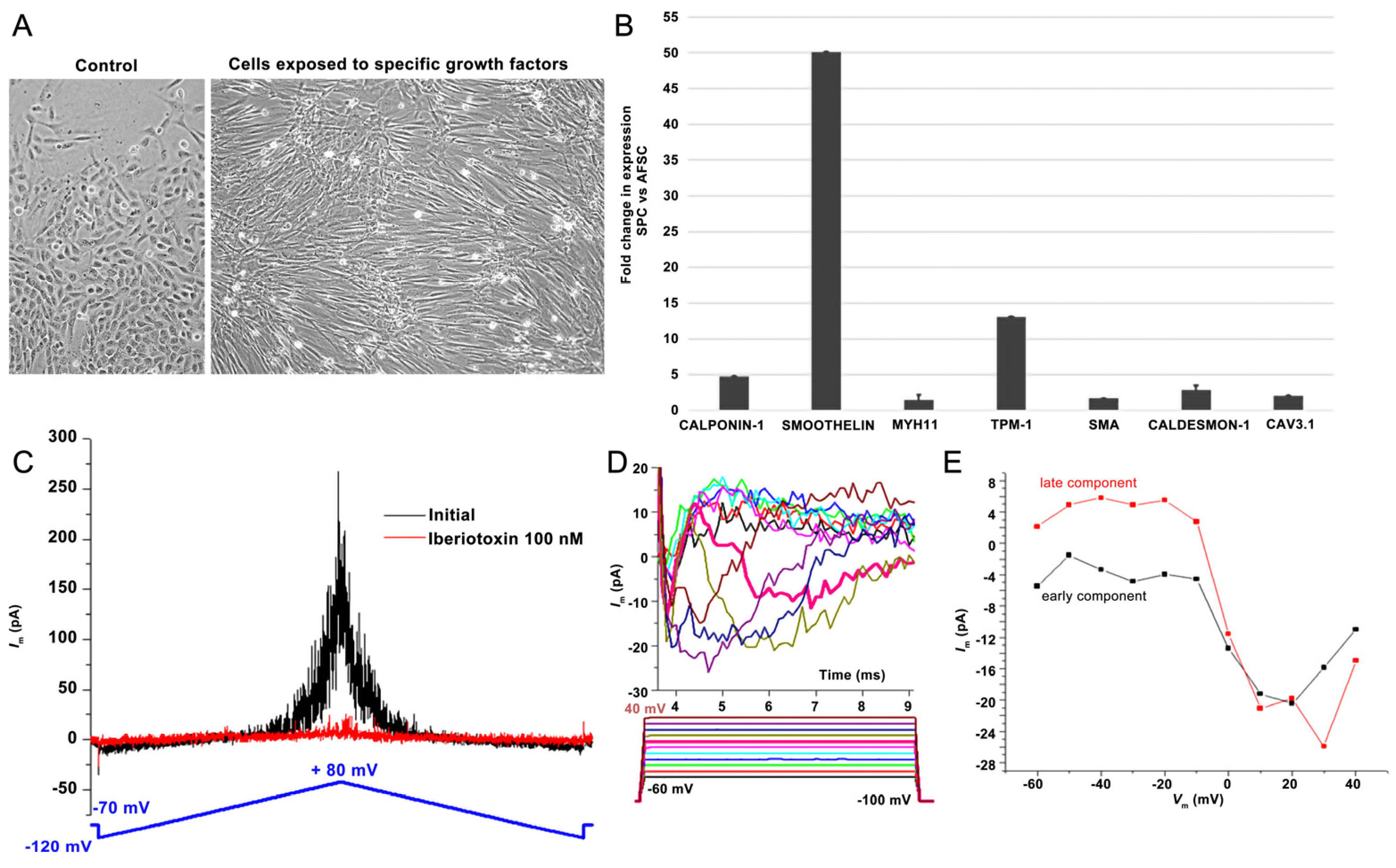
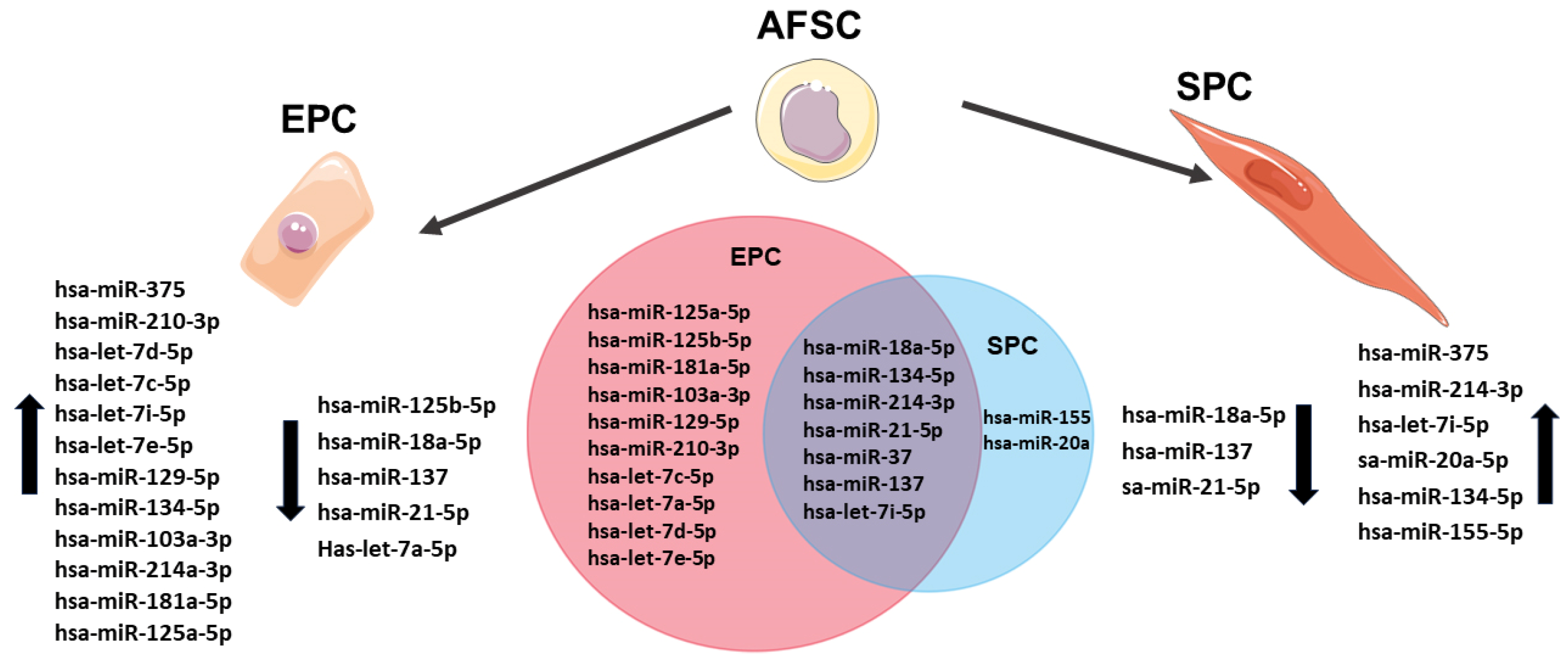
2. Results
2.1. AFSCs Differentiated to Endothelial Progenitor Cells (EPCs) and Smooth Muscle Progenitor Cells (SPCs)
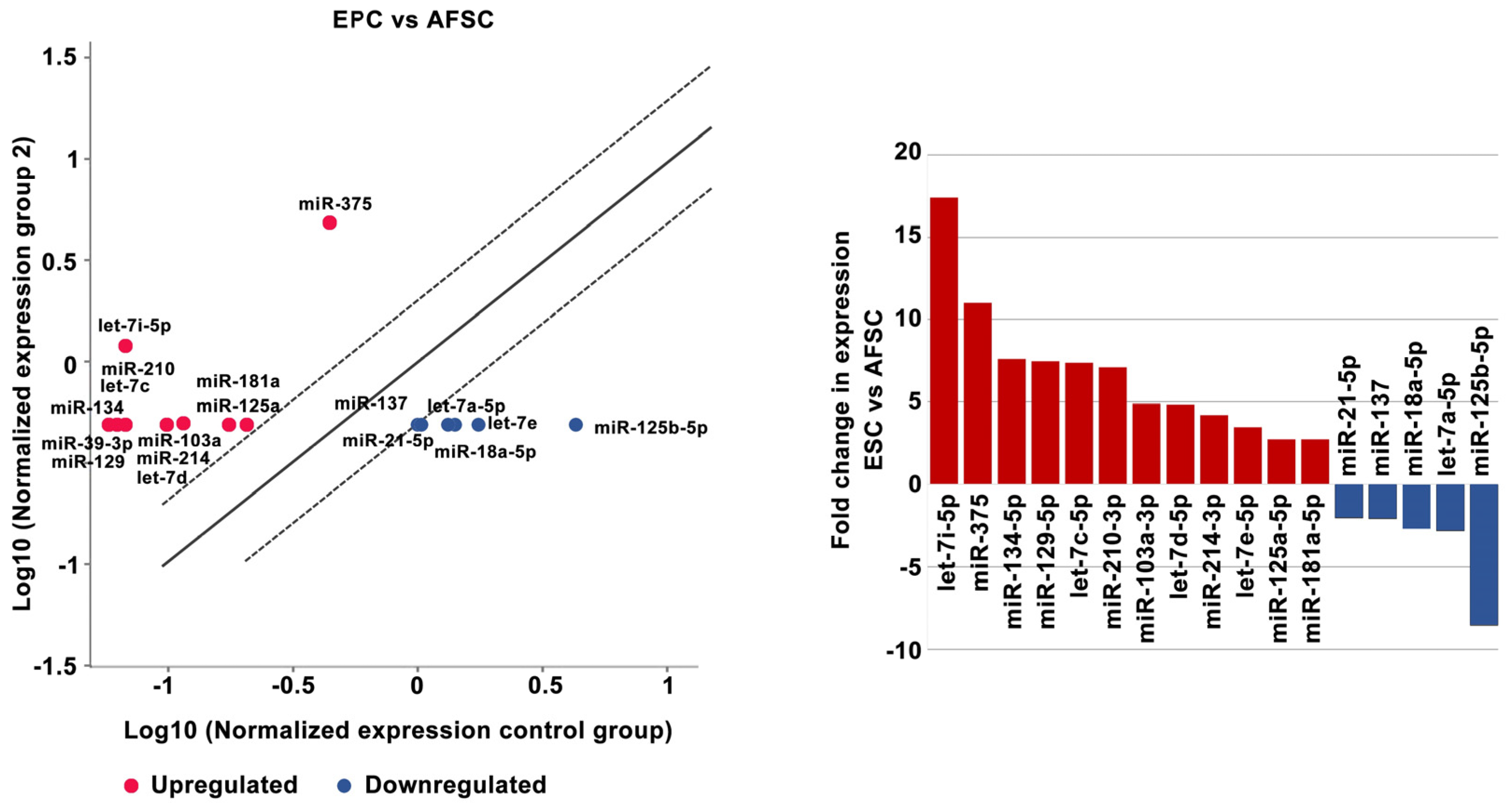
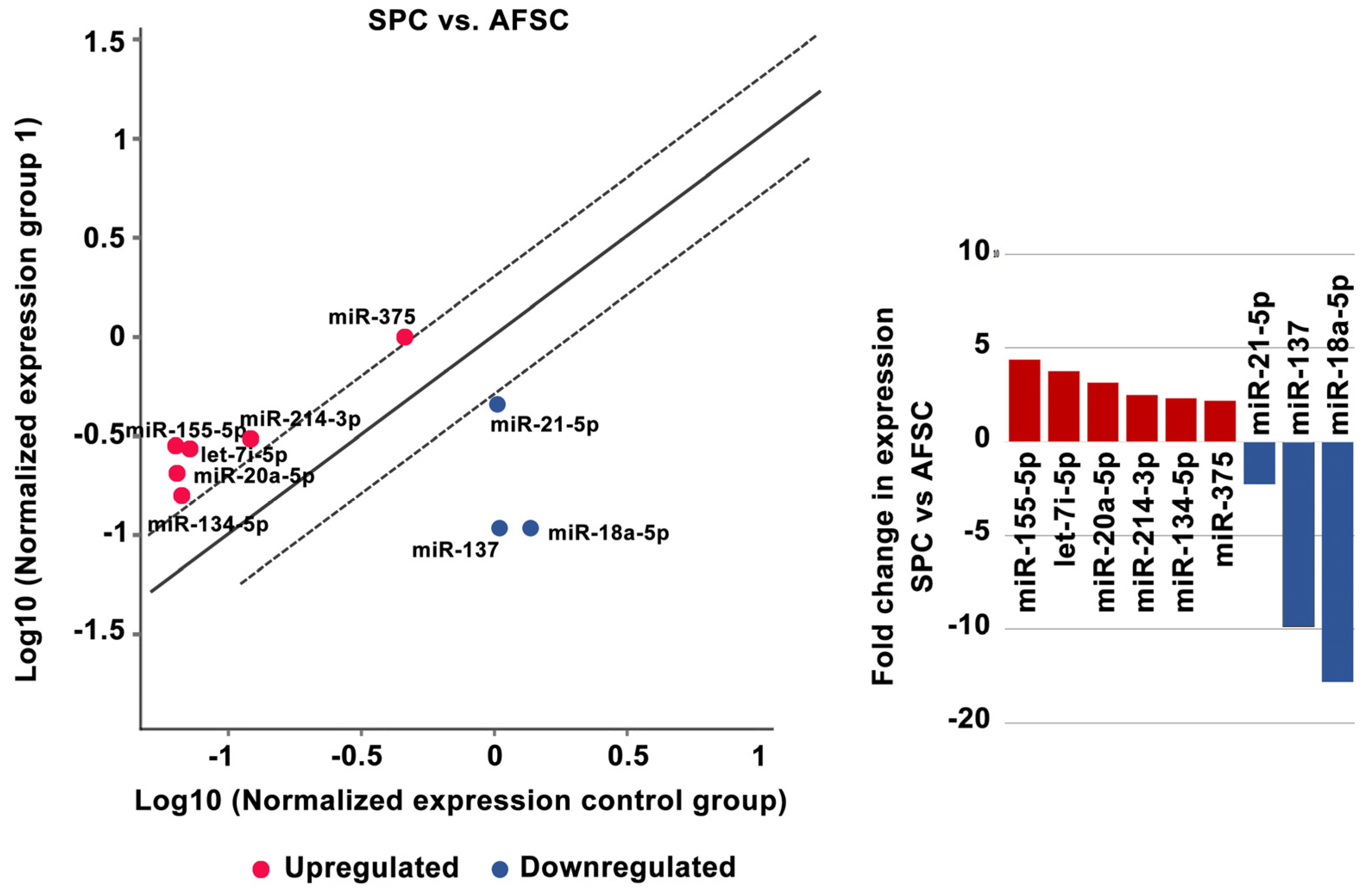
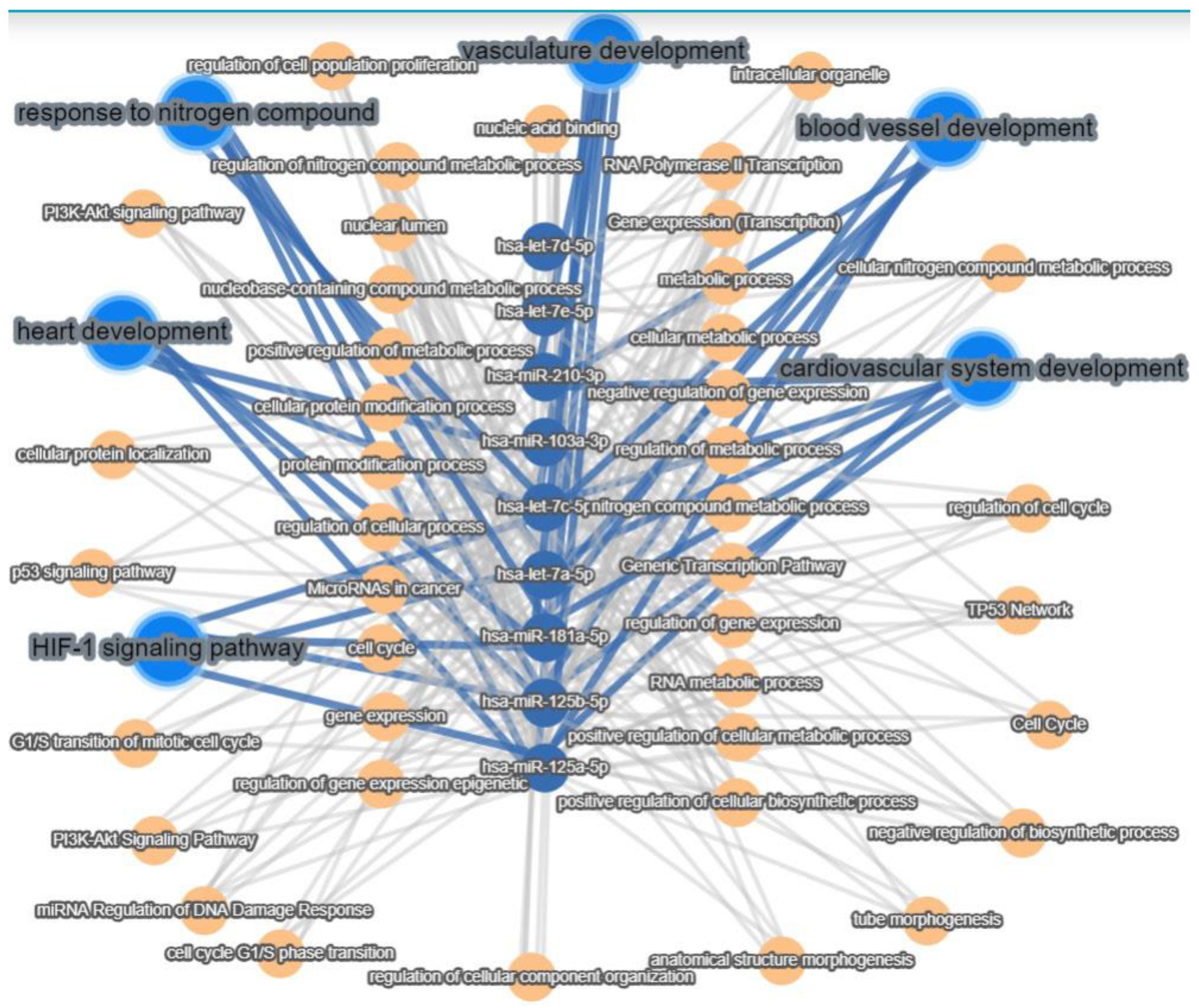
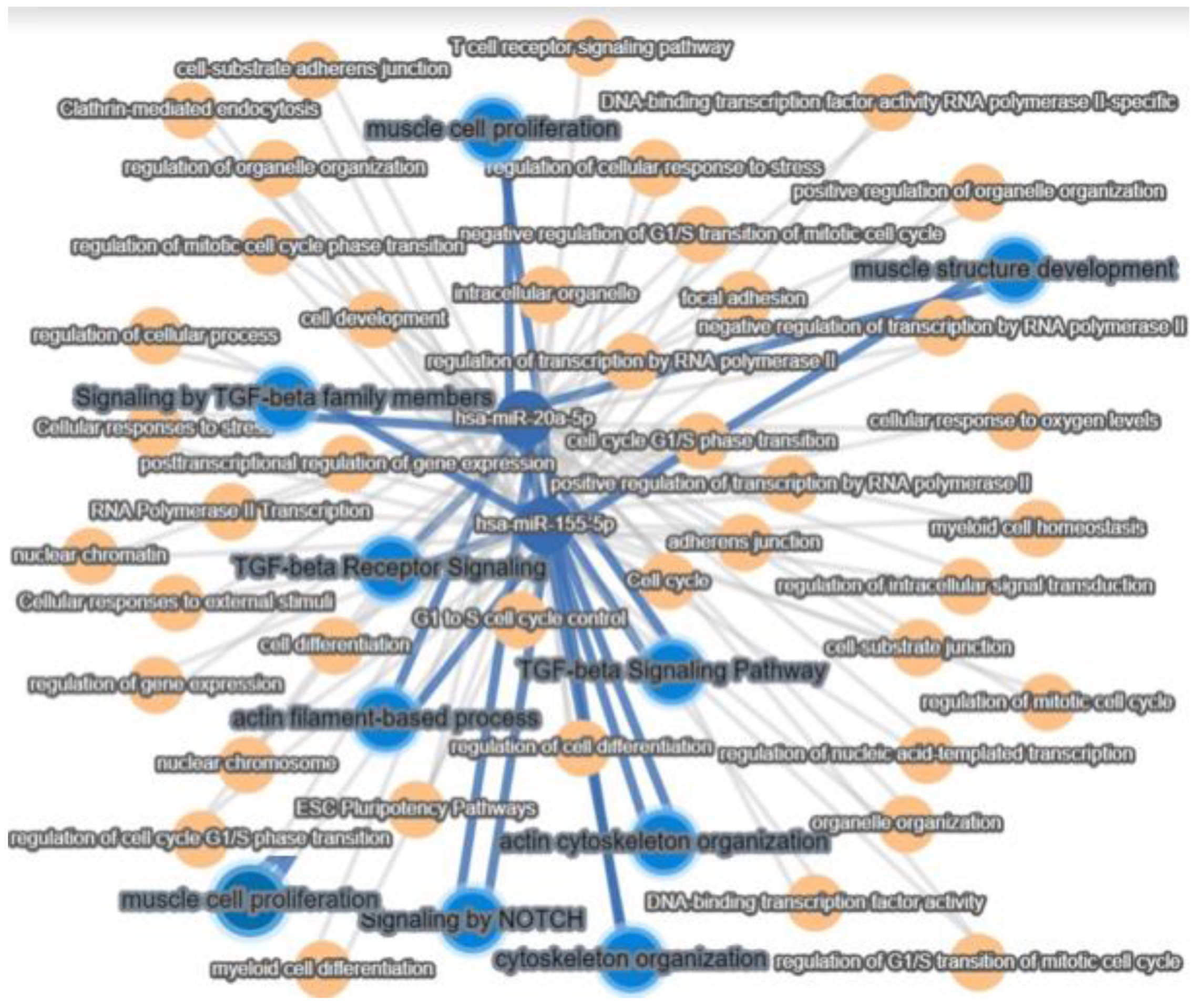
2.2. MiRNA Regulates the Differentiation of AFSCs
3. Discussion
4. Materials and Methods
4.1. Amniotic Fluid Stem Cells Differentiation and Characterization
4.2. Gene Expression and Functional Characterization of Endothelial Progenitor Cells (EPCs)
4.3. Gene Expression Assays on Smooth Muscle Progenitor Cells (SPCs)
4.4. Patch-Clamp Assays on Smooth Muscle Progenitor Cells (SPCs)
4.5. MiRNA Expression
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Loukogeorgakis, S.P.; De Coppi, P. Concise Review: Amniotic Fluid Stem Cells: The Known, the Unknown, and Potential Regenerative Medicine Applications. Stem Cells 2017, 35, 1663–1673. [Google Scholar] [CrossRef] [PubMed]
- Cananzi, M.; De Coppi, P. CD117+ amniotic fluid stem cells: State of the art and future perspectives. Organogenesis 2012, 8, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Resca, E.; Zavatti, M.; Maraldi, T.; Bertoni, L.; Beretti, F.; Guida, M.; La Sala, G.B.; Guillot, P.V.; David, A.L.; Sebire, N.J.; et al. Enrichment in c-Kit improved differentiation potential of amniotic membrane progenitor/stem cells. Placenta 2015, 36, 18–26. [Google Scholar] [CrossRef] [PubMed]
