
Activity of Colocasia esculenta (Taro) Corms against Gastric Adenocarcinoma Cells: Chemical Study and Molecular Characterization
 , , ,
, , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Production of the Methanolic Extract of Colocasia Corms and Leaves and Cytotoxicity in AGS Cells
2.2. Studies on Cytotoxicity of Colocasia Corm Fractions in AGS Cells
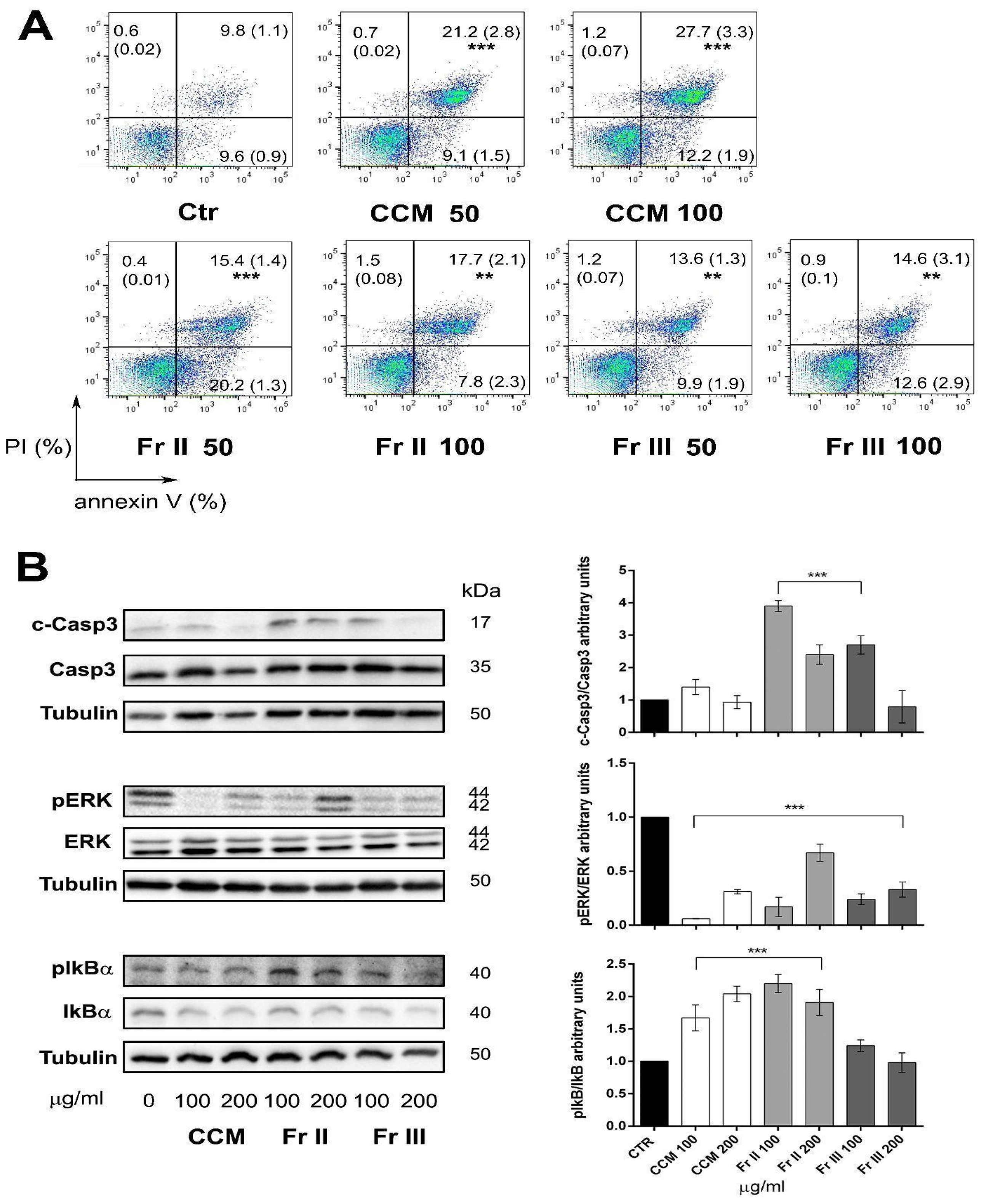
2.3. Colocasia Methanolic Extract and Fractions Induce Apoptosis in AGS Cells Modulating the Expression of Key Proteins Involved in Cell Proliferation, Cell Cycle, and Apoptosis Processes
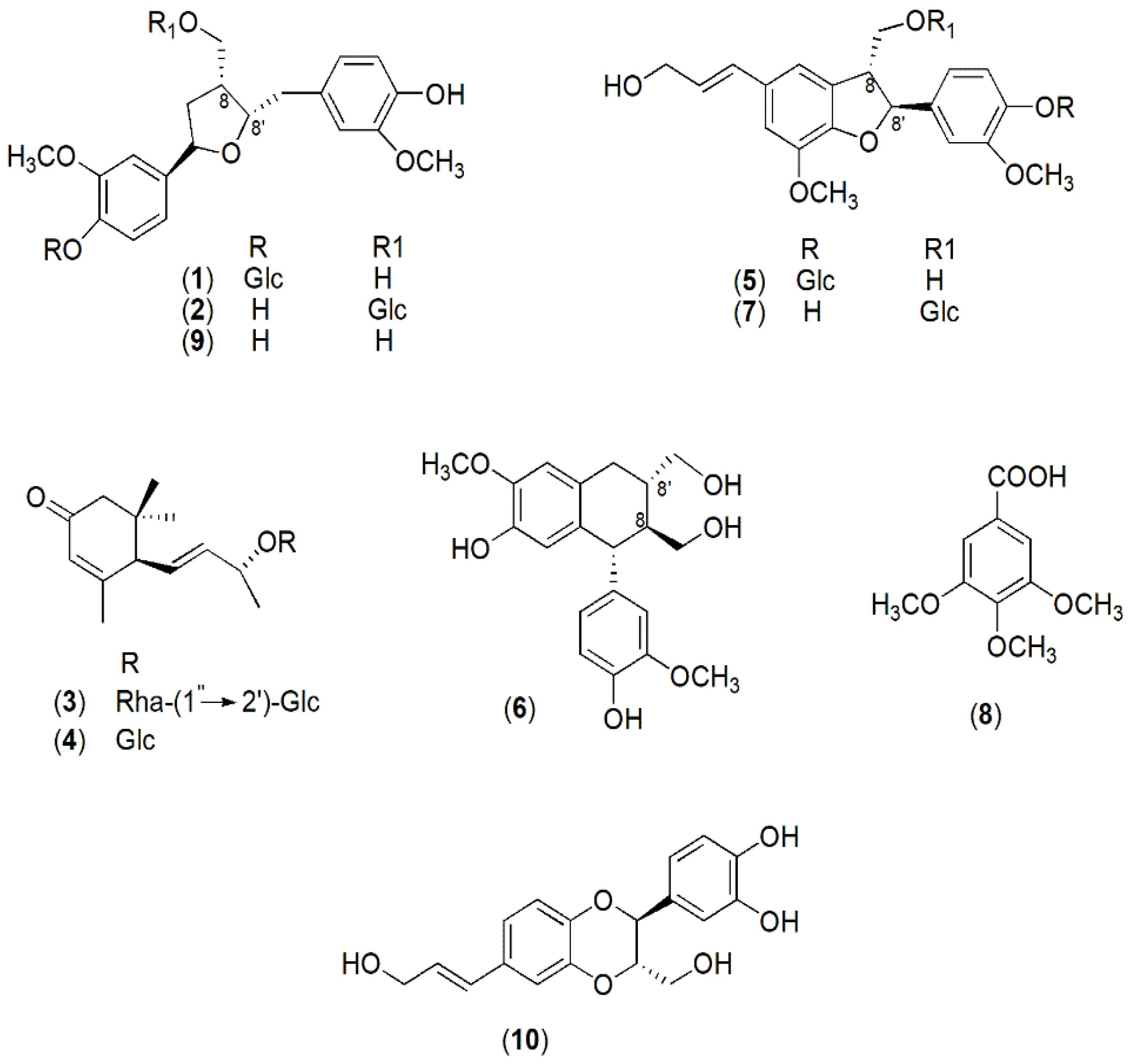
2.4. Chemical Profile and Biological Activity of Colocasia Constituents
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. General Experimental Procedures
4.3. Plant Material
4.4. Extraction Procedures
4.5. Isolation Procedure of Compounds 1–10
4.6. Spectroscopic Data of Compound 1–10
4.7. Quantitative HPLC Analysis of CCM
4.8. Cells
4.9. Determination of Cell Viability via MTT Assay
4.10. Cell Cycle Analysis
4.11. Apoptosis Analysis
4.12. Western Blot
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Otekunrin, O.A.; Sawicka, B.; Adeyonu, A.G.; Otekunrin, O.A. Cocoyam [Colocasia esculenta (L.) Schott ]: Exploring the Production, Health and Trade Potentials in Sub-Saharan Africa. Sustainability 2021, 13, 4483. [Google Scholar] [CrossRef]
- Grimaldi, I.M.; Muthukumaran, S.; Tozzi, G.; Nastasi, A.; Boivin, N.; Matthews, P.J.; Andel, T. Van Literary evidence for taro in the ancient Mediterranean: A chronology of names and uses in a multilingual world. PLoS ONE 2018, 13, e0198333. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Jan, R.; Kaur, R.; Riar, C.S. Taro (Colocasia esculenta). Antioxid. Veg. Nuts-Prop. Health Benefits 2020, 341–353. [Google Scholar] [CrossRef]
- Sudhakar, P.; Thenmozhi, V.; Srivignesh, S.; Dhanalakshmi, M. Colocasia esculenta (L.) Schott: Pharmacognostic and pharmacological review. J. Pharmacogn. Phytochem. 2020, 9, 1382–1386. [Google Scholar]
- Rashmi, D.R.; Raghu, N.; Gopenath, T.S.; Palanisamy, P.; Bakthavatchalam, P.; Karthikeyan, M.; Gnanasekaran, A.; Basalingappa, K.M. Taro (Colocasia esculenta): An overview. J. Med. Plants Stud. 2018, 6, 156–161. [Google Scholar]