- De Coppi, P.; Bartsch, G., Jr.; Siddiqui, M.M.; Xu, T.; Santos, C.C.; Perin, L.; Mostoslavsky, G.; Serre, A.C.; Snyder, E.Y.; Yoo, J.J.; et al. Isolation of amniotic stem cell lines with potential for therapy. Nat. Biotechnol. 2007, 25, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Shamsnajafabadi, H.; Soheili, Z.S. Amniotic fluid characteristics and its application in stem cell therapy: A review. Int. J. Reprod. Biomed. 2022, 20, 627–643. [Google Scholar] [CrossRef]
- Cananzi, M.; Atala, A.; De Coppi, P. Stem cells derived from amniotic fluid: New potentials in regenerative medicine. Reprod. Biomed. Online 2009, 18, 17–27. [Google Scholar] [CrossRef]
- Di Baldassarre, A.; D’Amico, M.A.; Izzicupo, P.; Gaggi, G.; Guarnieri, S.; Mariggiò, M.A.; Antonucci, I.; Corneo, B.; Sirabella, D.; Stuppia, L.; et al. Cardiomyocytes derived from human cardiopoietic amniotic fluids. Sci. Rep. 2018, 8, 12028. [Google Scholar] [CrossRef]
- Di Tizio, D.; Di Serafino, A.; Upadhyaya, P.; Sorino, L.; Stuppia, L.; Antonucci, I. The impact of epigenetic signatures on amniotic fluid stem cell fate. Stem Cells Int. 2018, 2018, 4274518. [Google Scholar] [CrossRef]
- Condorelli, G.; Latronico, M.V.; Cavarretta, E. microRNAs in cardiovascular diseases: Current knowledge and the road ahead. J. Am. Coll. Cardiol. 2014, 63, 2177–2187. [Google Scholar] [CrossRef]
- Milcu, A.I.; Anghel, A.; Muşat, O.; Munteanu, M.; Salavat, M.C.; Iordache, A.; Ungureanu, E.; Bonţe, D.C.; Borugă, O. Implications at the ocular level of miRNAs modifications induced by SARS-CoV-2 infection. Rom. J. Morphol. Embryol. 2022, 63, 55–59. [Google Scholar] [CrossRef]