- Ferreres, F.; Gonc, R.F.; Gil-izquierdo, A.; Silva, A.M.S.; Silva, B.; Andrade, P.B. Further Knowledge on the Phenolic Profile of Colocasia. J. Agric. Food Chem. 2012, 60, 7005–7015. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Dong, Z.; Liu, X.; Chen, H.; Lai, F.; Zhang, M. Structure characterization of two novel polysaccharides from Colocasia esculenta (taro) and a comparative study of their immunomodulatory activities. J. Funct. Foods 2018, 42, 47–57. [Google Scholar] [CrossRef]
- Keerthy, S.; Hanumanthachar, K.H. The pharmacological importance of Colocasia esculenta Linn: A review. J. Pharmacogn. Phytochem. 2019, 8, 1945–1948. [Google Scholar]
- Kumawat, N.S.; Chaudhari, S.P.; Wani, N.S.; Deshmukh, T.A.; Patil, V.R. Antidiabetic activity of ethanol extract of Colocasia esculenta leaves in alloxan induced diabetic rats. Int. J. Pharm. Tech. Res. 2010, 2, 1246–1249. [Google Scholar]
- Sakano, Y.; Mutsuga, M.; Tanaka, R.; Suganuma, H.; Inakuma, T.; Toyoda, M.; Goda, Y.; Shibuya, M.; Ebizuka, Y. Inhibition of Human Lanosterol Synthase by the Constituents of Colocasia esculenta (Taro). Biol. Pharm. Bull. 2005, 28, 299–304. [Google Scholar] [CrossRef]
- Pereira, P.R.; Carolina, A.; Teixeira, N.; Corr, F.; Vericimo, M.A.; Paschoalin, F. Tarin, a Potential Immunomodulator and COX-Inhibitor Lectin Found in Taro (Colocasia esculenta). Compr. Rev. Food Sci. Food Saf. 2018, 17, 878–891. [Google Scholar] [CrossRef] [PubMed]
- Kundu, N.; Campbell, P.; Hampton, B.; Lin, C.; Ma, X.; Ambulos, N.; Zhao, X.F.; Goloubeva, O.; Holt, D.; Fulton, A.M. Antimetastatic activity isolated from Colocasia esculenta (taro). Anti-Cancer Drugs 2012, 23, 200. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Lee, H.; Cho, S.Y.; Kim, Y.; Shin, K. Anti-metastatic effect of polysaccharide isolated from Colocasia esculenta is exerted through immunostimulation. Int. J. Mol. Med. 2013, 31, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro Pereira, P.; Bertozzi, É.; Mattos, D.A.; Carolina, A.; Teixeira, N.; Corr, F.; Vericimo, M.A.; Margaret, V.; Paschoalin, F. Anticancer and Immunomodulatory Benefits of Taro (Colocasia esculenta) Corms, an Underexploited Tuber Crop. Int. J. Mol. 2021, 22, 265. [Google Scholar] [CrossRef] [PubMed]
- Smyth, E.C.; Nilsson, M.; Grabsch, H.I.; Van Grieken, N.C.T.; Lordick, F. Gastric cancer. Lancet 2020, 396, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Bifulco, M.; Marasco, M.; Pisanti, S. School of Salerno a Lesson from the Past. AMEPRE 2008, 35, 602–603. [Google Scholar] [CrossRef]