- Trohatou, O.; Zagoura, D.; Bitsika, V.; Pappa, K.I.; Antsaklis, A.; Anagnou, N.P.; Roubelakis, M.G. Sox2 suppression by miR-21 governs human mesenchymal stem cell properties. Stem Cells Transl. Med. 2014, 3, 54–68. [Google Scholar] [CrossRef]
- Lazzarini, R.; Olivieri, F.; Ferretti, C.; Mattioli-Belmonte, M.; di Primio, R.; Orciani, M. mRNAs and miRNAs profiling of mesenchymal stem cells derived from amniotic fluid and skin: The double face of the coin. Cell Tissue Res. 2014, 355, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Gholizadeh-Ghaleh Aziz, S.; Pashaei-Asl, F.; Fardyazar, Z.; Pashaiasl, M. Isolation, characterization, cryopreservation of human amniotic stem cells and differentiation to osteogenic and adipogenic cells. PLoS ONE 2016, 11, e0158281. [Google Scholar] [CrossRef] [PubMed]
- Glemžaitė, M.; Navakauskienė, R. Osteogenic differentiation of human amniotic fluid mesenchymal stem cells is determined by epigenetic changes. Stem Cells Int. 2016, 2016, 6465307. [Google Scholar] [CrossRef]
- Lazzarini, R.; Sorgentoni, G.; Caffarini, M.; Sayeed, M.A.; Olivieri, F.; Di Primio, R.; Orciani, M. New miRNAs network in human mesenchymal stem cells derived from skin and amniotic fluid. Int. J. Immunopathol. Pharmacol. 2016, 29, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Miranda-Sayago, J.M.; Fernandez-Arcas, N.; Reyes-Engel, A.; Benito, C.; Narbona, I.; Alonso, A. Changes in CDKN 2 D, TP 53, and mi R 125a expression: Potential role in the evaluation of human amniotic fluid-derived mesenchymal stromal cell fitness. Genes Cells 2012, 17, 673–687. [Google Scholar] [CrossRef] [PubMed]