- Brown, A.C.; Reitzenstein, J.E.; Liu, J.; Jadus, M.R. The Anti-Cancer Effects of Poi (Colocasia esculenta) on Colonic Adenocarcinoma Cells in vitro. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2005, 771, 767–771. [Google Scholar]
- Kai, H.; Akamatsu, E.; Torii, E.; Kodama, H.; Yukizaki, C. Inhibition of proliferation by agricultural plant extracts in seven human adult T-cell leukaemia (ATL)-related cell lines. J. Nat. Med. 2011, 65, 651–655. [Google Scholar] [CrossRef]
- Kundu, N.; Ma, X.; Hoag, S.; Wang, F.; Ibrahim, A.; Godoy-ruiz, R.; Weber, D.J.; Fulton, A.M. An Extract of Taro (Colocasia esculenta) Mediates Potent Inhibitory Actions on Metastatic and Cancer Stem Cells by Tumor Cell-Autonomous and Immune-Dependent Mechanisms. Breast Cancer Basic Clin. Res. 2021, 15, 11782234211034937. [Google Scholar] [CrossRef]
- Corrêa, A.; Vericimo, M.; Dashevskiy, A.; Pereira, P.; Paschoalin, V. Liposomal Taro Lectin Nanocapsules Control Human Glioblastoma and Mammary Adenocarcinoma Cell Proliferation. Molecules 2019, 29, 471. [Google Scholar] [CrossRef]
- Chan, Y.S.; Wong, J.H.; Ng, T.B. A Cytokine-Inducing Hemagglutinin from Small Taros. Protein Pept. Lett. 2010, 17, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Suski, J.M.; Braun, M.; Strmiska, V.; Sicinski, P.; Contact, L. Targeting Cell-cycle Machinery in Cancer. Cancer Cell 2021, 39, 759–778. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Prives, C. Increased and altered DNA binding of human p53 by S and G2/M but not G1 cyclin-dependent kinases. Nature 1995, 496, 279–281. [Google Scholar]
- De Luca, A.; Maiello, M.R.; D’Alessio, A.D.; Pergameno, M.; Normanno, N. The RAS/RAF/MEK/ERK and the PI3K/AKT signalling pathways: Role in cancer pathogenesis and implications for therapeutic approaches. Exp. Opin. Ther. Targets 2012, 16, S17–S27. [Google Scholar] [CrossRef] [PubMed]
- Connolly, J.L.; Rodgers, S.E.; Clarke, P.; Ballard, D.W.; Kerr, L.D.; Tyler, K.L.; Dermody, T.S.; Irol, J.V. Reovirus-Induced Apoptosis Requires Activation of Transcription Factor NF-kappaB. Exp. Opin. Ther. Targets 2000, 74, 2981–2989. [Google Scholar]
- Kimura, M.; Haisa, M.; Uetsuka, H.; Takaoka, M.; Ohkawa, T.; Kawashima, R.; Yamatsuji, T.; Gunduz, M.; Kaneda, Y.; Tanaka, N. TNF combined with IFN-a accelerates NF-j B-mediated apoptosis through enhancement of Fas expression in colon cancer cells. Cell Death Differ. 2003, 10, 718–728. [Google Scholar] [CrossRef]
- Kim, S.B.; Kim, J.S.; Lee, J.H.; Yoon, W.J.; Lee, D.S.; Ko, M.S.; Kwon, B.S.; Choi, D.H.; Cho, H.R.; Lee, B.J.; et al. NF-κB activation is required for cisplatin-induced apoptosis in head and neck squamous carcinoma cells. FEBS Lett. 2006, 580, 311–318. [Google Scholar] [CrossRef]
- Jennewein, C.; Karl, S.; Baumann, B.; Micheau, O.; Debatin, K.; Fulda, S. Identification of a novel pro-apoptotic role of NF- j B in the regulation of TRAIL-and CD95-mediated apoptosis of glioblastoma cells. Oncogene 2011, 31, 1468–1474. [Google Scholar] [CrossRef]
- Eom, J.W.; Lim, J.W.; Kim, H. Lutein Induces Reactive Oxygen Species-Mediated Apoptosis in Gastric Cancer AGS Cells via NADPH Oxidase Activation. Molecules 2023, 28, 1178. [Google Scholar] [CrossRef]
- Kumar, V.; Sharma, H.K. Process optimization for extraction of bioactive compounds from taro (Colocasia esculenta), using RSM and ANFIS modeling. Food Meas. 2017, 11, 704–718. [Google Scholar] [CrossRef]
- Agbor-egbe, T.; Rickard, J.E. Identification of Phenolic Compounds in Edible Aroids. J. Sci. Food Agric. 1990, 51, 215–221. [Google Scholar] [CrossRef]
- Lee, S.; Wee, W.; Yong, J.; Syamsumir, D. Antimicrobial, antioxidant, anticancer property and chemical composition of different parts (corm, stem and leave) of Colocasia esculenta extract. Ann. Univ. Mariae Curie-Sklodowska Pharm. 2011, 24, 9–16. [Google Scholar]
- Eleazu, C.O. Characterization of the natural products in cocoyam (Colocasia esculenta) using GC—MS. Pharm. Biol. 2016, 54, 2880–2885. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Moon, E.; Kim, S.Y.; Lee, K.R. Lignans from the Tuber-barks of Colocasia antiquorum var. esculenta and Their Antimelanogenic Activity. J. Agric. Food Chem. 2010, 58, 4779–4785. [Google Scholar] [CrossRef] [PubMed]
- Osmakov, D.I.; Kalinovskii, A.P.; Belozerova, O.A.; Andreev, Y.A.; Kozlov, S.A. Lignans as Pharmacological Agents in Disorders Related to Oxidative Stress and Inflammation: Chemical Synthesis Approaches and Biological Activities. Int. J. Mol. Sci. 2022, 23, 6031. [Google Scholar] [CrossRef] [PubMed]