- Zentelytė, A.; Gasiūnienė, M.; Treigytė, G.; Baronaitė, S.; Savickienė, J.; Borutinskaitė, V.; Navakauskienė, R. Epigenetic regulation of amniotic fluid mesenchymal stem cell differentiation to the mesodermal lineages at normal and fetus-diseased gestation. J. Cell Biochem. 2020, 121, 1811–1822. [Google Scholar] [CrossRef]
- Gasiūnienė, M.; Zentelytė, A.; Treigytė, G.; Baronaitė, S.; Savickienė, J.; Utkus, A.; Navakauskienė, R. Epigenetic alterations in amniotic fluid mesenchymal stem cells derived from normal and fetus-affected gestations: A focus on myogenic and neural differentiations. Cell Biol. Int. 2019, 43, 299–312. [Google Scholar] [CrossRef]
- Miyamoto, T.; Furusawa, C.; Kaneko, K. Pluripotency, differentiation, and reprogramming: A gene expression dynamics model with epigenetic feedback regulation. PLoS Comput. Biol. 2015, 11, e1004476. [Google Scholar] [CrossRef]
- Bem, J.; Grabowska, I.; Daniszewski, M.; Zawada, D.; Czerwinska, A.M.; Bugajski, L.; Piwocka, K.; Fogtman, A.; Ciemerych, M.A. Transient microRNA expression enhances myogenic potential of mouse embryonic stem cells. Stem Cells 2018, 36, 655–670. [Google Scholar] [CrossRef]
- Iordache, F.; Buzila, C.A. Amniotic fetal stem cells—Derived endothelial progenitors, a support for personalized cardiovascular regenerative medicine. Arch. Balk. Med. Union 2022, 57, 8–16. [Google Scholar] [CrossRef]
- Che, P.; Liu, J.; Shan, Z.; Wu, R.; Yao, C.; Cui, J.; Zhu, X.; Wang, J.; Burnett, M.S.; Wang, S.; et al. miR-125a-5p impairs endothelial cell angiogenesis in aging mice via RTEF-1 downregulation. Aging Cell 2014, 13, 926–934. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Ge, J.; Wang, Z.; Ren, J.; Wang, X.; Xiong, H.; Gao, J.; Zhang, Y.; Zhang, Q. Let-7e modulates the inflammatory response in vascular endothelial cells through ceRNA crosstalk. Sci. Rep. 2017, 7, 42498. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xiao, X.; Zheng, J.; Li, M.; Yu, M.; Ping, F.; Wang, T.; Wang, X. Vildagliptin, a dipeptidyl peptidase-4 inhibitor, attenuated endothelial dysfunction through miRNAs in diabetic rats. Arch. Med. Sci. 2019, 17, 1378–1387. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhao, Q.; Gong, K.; Long, Y.; Zhang, J.; Li, Y.; Guo, X. Downregulation of miR-103a-3p contributes to endothelial progenitor cell dysfunction in deep vein thrombosis through PTEN targeting. Ann. Vasc. Surg. 2020, 64, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Yamakuchi, M. Endothelial senescence and microRNA. Biomol. Concepts 2012, 3, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Lei, Q.; Ma, J.; Ju, R. Effects of hypoxic-ischemic pre-treatment on microvesicles derived from endothelial progenitor cells. Exp. Ther. Med. 2020, 19, 2171–2178. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Flach, H.; Onizawa, M.; Wei, L.; McManusm, M.T.; Weiss, A. Negative regulation of Hif1a expression and TH17 differentiation by the hypoxia-regulated microRNA miR-210. Nat. Immunol. 2014, 15, 393–401. [Google Scholar] [CrossRef]
- Li, X.; Jia, Z.; Zhao, X.; Xu, M.; Chen, M. Expression of miR-210 in the peripheral blood of patients with newly diagnosed type 2 diabetes mellitus and its effect on the number and function of endothelial progenitor cells. Microvasc. Res. 2020, 131, 104032. [Google Scholar] [CrossRef]