- Zálešák, F.; Denis, D.J.; Pospíšil, J. Lignans and Neolignans: Plant secondary metabolites as a reservoir of biologically active substances. Pharmacol. Res. 2019, 146, 104284. [Google Scholar] [CrossRef]
- Jang, W.Y.; Kim, M.; Cho, J.Y. Antioxidant, Anti-Inflammatory, Anti-Menopausal, and Anti-Cancer Effects of Lignans and Their Metabolites. Int. J. Mol. Sci. 2022, 23, 15482. [Google Scholar] [CrossRef]
- Lee, S.Y.; Choi, S.U.; Lee, K.R. Three New Megastigmane Glycosides from Hylomecon vernalis. Bull. Korean Chem. Soc. 2011, 32, 3813–3816. [Google Scholar] [CrossRef]
- Quang, T.H.; Thanh, T.; Van Minh, C.; Van Kiem, P.; Yen, P.H.; Tai, H.; Nhiem, X.; Thao, P. Plantagiolides I and J, Two New Withanolide Glucosides from Tacca plantaginea with Nuclear Factor-kappaB Inhibitory and Peroxisome Proliferator-Activated Receptor Transactivational Activities. Chem. Pharm. Bull. 2012, 60, 1494–1501. [Google Scholar] [CrossRef]
- Zhang, Y.; Chao, L.; Ruan, J.; Zheng, C.; Yu, H.; Qu, L.; Han, L.; Wang, T. Fitoterapia Bioactive constituents from the rhizomes of Dioscorea septemloba Thunb. Fitoterapia 2016, 115, 165–172. [Google Scholar] [CrossRef]
- Scognamiglio, M.; Buommino, E.; Coretti, L.; Graziani, V.; Russo, R.; Caputo, P.; Donnarumma, G.; Abrosca, B.D.; Fiorentino, A. Phytochemistry Letters Phytochemical investigation and antimicrobial assessment of Bellis sylvestris leaves. Phytochem. Lett. 2016, 17, 6–13. [Google Scholar] [CrossRef]
- Pan, S.; Pirker, T.; Kunert, O.; Kretschmer, N.; Hummelbrunner, S.; Latkolik, S.L.; Rappai, J.; Dirsch, V.M.; Bochkov, V.; Bauer, R. C13 Megastigmane Derivatives From Epipremnum pinnatum: β-Damascenone Inhibits the Expression of Pro-Inflammatory Cytokines and Leukocyte Adhesion Molecules as Well as NF-κB Signaling. Front. Pharmacol. 2019, 10, 1351. [Google Scholar] [CrossRef] [PubMed]
- Esquivel-Campos, A.; Pérez-Gutiérrez, S.; Sánchez-Pérez, L.; Campos-Xolalpa, N.; Pérez-Ramos, J. Cytotoxicity and Antitumor Action of Lignans and Neolignans. In Secondary Metabolites-Trends and Reviews; IntechOpen: Rijeka, Croatia, 2022. [Google Scholar]
- Nhung, L.T.H.; Anh, N.T.H.; Tai, B.H.; Van Kiem, P. Isolation of lignans and neolignans from Pouzolzia sanguinea with their cytotoxic activity. Vietnam. J. Chem. 2021, 59, 146–152. [Google Scholar] [CrossRef]