- Shi, Z.H.; Han, X.Y.; Yao, M.D.; Liu, C.; Jiang, Q.; Yan, B. Differential MicroRNA expression pattern in endothelial Progenitor cells during diabetic retinopathy. Front. Cell Dev. Biol. 2021, 9, 773050. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, K.C.; Wu, W.; Subramaniam, S.; Shy, J.Y.; Chiu, J.J.; Li, J.Y.; Chien, S. MicroRNA-21 targets peroxisome proliferators-activated receptor-alpha in an autoregulatory loop to modulate flow-induced endothelial inflammation. Proc. Natl. Acad. Sci. USA 2011, 108, 10355–10360. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Yang, P.; Xiong, Q.; Song, X.; Yang, X.; Liu, L.; Yuan, W.; Rui, Y.C. MicroRNA-125a/b-5p inhibits endothelin-1 expression in vascular endothelial cells. J. Hypertens. 2010, 28, 1646–1654. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.C.; Hsieh, I.C.; Hsi, E.; Wang, Y.S.; Dai, C.Y.; Chou, W.W.; Juo, S.H. Negative feedback regulation between microRNA let-7g and the oxLDL receptor LOX-1. J. Cell Sci. 2011, 124, 4115–4124. [Google Scholar] [CrossRef] [PubMed]
- Bao, M.H.; Zhang, Y.W.; Lou, X.Y.; Cheng, Y.; Zhou, H.H. Protective effects of let-7a and let-7b on oxidized low-density lipoprotein induced endothelial cell injuries. PLoS ONE 2014, 9, e106540. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Li, K.; Huang, W.; Zhang, X. MiR-137 inhibited cell proliferation and migration of vascular smooth muscle cells via targeting IGFBP-5 and modulating the mTOR/STAT3 signaling. PLoS ONE 2017, 12, e0186245. [Google Scholar] [CrossRef]
- Kee, H.J.; Kim, G.R.; Cho, S.N.; Kwon, J.S.; Ahn, Y.; Kook, H.; Jeong, M.H. mir-18a-5p microrna increases vascular smooth muscle cell differentiation by downregulating syndecan4. Korean Circ. J. 2014, 44, 255–563. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Zhou, M.H.; Li, S.P.; Zhao, H.M.; Zhang, Y.R.; Chen, D.; Wu, Y.X.; Pang, Q.F. MiR-155-5p attenuates vascular smooth muscle cell oxidative stress and migration via inhibiting BACH1 expression. Biomedicines 2023, 11, 1679. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, H.; Xing, W.; Li, J.; Du, R.; Cao, D.; Wang, Y.; Yang, X.; Zhong, G.; Zhao, Y.; et al. Vascular smooth muscle cell-specific miRNA-214 knockout inhibits angiotensin II-induced hypertension through upregulation of Smad7. FASEB J. 2021, 35, e21947. [Google Scholar] [CrossRef]
- Kern, F.; Aparicio-Puerta, E.; Li, Y.; Fehlmann, T.; Kehl, T.; Wagner, V.; Ray, K.; Ludwig, N.; Lenhof, H.P.; Meese, E.; et al. miRTargetLink 2.0-interactive miRNA target gene and target pathway networks. Nucleic Acids Res. 2021, 49, W409–W416. [Google Scholar] [CrossRef]
- Iordache, F.; Constantinescu, A.; Andrei, E.; Amuzescu, B.; Halitzchi, F.; Savu, L.; Maniu, H. Electrophysiology, immunophenotype, and gene expression characterization of senescent and cryopreserved human amniotic fluid stem cells. J. Physiol. Sci. 2016, 66, 463–476. [Google Scholar] [CrossRef]
- Airini, R.; Iordache, F.; Alexandru, D.; Savu, L.; Epureanu, F.B.; Mihailescu, D.; Amuzescu, B.; Maniu, H. Senescence-induced immunophenotype, gene expression and electrophysiology changes in human amniocytes. J. Cell Mol. Med. 2019, 23, 7233–7245. [Google Scholar] [CrossRef] [PubMed]
- Mann, S.A.; Heide, J.; Knott, T.; Airini, R.; Epureanu, F.B.; Deftu, A.F.; Deftu, A.T.; Radu, B.M.; Amuzescu, B. Recording of multiple ion current components and action potentials in human induced pluripotent stem cell-derived cardiomyocytes via automated patch-clamp. J. Pharmacol. Toxicol. Methods 2019, 100, 106599. [Google Scholar] [CrossRef] [PubMed]