- Kuang, H.; Xia, Y.; Yang, B.; Wang, Q.; Lü, S. Lignan Constituents from Chloranthus japonicus Sieb. Arch. Pharmacal Res. 2009, 32, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Van Ban, H.; Van, T.T.T.; Van Chien, V.; Hue, N.T.; Van Cuong, P.; Vuong, N.Q. Lignans from Leaves of Amesiodendron chinense and their cytotoxic activity. J. Sci. Technol. 2020, 58, 442–449. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant Phenolics: Extraction, Analysis and Their Antioxidant and Anticancer Properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Anuntiyo, J.; Malemud, C.J.; Haqqi, T.M. Biological Basis for the Use of Botanicals in Osteoarthritis and Rheumatoid Arthritis: A Review. Evid.-Based Complement. Altern. Med. 2005, 2, 301–308. [Google Scholar] [CrossRef]
- Ayouaz, S.; Oliveira-Alves, S.C.; Lefsih, K.; Serra, A.T.; Da Silva, A.B.; Samah, M.; Karczewski, J.; Madani, K.; Bronze, M.R. Phenolic compounds from Nerium oleander leaves: Antiproliferative and cytotoxic activities. Food Funct. 2020, 11, 6319–6331. [Google Scholar] [CrossRef]
- Aïssaoui, H.; Mencherini, T.; Esposito, T.; De Tommasi, N.; Gazzerro, P.; Benayache, S.; Benayache, F.; Mekkiou, R. Heliotropium bacciferum Forssk. (Boraginaceae) extracts: Chemical constituents, antioxidant activity and cytotoxic effect in human cancer cell lines. Nat. Prod. Res. 2018, 6419, 1813–1818. [Google Scholar] [CrossRef]
- Boudermine, S.; Malafronte, N.; Mencherini, T.; Esposito, T.; Aquino, R.P.; Beghidja, N.; Benayache, S.; D’Ambola, M.; Vassallo, A. Phenolic Compounds from Limonium pruinosum Sihem. Nat. Prod. Commun. 2015, 10, 319–321. [Google Scholar] [CrossRef]
- Minh, T.T.; Thi, N.; Thuong, M. Lignans from the stems of Clerodendrum inerme Gaertn. Vietnam. J. Chem. 2021, 59, 187–191. [Google Scholar] [CrossRef]
- Wang, L.Q.; Zhao, Y.X. Lignans from Gnetum montanum Markgr. f. megalocarpua. Chem. Nat. Compd. 2009, 45, 360–361. [Google Scholar]
- Yu, L.; Hu, W.; Ding, G.; Li, R.; Wei, J.; Zou, Z.; Wang, M. Gusanlungionosides A À D, Potential Tyrosinase Inhibitors from Arcangelisia gusanlung. J. Nat. Prod. 2011, 74, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Matsunami, K.; Otsuka, H.; Takeda, Y. Structural Revisions of Blumenol C Glucoside and Byzantionoside B. Chem. Pharm. Bull. 2010, 58, 438–441. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Park, M.; Park, Y.H. A lignan from Rubia akane. Arch. Pharmacal Res. 1990, 13, 289–291. [Google Scholar] [CrossRef]