- Scheel, O.; Frech, S.; Amuzescu, B.; Eisfeld, J.; Lin, K.H.; Knott, T. Action Potential Characterization of Human Induced Pluripotent Stem Cell–Derived Cardiomyocytes Using Automated Patch-Clamp Technology. ASSAY Drug Dev. Technol. 2014, 12, 457–469. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iordache, F.; Petcu, A.; Pisoschi, A.M.; Stanca, L.; Geicu, O.I.; Bilteanu, L.; Curuțiu, C.; Amuzescu, B.; Serban, A.I. PCR Array Profiling of miRNA Expression Involved in the Differentiation of Amniotic Fluid Stem Cells toward Endothelial and Smooth Muscle Progenitor Cells. Int. J. Mol. Sci. 2024, 25, 302. https://doi.org/10.3390/ijms25010302
Iordache F, Petcu A, Pisoschi AM, Stanca L, Geicu OI, Bilteanu L, Curuțiu C, Amuzescu B, Serban AI. PCR Array Profiling of miRNA Expression Involved in the Differentiation of Amniotic Fluid Stem Cells toward Endothelial and Smooth Muscle Progenitor Cells. International Journal of Molecular Sciences. 2024; 25(1):302. https://doi.org/10.3390/ijms25010302
Chicago/Turabian StyleIordache, Florin, Adriana (Ionescu) Petcu, Aurelia Magdalena Pisoschi, Loredana Stanca, Ovidiu Ionut Geicu, Liviu Bilteanu, Carmen Curuțiu, Bogdan Amuzescu, and Andreea Iren Serban. 2024. "PCR Array Profiling of miRNA Expression Involved in the Differentiation of Amniotic Fluid Stem Cells toward Endothelial and Smooth Muscle Progenitor Cells" International Journal of Molecular Sciences 25, no. 1: 302. https://doi.org/10.3390/ijms25010302