- Akdemir, Z.Ş.; Tatli, İ.İ.; Bedir, E.; Khan, I.A. Neolignan and Phenylethanoid Glycosides from Verbascum salviifolium Boiss. Turk. J. Chem. 2004, 28, 621–628. [Google Scholar]
- Agbo, M.O.; Ezealisiji, K.M.; Elijah, J.; Ukekwe, F.I.; Obonga, W.O. Gallic Acid Derivatives (GADs) from Loranthus micranthus Linn. Parasitic on Gallic Acid Derivatives (GADs) from Loranthus micranthus Linn. Parasitic on Hevea brasiliensis with Antioxidative Capacity. Dhaka Univ. J. Pharm. Sci. 2016, 14, 139–145. [Google Scholar] [CrossRef]
- Fukuyama, Y.; Hasegawa, T.; Toda, M.; Kodama, M.; Okazaki, H. Structures of americanol A and isoamericanol A having a neurotrophic property from the seeds of Phytolacca americana. Chem. Pharm. Bull. 1992, 40, 252–254. [Google Scholar] [CrossRef]
- Picerno, P.; Sansone, F.; Mencherini, T.; Prota, L.; Aquino, R.P.; Rastrelli, L.; Lauro, M.R. Citrus bergamia juice: Phytochemical and technological studies. Nat. Prod. Commun. 2011, 6, 951–955. [Google Scholar] [CrossRef]
- Massaro, M.; Martinelli, R.; Gatta, V.; Scoditti, E. Transcriptome-based identification of new anti-anti-inflammatory and vasodilating properties of the n-3 fatty acid docosahexaenoic acid in vascular endothelial cell under proinflammatory conditions. PLoS ONE 2015, 10, e0129652. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esposito, T.; Pisanti, S.; Mauro, L.; Mencherini, T.; Martinelli, R.; Aquino, R.P. Activity of Colocasia esculenta (Taro) Corms against Gastric Adenocarcinoma Cells: Chemical Study and Molecular Characterization. Int. J. Mol. Sci. 2024, 25, 252. https://doi.org/10.3390/ijms25010252
Esposito T, Pisanti S, Mauro L, Mencherini T, Martinelli R, Aquino RP. Activity of Colocasia esculenta (Taro) Corms against Gastric Adenocarcinoma Cells: Chemical Study and Molecular Characterization. International Journal of Molecular Sciences. 2024; 25(1):252. https://doi.org/10.3390/ijms25010252
Chicago/Turabian StyleEsposito, Tiziana, Simona Pisanti, Luciano Mauro, Teresa Mencherini, Rosanna Martinelli, and Rita Patrizia Aquino. 2024. "Activity of Colocasia esculenta (Taro) Corms against Gastric Adenocarcinoma Cells: Chemical Study and Molecular Characterization" International Journal of Molecular Sciences 25, no. 1: 252. https://doi.org/10.3390/ijms25010252